野豌豆属43份牧草种质的染色体形态观察与分析
2015-06-24王彦荣刘志鹏
刘 鹏,王彦荣,刘志鹏
(兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃 兰州 730020)
野豌豆属43份牧草种质的染色体形态观察与分析
刘 鹏,王彦荣,刘志鹏
(兰州大学草地农业科技学院,草地农业生态系统国家重点实验室,甘肃 兰州 730020)
野豌豆属(Vicia)牧草为自花授粉二倍体植物,具有很好的防风固沙及保水作用,适宜在高寒和高海拔地区种植,是我国高原牧区重要的蛋白饲料来源。本研究从中国、美国、德国、埃塞俄比亚等22个国家,收集了43份野豌豆属牧草种质,对其根尖细胞的有丝分裂中期染色体进行核型分析。结果表明,染色体数目有5种:10、12、14、16、24。核型分类有6种:2A、1B、2B、3B、2C、3C。本研究统计了野豌豆属43份种质的核型数据,为揭示野豌豆属细胞学特性和演化趋势奠定了基础。
野豌豆属;染色体;核型分析
野豌豆属(Vicia)牧草属于自花传粉的二倍体物种,是世界上最重要的一年或多年生豆科植物[1]。该属植物约有190个物种,Ball[2]将该属划分成4个亚属:Vicia,Cracca,Ervum和Faba。随后研究人员进一步分析了Vicia和Vicilla两个亚属的生物学特性,虽然Vicia亚属的物种少于Vicilla亚属,但是Vicia亚属包括很多具有重要农业价值的牧草[3-5]。Maxted[6]在原表型分类的基础上又将Vicia亚属细分为9大系列,38个物种,14个亚种。
由于其生育周期短,营养价值高,在农业中用途广泛,可用于干草、粮食、青贮饲料、绿肥等[7-8]。其中箭筈豌豆作为野豌豆属常见的牧草饲料,南志标等[9]通过在青藏高原地区引种野豌豆属(Viciaspp.)和山黛豆属(Lathyrusspp.),成功培育了适宜在高寒和高海拔地区种植的牧草种质兰箭3号以及品系2556和2560,其具有早熟,生育周期短,种子产量高等特点。其中兰箭3号种子产量很高,且能有效改善土壤肥力。品系2556和2560的地上生物量较高,营养丰富。生产和区域试验表明,该品种可在黄土高原地区,作为秋播作物收割后的复种作物种植,是高原牧区重要的蛋白饲料来源[10-12]。
野豌豆属植物主要分布在北半球温带地区,拉丁美洲有零星分布[13]。根据中国数字植物标本馆(http://www.cvh.org.cn/cms/)和宋敏等[14]的统计结果表明,在我国本属植物广布于全国各省,华北、西北、西南较多,其中栽培种5种,野生种36种,华南和台湾地区分布很少,仅有野生种1种。目前,对野豌豆属植物细胞染色体多样性的研究相对较少。对其染色体生物学特性研究,可为植物物种的科学分类和种质资源鉴定提供重要的形态依据。
关于野豌豆属植物染色体形态的研究,国内外学者已在野豌豆属核型上开展过相关工作,对V.amoena,V.cracca,V.pannonica及V.sativa等野豌豆属物种核型进行了相关研究,Castiglione等[15]和Caputo等[16]观察V.sativa,V.lathyroides,V.sativasubsp.sativa及V.grandiflora的染色体数目分别是12,12,12和14,通过聚类分析表明V.lathyroides和V.sativasubsp.sativa及V.grandiflora的亲缘关系较远。V.sativa与V.grandiflora的亲缘关系也较远。V.galeata,V.hyrcanica和V.anatolica的染色体数目分别是12,12和10,通过聚类分析表明V.galeata和V.hyrcanica的亲缘关系较近,V.galeata,V.hyrcanica两者与V.grandiflora的亲缘关系较远。
国内学者对于该属核型也进行了相关研究,如苏坤梅和杨德奎[17]所研究的V.amoena材料均来源于千佛山。冉雪琴等[18]对贵州野豌豆属4种植物核型进行了研究。管超等[19]对野豌豆属4个品种(系)的染色体核型进行了观察,主要是春箭筈豌豆和狭叶野豌豆(333/A)。尽管国内学者对该属展开了相关工作,但材料来源都局限于中国,存在一定的地域限制性。
染色体是细胞水平遗传物质的表征,与生长形态相比,染色体不易受外部环境影响,可作为物种起源、分类、演化和鉴别的重要依据[20-21]。本研究收集了中国、德国、埃塞俄比亚和加拿大的野豌豆属染色体材料,使野豌豆属细胞学染色体资料具有广域性,更加完善。本研究对来自22个国家的43份野豌豆属植物材料的染色体形态进行了观察,获得了新材料的染色体形态学信息,为该属植物染色体图像数据库的建立提供部分细胞学证据,以期为其种质资源多样性研究和新品种选育提供理论依据[19-21]。
1 材料和方法
1.1 试验材料及来源
野豌豆属植物种子由美国国家种质资源库(NPGS)和兰州大学草地农业科技学院提供。材料具体名称及来源地如表1所示。
1.2 试验方法
室温下,将成熟种子浸泡12 h,于20 ℃恒温培养箱中培养萌发,待根长至2~3 cm时于上午9:00-10:00时切取1 cm长度的根尖,用饱和对二氯苯和8-羟基喹啉的1:1混合液在15 ℃下处理3 h,蒸馏水冲洗5次,卡诺氏固定液室温固定24 h,其中(甲醇∶冰醋酸=3∶1),蒸馏水反复冲洗。将根尖置于1 mol·L-1的HCl中60 ℃解离5~10 min,将根尖放在载玻片上,用刀片将白色不透明部分切下,滴加卡宝品红染液及1滴45%醋酸,染色30 min,常规压片法制片[22-23],镜检。
