基于叶片解剖结构对青藏高原25种灌木的抗旱性评价
2015-06-23李吉跃王军辉马建伟
潘 昕,邱 权,李吉跃,王军辉,何 茜,苏 艳,马建伟,杜 坤
(1华南农业大学林学院,广东广州510642;2福州市林业局,福建福州350007; 3中国林业科学研究院林业研究所/国家林业局林木培育重点实验室,北京100091; 4甘肃省小陇山林业科学研究所,甘肃天水741022)
基于叶片解剖结构对青藏高原25种灌木的抗旱性评价
潘 昕1,2,邱 权1,李吉跃,王军辉3,何 茜1,苏 艳1,马建伟4,杜 坤4
(1华南农业大学林学院,广东广州510642;2福州市林业局,福建福州350007; 3中国林业科学研究院林业研究所/国家林业局林木培育重点实验室,北京100091; 4甘肃省小陇山林业科学研究所,甘肃天水741022)
【目的】研究青藏高原25种灌木叶片解剖结构和气孔特征,了解其水分适应机制,为青藏高原地区造林筛选优良耐旱植物提供参考依据.【方法】通过常规石蜡切片技术,对比叶片角质层、上下表皮、栅栏组织、海绵组织和叶片厚度等12项指标,运用主成分分析和隶属函数法对供试植物进行抗旱性能分析与评价.【结果和结论】25种植物叶片具有典型的旱生结构,叶片厚度最高为323.00μm、平均为186.25μm,上表皮角质层厚度最高为4.83μm、平均为1.71μm,上表皮平均比下表皮厚30.00%,气孔小而密集,这是植物长期适应高原干旱环境的进化结果,12项指标的种间差异极显著.通过主成分分析法结合各指标的变异系数筛选出5项具有代表性的抗旱指标:气孔密度、叶片厚度、栅栏组织厚度、海绵组织厚度和角质层厚度,运用隶属函数值法对各树种的抗旱性能评分排序,将25种植物分为抗旱性极强树种、抗旱性较强树种、抗旱性一般树种和抗旱性较弱树种.
灌木;叶片解剖结构;气孔特征;抗旱性;干旱胁迫;青藏高原
植物的抗旱能力是在长期受到水分胁迫后通过遗传变异和自然选择而逐渐形成的.这种胁迫抗性与植物的内部结构、生理状况有密切的关系[1].青藏高原平均海拔4 000 m以上,被称为世界的第三极[2],具有太阳辐射强、日照时间长、湿度分布不均衡、低温等独特的高原气候特点,降水分布表现出明显的地域性和季节性差异,降水量主要受西南季风控制,东南部年均降水量>400 mm,西北部不足100 mm,植被从东南到西北随自然条件的水平、垂直及坡向变化,依次出现森林、草