1.3 核型分析
用Nikon Eclipse 80i显微镜进行镜检,用显微图像捕捉系统(NIS-Element SF3.0)进行拍照。每份材料随机观察30个细胞,选取5个分散良好的中期分裂相细胞进行拍照分析。根据李懋学和陈瑞阳[24]制定的标准进行核型分析,按照Stebbins[25]的分类标准模式划分类型,依据Arano[26]的方法对核型不对称系数(As.K%)进行测算,染色体统计按常规统计学方法[27]。
2 结果
2.1 染色体数目
对43份野豌豆属植物材料进行了染色体计数及核型分析,染色体形态照片及模式图见图1、2、3,核型公式及核型参数见表2、3。

表1 43份野豌豆属种质名称及其来源

图1 43份野豌豆属种质的染色体形态、核型图和核型模式图Fig.1 Morphology of somatic chromosomes and karyotypes, idiograms of 43 Vicia accessions

续图1(1) 43份野豌豆属种质的染色体形态、核型图和核型模式图 ContinuedFig.1(1) Morphology of somatic chromosomes and karyotypes, idiograms of 43 Vicia accessions
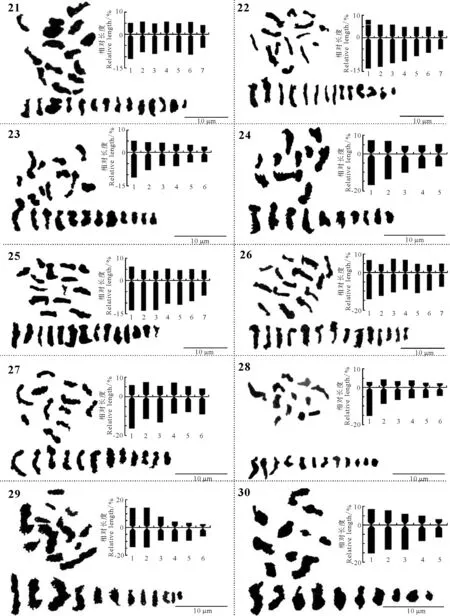
续图1(2) 43份野豌豆属种质的染色体形态、核型图和核型模式图 ContinuedFig.1(2) Morphology of somatic chromosomes and karyotypes, idiograms of 43 Vicia accessions

续图1(3) 43份野豌豆属种质的染色体形态、核型图和核型模式图 ContinuedFig.1(3) Morphology of somatic chromosomes and karyotypes, idiograms of 43 Vicia accessions
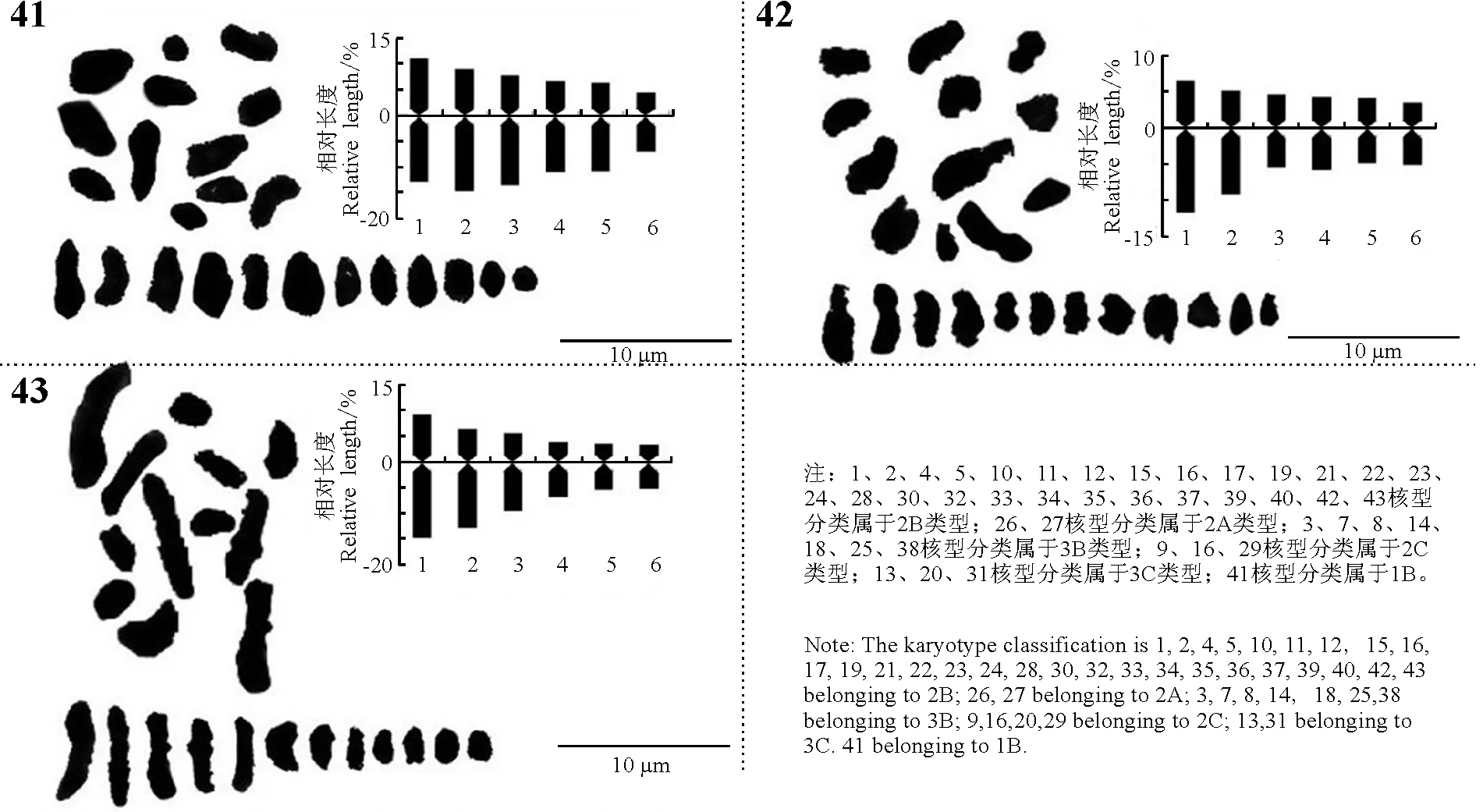
续图1(4) 43份野豌豆属种质的染色体形态、核型图和核型模式图 ContinuedFig.1(4) Morphology of somatic chromosomes and karyotypes, idiograms of 43 Vicia accessions
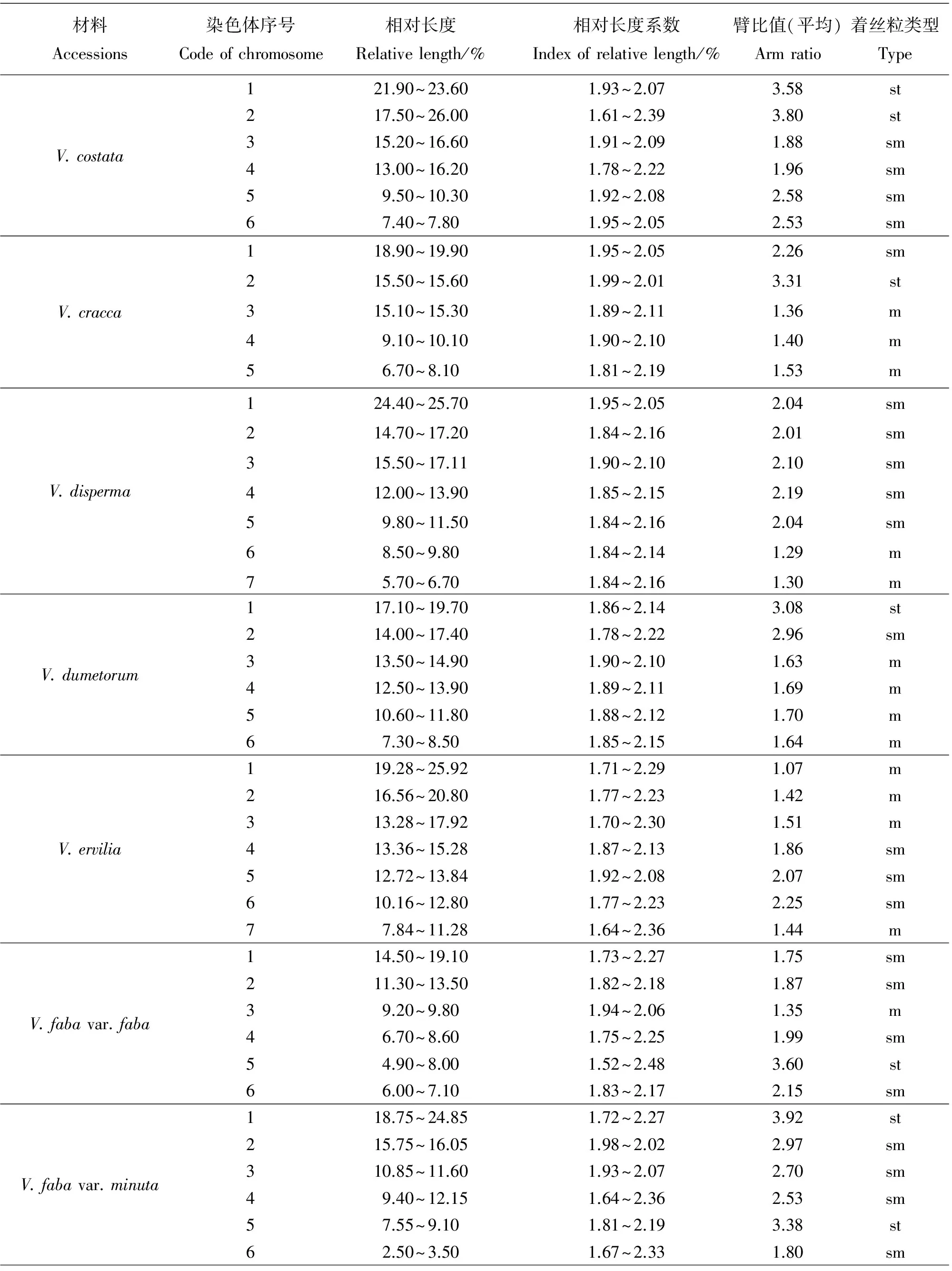
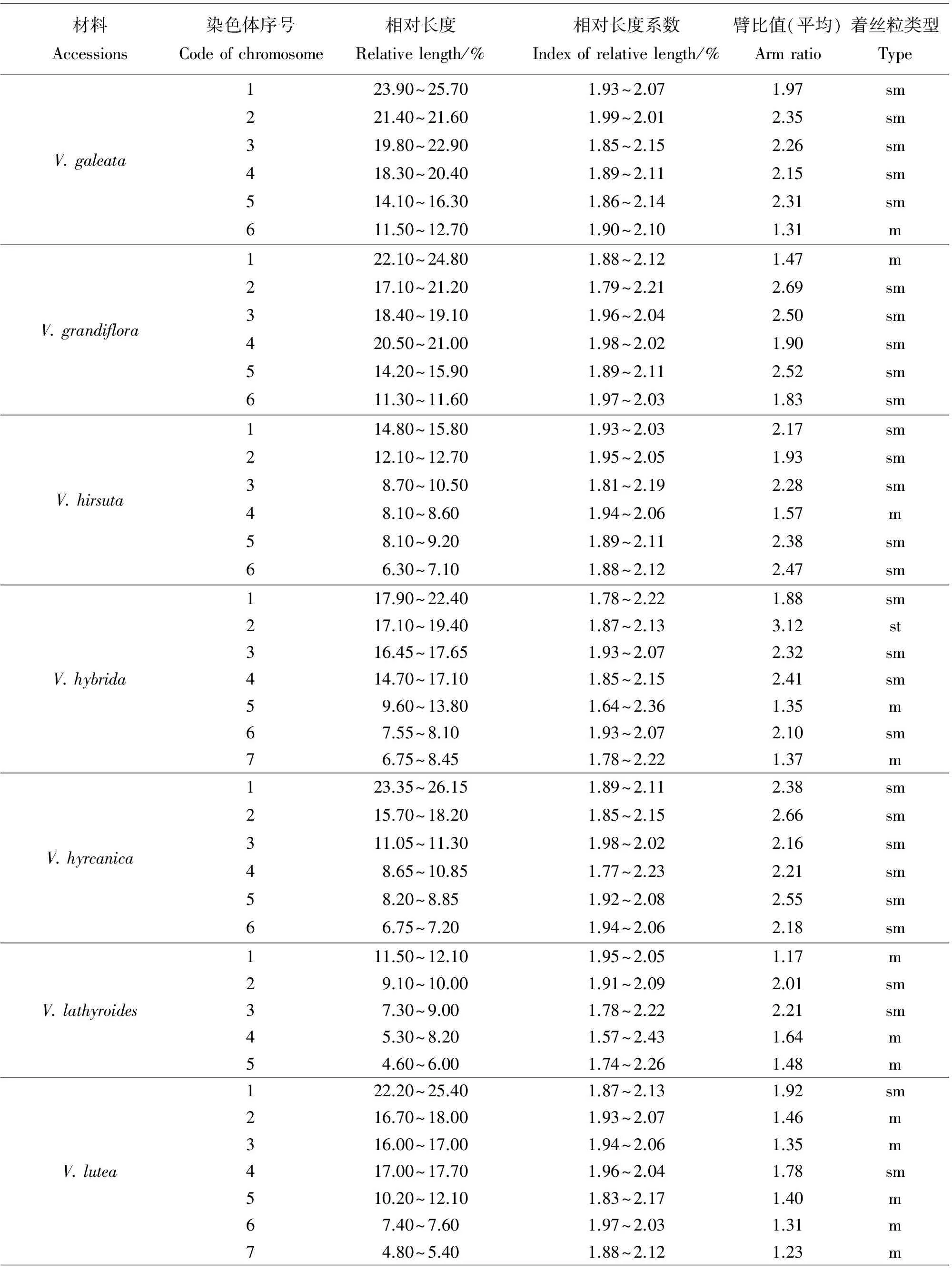
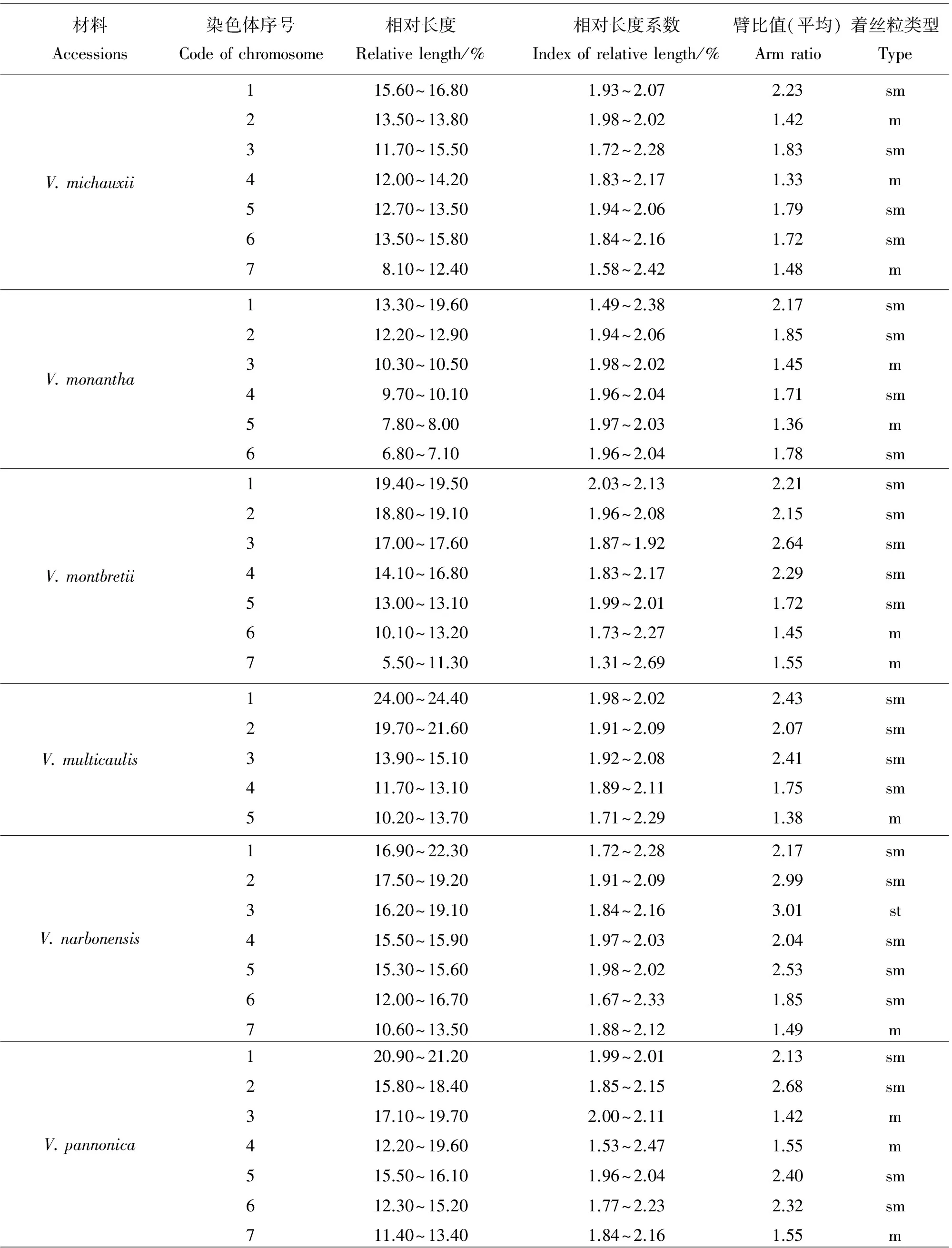
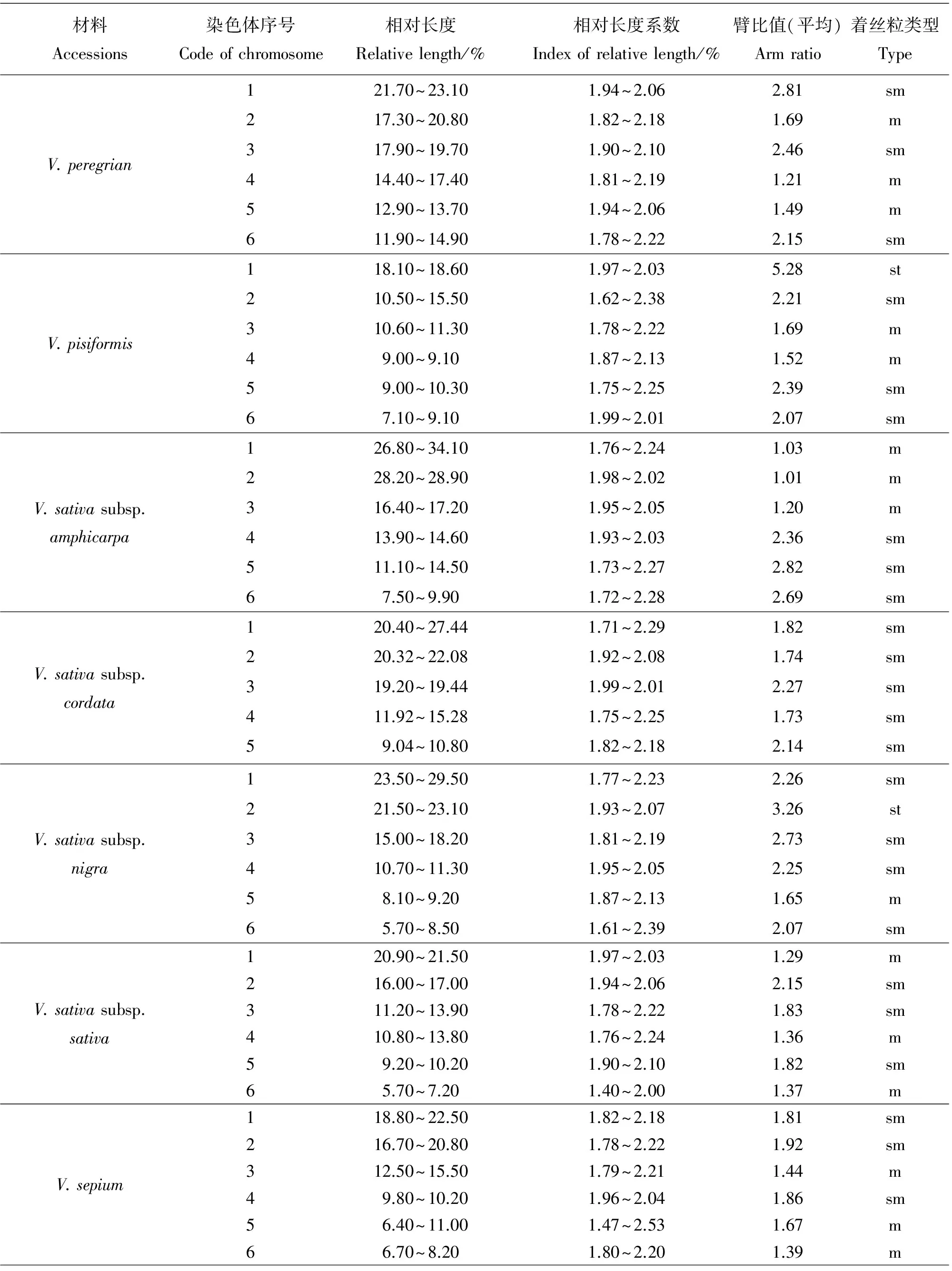
染色体数目有5种类型,分别是10,12,14,16和24条染色体。其中染色体数目2n=2x=10的种质有8份,V.anatolica、V.articulata、V.cracca和V.lathyroides等,均为二倍体,核型不对称系数变化范围59.72%~67.72%,臂比>2所占全部染色体比例范围是0.10~0.60;最长染色体与最短染色体的比值介于2.26~3.04;除V.cracca是3B类型外其余7份种质均为2B类型。染色体数目2n=2x=12的种质有23份,V.americana、V.benghalensis、V.fabavar.faba和V.sativa2505等,均为二倍体,核型不对称系数变化范围58.06%~74.76%,臂比>2所占全部染色体比例范围是0.00~0.83;最长染色体与最短染色体的比值介于1.74~9.94。核型类型1B和2A各1份,分别是V.sativa2505和V.peregrian。2B类型有13份种质,是V.americana、V.dumetorum、V.fabavar.faba和V.grandiflora等。核型类型是3B的种质有4份,分别是V.costata、V.galeata、V.hyrcanica和V.unijuga。属于2C类型的种质有2份,分别是V.benghalensis和V.sativasubsp.amphicarpa。核型分类属于3C的种质有2份,分别是V.sativasubsp.nigra和V.fabavar.minuta。
染色体数目2n=2x=14的种质有10份,V.aintabensis、V.disperma、V.ervilia和V.narbonensis等。均为二倍体,核型不对称系数变化范围是60.50%~69.17%,臂比>2所占全部染色体比例范围是0.07~0.64;最长染色体与最短染色体的比值介于1.86~5.29;核型类型2A种质有1份,是V.pannonica。2B类型有6份种质,是V.aintabensis、V.ervilia、V.hybrida和V.grandiflora等。核型类型3B种质有1份,是V.narbonensis;属于2C类型的种质有2份,分别是V.disperma和V.lutea。
染色体数目2n=2x=16的种质有1份,是V.tenuifoliasubsp.tenuifolia,为二倍体,核型不对称系数67.22%,臂比>2所占全部染色体比例为0.19;最长染色体与最短染色体的比值是2.72;核型类型为2B。染色体数目2n=4x=24的种质有1份,是V.amoena,为四倍体,核型不对称系数70.30%,臂比>2所占全部染色体比例为0.83;最长染色体与最短染色体的比值是3.75,核型类型为3B。
2.2 核型分类
按照核型分类标准,43份种质分为6类,核型属于1B只有1份种质,是V.sativa2505,核型不对称系数范围是60.78%,臂比>2所占全部染色体比例为0.00,最长染色体与最短染色体的比值范围是2.86。核型2A有2份种质,分别是V.pannonica和V.peregrian。核型不对称系数范围是65.28%~65.34%,臂比>2所占全部染色体比例为0.43~0.50,最长染色体与最短染色体的比值范围是1.86~1.94。
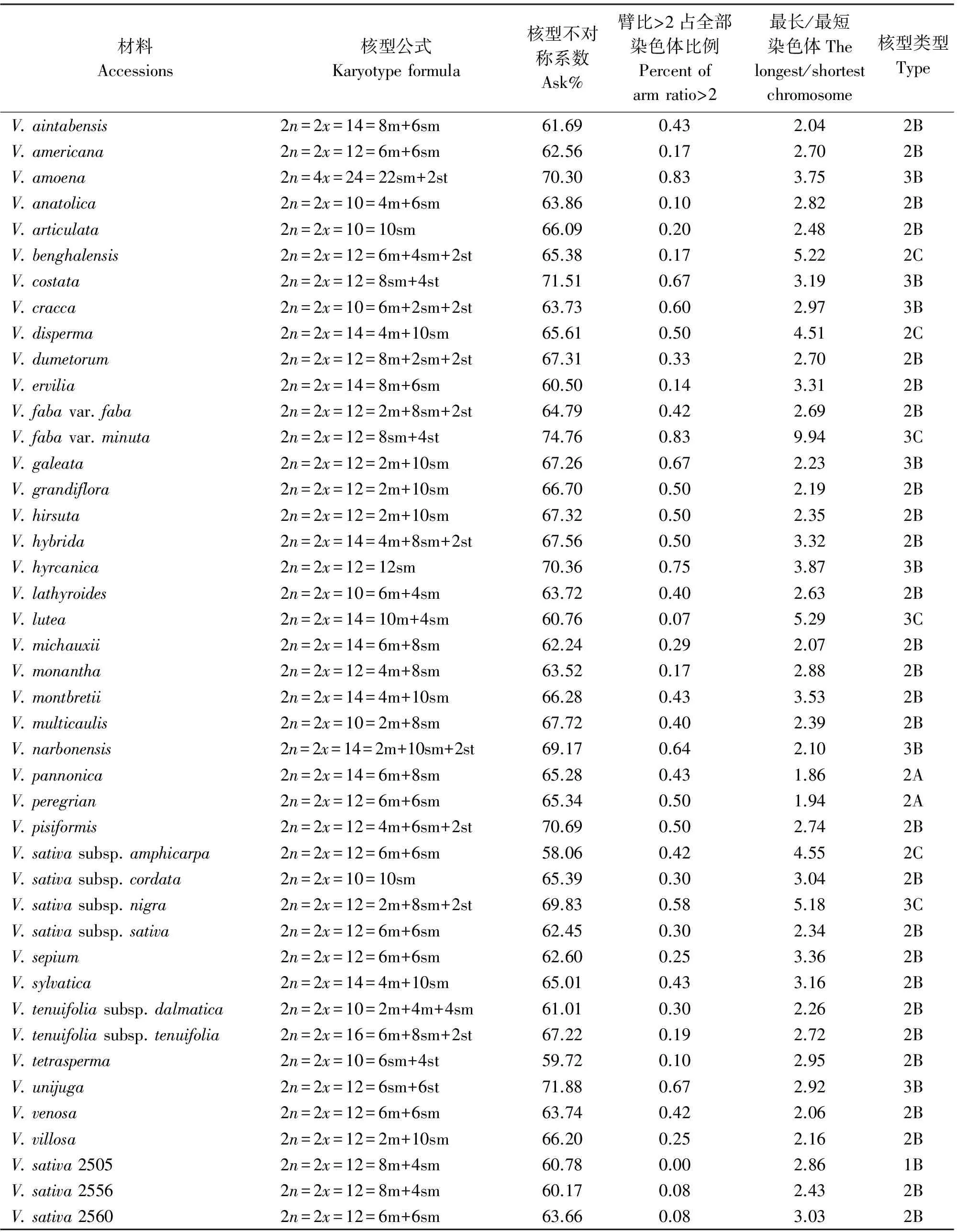
表2 野豌豆属43份种质的染色体核型参数
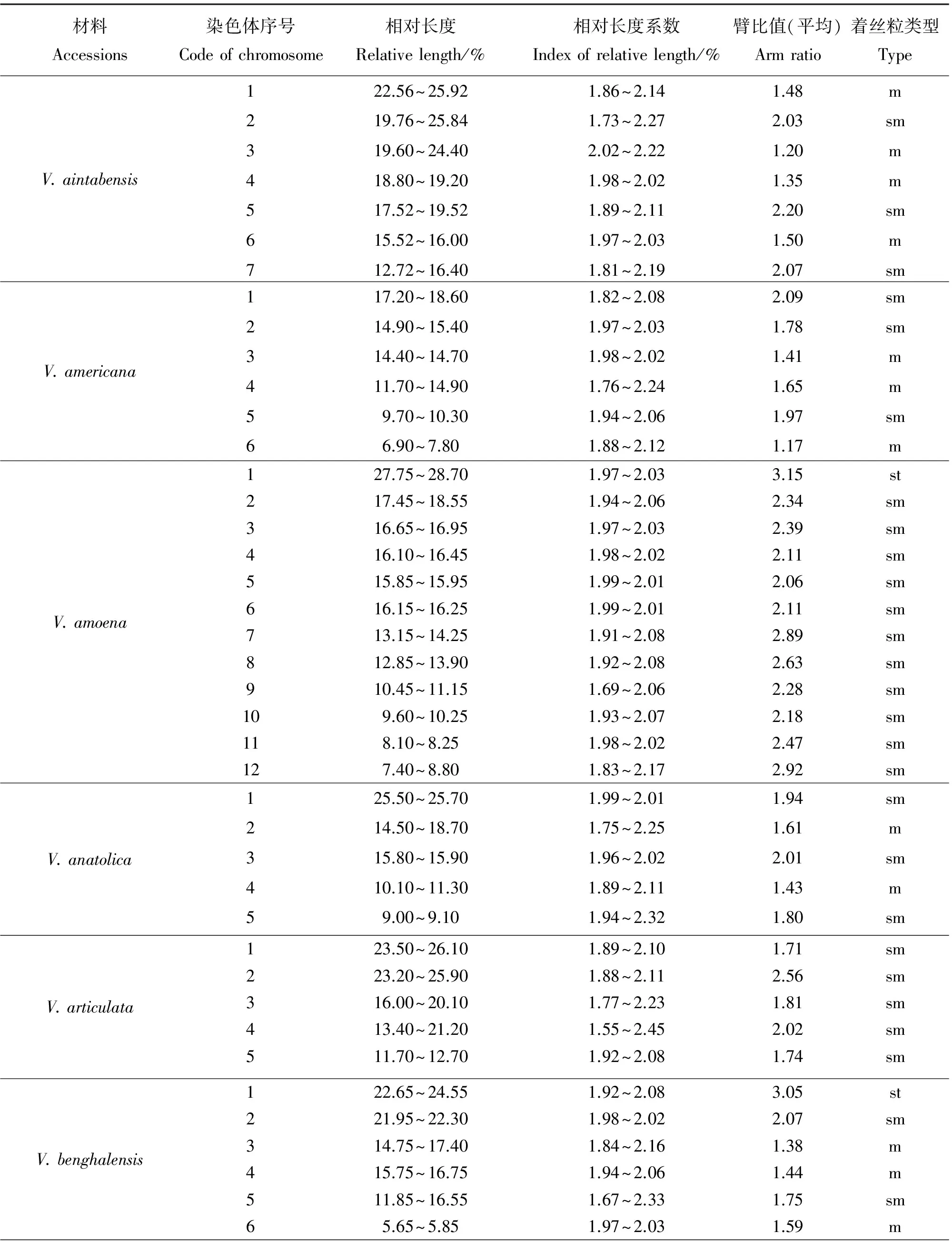
表3 野豌豆属不同物种的核型参数比较
续表3(1)
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.costata121.90~23.601.93~2.073.58st217.50~26.001.61~2.393.80st315.20~16.601.91~2.091.88sm413.00~16.201.78~2.221.96sm59.50~10.301.92~2.082.58sm67.40~7.801.95~2.052.53smV.cracca118.90~19.901.95~2.052.26sm215.50~15.601.99~2.013.31st315.10~15.301.89~2.111.36m49.10~10.101.90~2.101.40m56.70~8.101.81~2.191.53mV.disperma124.40~25.701.95~2.052.04sm214.70~17.201.84~2.162.01sm315.50~17.111.90~2.102.10sm412.00~13.901.85~2.152.19sm59.80~11.501.84~2.162.04sm68.50~9.801.84~2.141.29m75.70~6.701.84~2.161.30mV.dumetorum117.10~19.701.86~2.143.08st214.00~17.401.78~2.222.96sm313.50~14.901.90~2.101.63m412.50~13.901.89~2.111.69m510.60~11.801.88~2.121.70m67.30~8.501.85~2.151.64mV.ervilia119.28~25.921.71~2.291.07m216.56~20.801.77~2.231.42m313.28~17.921.70~2.301.51m413.36~15.281.87~2.131.86sm512.72~13.841.92~2.082.07sm610.16~12.801.77~2.232.25sm77.84~11.281.64~2.361.44mV.fabavar.faba114.50~19.101.73~2.271.75sm211.30~13.501.82~2.181.87sm39.20~9.801.94~2.061.35m46.70~8.601.75~2.251.99sm54.90~8.001.52~2.483.60st66.00~7.101.83~2.172.15smV.fabavar.minuta118.75~24.851.72~2.273.92st215.75~16.051.98~2.022.97sm310.85~11.601.93~2.072.70sm49.40~12.151.64~2.362.53sm57.55~9.101.81~2.193.38st62.50~3.501.67~2.331.80sm
续表3(2)
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.galeata123.90~25.701.93~2.071.97sm221.40~21.601.99~2.012.35sm319.80~22.901.85~2.152.26sm418.30~20.401.89~2.112.15sm514.10~16.301.86~2.142.31sm611.50~12.701.90~2.101.31mV.grandiflora122.10~24.801.88~2.121.47m217.10~21.201.79~2.212.69sm318.40~19.101.96~2.042.50sm420.50~21.001.98~2.021.90sm514.20~15.901.89~2.112.52sm611.30~11.601.97~2.031.83smV.hirsuta114.80~15.801.93~2.032.17sm212.10~12.701.95~2.051.93sm38.70~10.501.81~2.192.28sm48.10~8.601.94~2.061.57m58.10~9.201.89~2.112.38sm66.30~7.101.88~2.122.47smV.hybrida117.90~22.401.78~2.221.88sm217.10~19.401.87~2.133.12st316.45~17.651.93~2.072.32sm414.70~17.101.85~2.152.41sm59.60~13.801.64~2.361.35m67.55~8.101.93~2.072.10sm76.75~8.451.78~2.221.37mV.hyrcanica123.35~26.151.89~2.112.38sm215.70~18.201.85~2.152.66sm311.05~11.301.98~2.022.16sm48.65~10.851.77~2.232.21sm58.20~8.851.92~2.082.55sm66.75~7.201.94~2.062.18smV.lathyroides111.50~12.101.95~2.051.17m29.10~10.001.91~2.092.01sm37.30~9.001.78~2.222.21sm45.30~8.201.57~2.431.64m54.60~6.001.74~2.261.48mV.lutea122.20~25.401.87~2.131.92sm216.70~18.001.93~2.071.46m316.00~17.001.94~2.061.35m417.00~17.701.96~2.041.78sm510.20~12.101.83~2.171.40m67.40~7.601.97~2.031.31m74.80~5.401.88~2.121.23m
续表3(3)
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.michauxii115.60~16.801.93~2.072.23sm213.50~13.801.98~2.021.42m311.70~15.501.72~2.281.83sm412.00~14.201.83~2.171.33m512.70~13.501.94~2.061.79sm613.50~15.801.84~2.161.72sm78.10~12.401.58~2.421.48mV.monantha113.30~19.601.49~2.382.17sm212.20~12.901.94~2.061.85sm310.30~10.501.98~2.021.45m49.70~10.101.96~2.041.71sm57.80~8.001.97~2.031.36m66.80~7.101.96~2.041.78smV.montbretii119.40~19.502.03~2.132.21sm218.80~19.101.96~2.082.15sm317.00~17.601.87~1.922.64sm414.10~16.801.83~2.172.29sm513.00~13.101.99~2.011.72sm610.10~13.201.73~2.271.45m75.50~11.301.31~2.691.55mV.multicaulis124.00~24.401.98~2.022.43sm219.70~21.601.91~2.092.07sm313.90~15.101.92~2.082.41sm411.70~13.101.89~2.111.75sm510.20~13.701.71~2.291.38mV.narbonensis116.90~22.301.72~2.282.17sm217.50~19.201.91~2.092.99sm316.20~19.101.84~2.163.01st415.50~15.901.97~2.032.04sm515.30~15.601.98~2.022.53sm612.00~16.701.67~2.331.85sm710.60~13.501.88~2.121.49mV.pannonica120.90~21.201.99~2.012.13sm215.80~18.401.85~2.152.68sm317.10~19.702.00~2.111.42m412.20~19.601.53~2.471.55m515.50~16.101.96~2.042.40sm612.30~15.201.77~2.232.32sm711.40~13.401.84~2.161.55m
续表3(4)
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.peregrian121.70~23.101.94~2.062.81sm217.30~20.801.82~2.181.69m317.90~19.701.90~2.102.46sm414.40~17.401.81~2.191.21m512.90~13.701.94~2.061.49m611.90~14.901.78~2.222.15smV.pisiformis118.10~18.601.97~2.035.28st210.50~15.501.62~2.382.21sm310.60~11.301.78~2.221.69m49.00~9.101.87~2.131.52m59.00~10.301.75~2.252.39sm67.10~9.101.99~2.012.07smV.sativasubsp.amphicarpa126.80~34.101.76~2.241.03m228.20~28.901.98~2.021.01m316.40~17.201.95~2.051.20m413.90~14.601.93~2.032.36sm511.10~14.501.73~2.272.82sm67.50~9.901.72~2.282.69smV.sativasubsp.cordata120.40~27.441.71~2.291.82sm220.32~22.081.92~2.081.74sm319.20~19.441.99~2.012.27sm411.92~15.281.75~2.251.73sm59.04~10.801.82~2.182.14smV.sativasubsp.nigra123.50~29.501.77~2.232.26sm221.50~23.101.93~2.073.26st315.00~18.201.81~2.192.73sm410.70~11.301.95~2.052.25sm58.10~9.201.87~2.131.65m65.70~8.501.61~2.392.07smV.sativasubsp.sativa120.90~21.501.97~2.031.29m216.00~17.001.94~2.062.15sm311.20~13.901.78~2.221.83sm410.80~13.801.76~2.241.36m59.20~10.201.90~2.101.82sm65.70~7.201.40~2.001.37mV.sepium118.80~22.501.82~2.181.81sm216.70~20.801.78~2.221.92sm312.50~15.501.79~2.211.44m49.80~10.201.96~2.041.86sm56.40~11.001.47~2.531.67m66.70~8.201.80~2.201.39m
续表3(5)
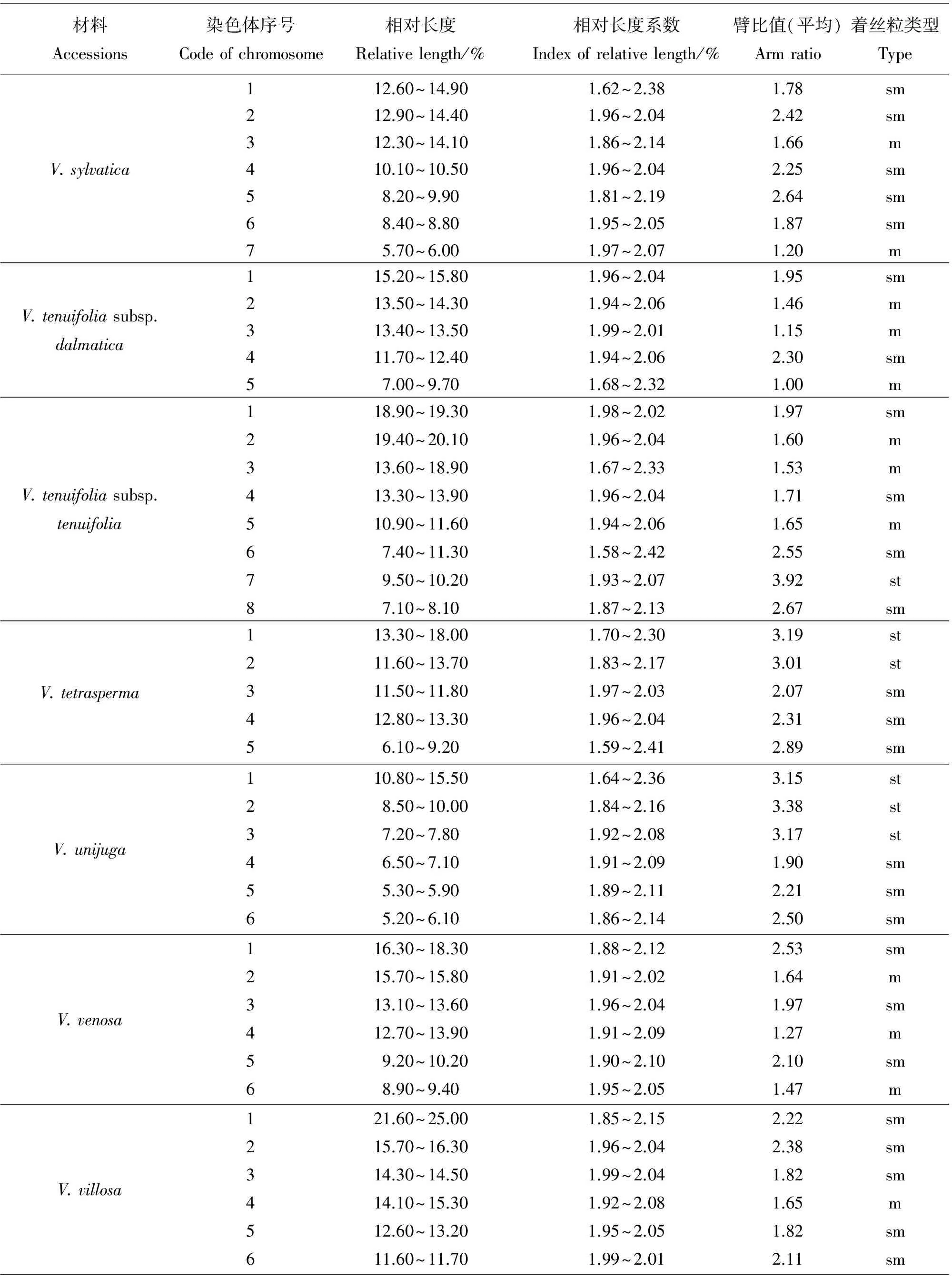
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.sylvatica112.60~14.901.62~2.381.78sm212.90~14.401.96~2.042.42sm312.30~14.101.86~2.141.66m410.10~10.501.96~2.042.25sm58.20~9.901.81~2.192.64sm68.40~8.801.95~2.051.87sm75.70~6.001.97~2.071.20mV.tenuifoliasubsp.dalmatica115.20~15.801.96~2.041.95sm213.50~14.301.94~2.061.46m313.40~13.501.99~2.011.15m411.70~12.401.94~2.062.30sm57.00~9.701.68~2.321.00mV.tenuifoliasubsp.tenuifolia118.90~19.301.98~2.021.97sm219.40~20.101.96~2.041.60m313.60~18.901.67~2.331.53m413.30~13.901.96~2.041.71sm510.90~11.601.94~2.061.65m67.40~11.301.58~2.422.55sm79.50~10.201.93~2.073.92st87.10~8.101.87~2.132.67smV.tetrasperma113.30~18.001.70~2.303.19st211.60~13.701.83~2.173.01st311.50~11.801.97~2.032.07sm412.80~13.301.96~2.042.31sm56.10~9.201.59~2.412.89smV.unijuga110.80~15.501.64~2.363.15st28.50~10.001.84~2.163.38st37.20~7.801.92~2.083.17st46.50~7.101.91~2.091.90sm55.30~5.901.89~2.112.21sm65.20~6.101.86~2.142.50smV.venosa116.30~18.301.88~2.122.53sm215.70~15.801.91~2.021.64m313.10~13.601.96~2.041.97sm412.70~13.901.91~2.091.27m59.20~10.201.90~2.102.10sm68.90~9.401.95~2.051.47mV.villosa121.60~25.001.85~2.152.22sm215.70~16.301.96~2.042.38sm314.30~14.501.99~2.041.82sm414.10~15.301.92~2.081.65m512.60~13.201.95~2.051.82sm611.60~11.701.99~2.012.11sm
续表3(6)
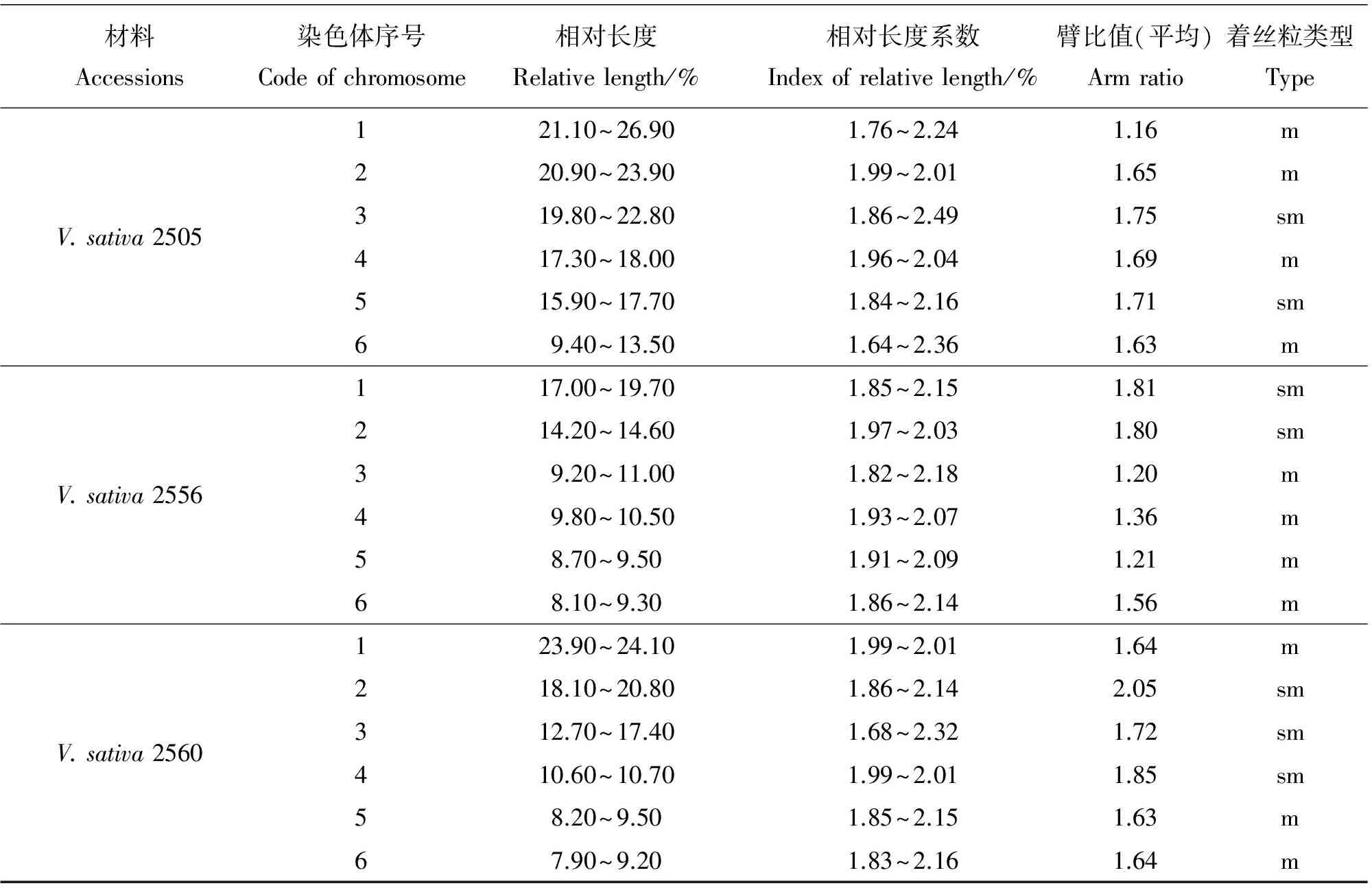
材料Accessions染色体序号Codeofchromosome相对长度Relativelength/%相对长度系数Indexofrelativelength/%臂比值(平均)Armratio着丝粒类型TypeV.sativa2505121.10~26.901.76~2.241.16m220.90~23.901.99~2.011.65m319.80~22.801.86~2.491.75sm417.30~18.001.96~2.041.69m515.90~17.701.84~2.161.71sm69.40~13.501.64~2.361.63mV.sativa2556117.00~19.701.85~2.151.81sm214.20~14.601.97~2.031.80sm39.20~11.001.82~2.181.20m49.80~10.501.93~2.071.36m58.70~9.501.91~2.091.21m68.10~9.301.86~2.141.56mV.sativa2560123.90~24.101.99~2.011.64m218.10~20.801.86~2.142.05sm312.70~17.401.68~2.321.72sm410.60~10.701.99~2.011.85sm58.20~9.501.85~2.151.63m67.90~9.201.83~2.161.64m
核型2B有27份种质,V.aintabensis、V.americana、V.ervilia和V.sativa2556等。核型不对称系数范围是59.72%~70.69%,臂比>2所占全部染色体比例范围为0.08~0.50,最长染色体与最短染色体的比值范围是2.04~3.53。核型3B有7份种质,V.amoena、V.costata、V.cracca和V.narbonensis等。核型不对称系数范围是63.73%~71.88%,臂比>2所占全部染色体比例范围为0.60~0.83,最长染色体与最短染色体的比值范围是2.10~3.87。核型2C有3份种质,分别是V.benghalensis、V.disperma和V.sativasubsp.amphicarpa。核型不对称系数范围是58.06%~65.61%,臂比>2所占全部染色体比例范围为0.17~0.50,最长染色体与最短染色体的比值范围是4.51~5.22。核型3C有3份种质,分别是V.fabavar.minuta、V.lutea和V.sativasubsp.nigra。核型不对称系数范围是60.76%~74.76%,臂比>2所占全部染色体比例范围为0.07~0.83,最长染色体与最短染色体的比值范围是5.18~9.94。
野豌豆属植物染色体一般由中部着丝点染色体m,近中部着丝点染色体sm和近端部着丝粒染色体st;3种染色体构成。V.articulata、V.sativasubsp.cordata和V.hyrcanica完全由近中部着丝点染色体sm构成,染色体数目分别是10、10和12条。在43份种质中只有V.montbretii和V.tenuifoliasubsp.dalmatica有随体,分别在第1和第4对染色体上。染色体相对长度变异最大为V.amoena,范围是7.40%~28.70%;相对长度变异最小为V.unijuga范围是5.20%~15.50%。剩余41份种质相对长度变异介于V.unijuga和V.amoena。
3 讨论与结论
染色体作为遗传信息的载体,结构和数目的变化是物种进化的主要内容,对染色体规律和变化趋势的研究,在实践和理论上具有重要意义[28-29]。本研究通过对来自不同国家的43份野豌豆属种质的核型统计分析,体细胞染色体倍性为二倍体(除V.amoena),基础染色体数目为5,6,7,8基础染色体数目多变。由此说明,该属植物为了适应不同生活环境,在长期进化过程中,染色体数目和结构呈现相关的演化趋势,造成核型发生改变。物种进化与细胞中染色体数目的改变紧密相连,染色体的多倍化是物种核型进化的重要机理,说明V.amoena处于相对进化的地位[30]。张红梅等[31]于2014年首次报道了V.tenuifoliasubsp.tenuifolia的核型数据,丰富了野豌豆属的细胞学资料。
野豌豆属不同种质在染色体数目、形态结构等方面具有相似性,这说明他们具有一定的亲缘关系[32]。本研究所涉及的43个物种染色体核型与前人也有一定的差异。除V.tenuifoliasubsp.tenuifolia的核型数据外,其余42份种质的核型数据已有报道,与本研究的核型数据存在一定差异,原因可能是,由于预处理的方法和试剂不同,染色体数目与现有报道一致,但核型公式有差别:如Arslan等[33]研究表明V.hybrida的核型公式2n=2x=14=1m+4sm+1a+1st。管超等[19]对春箭筈豌豆的V.sativa2505、2556和2560进行染色体分析发现:V.sativa2505的核型公式为2n=2x=12=8m+2sm+2st;春箭2556和2560的核型公式为2n=2x=12=10m+2sm。以上材料与本研究观察染色体数目相同,但核型公式有差异,本研究观察到V.hybrida的核型公式为2n=2x=14=4m+8sm+2st;V.sativa2505的核型公式为2n=2x=12=8m+4sm;V.sativa2556和V.sativa2560的核型公式分别是:2n=2x=12=8m+4sm和2n=2x=12=6m+6sm。刘玉红[34]发现V.unijuga的核型公式2n=12=4m+4sm+4st,第6对短臂具随体。与本研究观察到的V.unijuga核型公式2n=2x=12=6sm+6st有差异,未发现随体;山野豌豆(V.amoena)核型公式2n=24=6m+12sm+6st,第8、12对染色体均有随体。与本研究所观察的染色体数目一致2n=4x=24=22sm+2st,核型公式有差异,未发现随体。
由于材料来源和栽培技术不同引起变异,也会造成染色体数目和结构发生改变。如Gaffarzadeh等[35]研究表明V.villosa的染色体数目2n=2x=14;本研究中V.villosa的染色体数目2n=2x=12。Laghetti等[36]研究表明V.articulata的染色体数目2n=2x=14;本研究中V.articulata的染色体数目2n=2x=10。Galasso等[37]研究表明V.benghalensis的染色体数目2n=2x=14;本研究中V.benghalensis的染色体数目2n=2x=12。Li等[38]研究表明V.cracca的染色体数目2n=2x=12;本研究中V.cracca的染色体数目2n=2x=10。Schifino等[39]研究表明V.hirsuta和V.pannonica的染色体数目分别是2n=2x=14和2n=2x=12;本研究中V.hirsuta和V.pannonica的染色体数目2n=2x=12和2n=2x=14。Castiglione等[15]观察V.lathyroides和V.grandiflora的染色体数目分别是2n=2x=12和2n=2x=14。本研究中V.lathyroides及V.grandiflora的染色体数目2n=2x=10和2n=2x=12。
不同品种的核型均会出现不同程度的差异,主要有染色体臂比值、不对称系数、核型公式等方面,原因可能是野豌豆属植物邻近染色体间长短、形态差异不显著,造成染色体的配对和排列不同;也可能是在试验过程中的取材,处理方法和压片技术等造成的差异[30-32]。染色体数目减少的机制很可能是由于染色体发生易位和融合[18]。
Stebbins[25]认为核型对称性与物种进化相辅相成,核型对称性越高的生物,染色体变异较小,物种进化程度相对较低;核型不对称程度高,染色体变异大,进化程度较高。本研究中V.pannonica和V.peregrian的染色体核型为2A型,核型对称性明显,表明这两个物种在系统演化过程中居于较原始的地位;而V.fabavar.minuta、V.lutea、V.sativasubsp.nigra的染色体核型为3C型,核型不对称性明显,属于相对进化的类型。
致谢:该论文是第二届全国草业生物技术大会评选出的优秀论文,并得到中国草业生物技术专业委员会提供的版面费支持。
[1] Castiglione M R,Gelati M T,Frediani M,Ravalli C,Venora G,Caputo P,Cremonini R.Cytology ofViciaspecies X.karyotype evolution and phylogenetic implication inViciaspecies of the sectionsAtossa,Microcarinae,WiggersiaandVicia[J].Protoplasm,2011,248(4):707-716.
[2] Ball P W.Flora Europaea[M].Cambridge:Cambridge University,1968:129-136.
[3] Castiglione M R,Frediani M,Ravalli C,Venora G,Cremonini R.Cytological characterization ofViciaoroboidesWulfen in Jacq[J].Protoplasm,2009,236(1):21-27.
[4] Caputo P,Frediani M,Venora G,Ravalli C,Ambrosio M,Cremonini R.Nuclear DNA contents,rDNAs and karyotype evolution in subgenusVicia:Ⅲ.The heterogeneous sectionHypechusa[J].Protoplasm,2006,228(4):167-177.
[5] Frediani M,Caputo P,Venora G,Ravalli C,Ambrosio M,Cremonini R.Nuclear DNA contents,rDNAs and karyotype evolution inViciasubgenusVicia:Ⅱ.SectionPeregrinae[J].Protoplasm,2005,226(3):181-190.
[6] Maxted N.A phenetic investigation ofViciaL. subgenusVicia(Leguminosae,Vicieae)[J].Botanical Journal of the Linnean Society,1993,111(2):155-182.
[7] Liu Z P,Ma L C,Nan Z B,Wang Y R.Comparative transcriptional profiling provides insights into the evolution and development of the zygomorphic flower ofViciasativa(Papilionoideae)[J].PLoS One,2013,8(2):e57338.
[8] Liu Z P,Liu P,Luo D,Liu W X,Wang Y R.Exploiting illumina sequencing for the development of 95 novel polymorphic EST-SSR markers in common vetch (Viciasativasubsp.sativa)[J].Molecules,2014,19(5):5777-5789.
[9] 南志标,张吉宇,王彦荣,李春杰,聂斌,张建全,赵宏.五个箭筈豌豆品系基因型与环境互作效应及农艺性状稳定性[J].生态学报,2004,24(3):395-401.
[10] 贺学礼,赵莉丽,谭志远,叶秦,朱铭娥,陈丽娟.7种豆科植物染色体形态的研究[J].中国草地,1994(3):62-64.
[11] 姚雅琴,蒋选利,郁枫.豆科Leguminosae植物部分种染色体形态研究[J].西北农业大学学报,1990,18(2):78-82.
[12] 刘鹏,马利超,王宇,刘志鹏.野豌豆属牧草种质花粉形态的扫描电镜观察[J].草业科学,2013,30(11):1755-1761.
[13] 夏振岱.中国野豌豆属的分类研究[J].植物分类学报,1996,34(4):421-433.
[14] 宋敏,于洪柱,娄玉洁,陈涛,徐安凯.山野豌豆生物学特性及其利用[J].草业与畜牧,2011(4):5-6.
[15] Castiglione M R,Gelati M T,Frediani M,Gelati M T,Venora G,Giorgetti L,Caputo P,Cremonini R.Cytological and molecular characterization ofViciabarbazitaeTen. & Guss[J].Protoplasma,2012,249(3):779-788.
[16] Caputo P,Frediani M,Gelati M T,Venora G,Crremonini R,Castiglione M R.Karyological and molecular characterisation of subgenusVicia(Fabaceae)[J].Plant Biosystem,2013,147(4):1242-1552.
[17] 苏坤梅,杨德奎.山野豌豆的染色体数目及核型分析[J].山东科学,2001,14(3):48-50.
[18] 冉雪琴,朱晓彤,朱邦长.贵州野豌豆属植物的核型研究[J].草业科学,1997,14(1):5-9.
[19] 管超,张吉宇,王彦荣,聂斌.野豌豆属4个品种(系)的染色体核型[J].草业科学,2012,29(10):1540-1545.
[20] 蔡利娟,周娅,周兰英,蒲光兰,马诗钰.9种松属植物的核型及亲缘关系[J].东北林业大学学报,2014,42(2):57-60.
[21] 赵振军,黎星辉.阿拉伯茶染色体核型分析[J].长江大学学报,2014,11(5):17-21.
[22] 李懋学,张赞阳.作物染色体及其研究技术[M].北京:中国农业出版社,1996:292-297.
[23] 李璇,段青,王祥宁,崔光芬,贾文杰,马璐琳,蒋亚莲,王继华,吴丽芳.云南泸定百合12个野生居群的核型分析研究[J].园艺学报,2014,41(5):935-945.
[24] 李懋学,陈瑞阳.关于植物核型的标准化问题[J].武汉植物学研究,1985,3(4):297-302.
[25] Stebbins G L.Chromosomal Evolution in Higher Plants[M].London:Edward Amold,1971:85-105.
[26] Arano H.Cytological studies in subfamily Carduoideae(Compositae) of Japan[J].Botanical Magazine of Tokyo,1963,76:32-39.
[27] 杜荣骞.生物统计学[M].北京:高等教育出版社,1990.
[28] 胡凤荣,王斐,鲍仁蕾,谢蔚然.风信子根尖预处理及核型分析[J].西北植物学报,2012,32(10):2030-2034.
[29] 张自强,于肖夏,鞠天华,于卓,蒙美莲,王丹,姜超,崔阔澍.3个马铃薯杂种优良株系的核型及SSR分析[J].西北植物学报,2014,34(7):1318-1324.
[30] Fritsch R M,Blattner F R,Gurushidze M.New classification ofAlliumL. subg.Melanocrommyum(Webb & Berthel) Rouy (Alliaceae) based on molecular and morphological characters[J].Phyton,2010,49(2):145-220.
[31] 张红梅,吴鹏,蔡宝宏,李思颖,杨文平,李明.玻璃苣染色体核型分析[J].生物学通报,2014,49(5):47-49.
[32] 唐建敏,汪婷,郭雅丹,曾艳,唐茜.荥经琵琶茶野生大茶树染色体核型分析[J].贵州农业科学,2014,42(1):12-15.
[34] 刘玉红.野豌豆属五种牧草的核型分析[J].中国草地学报,1986(5):38-43.
[35] Gaffarzadeh L,Badrzadeh M,Asghari R.Karyotype of severalViciaspecies from Iran[J].Asian Journal of Plant Science,2008,7(4):417-420.
[36] Laghetti G,Piergiovanni A R,Galasso T,Hammer K,Perrino P.Single-flowered vetch (ViciaarticulataHornem.):A relic crop in Italy[J].Genetic Resources and Crop Evolution,2000,47(4):461-465.
[37] Galasso I,Sonnante G,Tota D G,Pignone D.Comparison of molecular cytogenetic and genetic analyses in accession of the two biotypes ofViciabenghalensisL.[J].Annals of Botany,1997,79(3):311-317.
[38] Li R J,Taylor S,Jenkins G.Unravelling the phylogeny of tetraploidViciaamoena(Fabaceae) and its diploid relatives using chromosomal landmarks[J].Hereditas,2001,134(3):219-224.
[39] Schifino M T,Lau A H,Simioni C.The generaViciaandLathyrus(Leguminosae) in Rio Grande do Sul (Southern Brazil):Cytogenetics of native,naturalized and exotic species[J].Revista Brasileira de Genetica,1994,17(3):313-319.
(责任编辑 王芳)
2015年5月国际市场主要畜产品与饲料价格分析
5月国际饲料价格普遍下跌,畜产品价格持续上涨。
一、玉米、大豆、棉籽饼、豆粉、豆粕、高梁和苜蓿粉市场价格持续下跌,跌幅较小;菜籽市场价格出现反弹
5月份玉米、大豆、棉籽饼、豆粉、豆粕、高梁和苜蓿粉市场平均价格分别为141.42、351.95、320.25、313.66、308.97、136.55和283.00美元·t-1,环比分别下降3.84%、1.30%、2.62%、2.35%、2.42%、3.52%和0.70%。菜籽市场平均价格为426.17美元·t-1,环比上涨2.52%。
二、育肥牛、鸡肉、羊肉、牛肉、猪肉、瘦肉猪和牛奶市场价格持续上涨,羊羔肉市场价格较4月份平稳
5月份育肥牛、鸡肉、羊肉、牛肉、猪肉、瘦肉猪和牛奶市场平均价格分别为4.79、2.56、1.91、5.04、1.89、1.79和0.28美元·kg-1,环比分别上涨0.37%、0.20%、8.63%、0.16%、0.92%、26.33%和2.07%。羊羔肉市场平均价格为3.26美元·kg-1,与4月份持平。
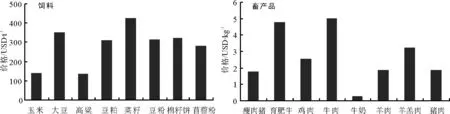
图1 2015年5月国际市场主要饲料与畜产品价格
数据来源:国际市场商品价格网 http://price.mofcom.gov.cn/;中国农业信息http://www.agri.gov.cn/;鸡肉 http://www.indexmundi.com/;羊羔肉、羊肉 http://www.interest.co.nz/rural;牛肉http://www.thebeefsite.com/;猪肉 http://www.thepigsite.com/;货币汇率:http://qq.ip138.com/hl.asp。
(兰州大学草地农业科技学院 王迎新 整理)
Karyotypes analysis of 43Viciaaccessions
LIU Peng, WANG Yan-rong, LIU Zhi-peng
(College of Pastoral Agriculture Science and Technology, Lanzhou University; State Key Laboratory of Grassland Agro-ecosystems, Lanzhou 730020, China)
Most species inViciagenus are important legume crop that are commonly used as windbreak and sand water conservation. It is suitable for cultivation in alpine and high-altitude areas, which supply protein feed for Tibet plateau in China. Here, we collected 43Viciagermplasms from 22 countries such as China, America, Germany and Ethiopia. The chromosomal karyotypes of 43 accessions were analyzed by the root tip cells. The results showed that there were five types of chromosome number: 10, 12, 14, 16 and 24. There are six types of karyotypes: 2A, 1B, 2B, 3B, 2C and 3C. In the present study, the chromasome karyotypes of 43Viciaaccessions were comprehensively calculated which provided the important scientific basis for the researches of cytological characteristics and evolutionary.
Vicia; chromasome; karyotype analysis
LIU Zhi-peng E-mail:lzp@lzu.edu.cn
10.11829j.issn.1001-0629.2014-0593
2014-12-22 接受日期:2015-03-03
国家重点基础研究发展计划(2014CB138704);国家自然科学基金项目(31272492)
刘鹏(1989-),女,甘肃兰州人,在读硕士生,主要从事牧草育种与分子生物学研究。E-mail:liup12@lzu.edu.cn
刘志鹏(1979-),男,陕西武功人,副教授,博士,主要从事牧草育种与分子生物学研究。E-mail:lzp@lzu.edu.cn
S551+.9;Q944
A
1001-0629(2015)06-0908-19
刘鹏,王彦荣,刘志鹏.野豌豆属43份牧草种质的染色体形态观察与分析[J].草业科学,2015,32(6):908-926.LIU Peng,WANG Yan-rong,LIU Zhi-peng.Karyotypes analysis of 43Viciaaccessions[J].Pratacultural Science,2015,32(6):908-926.
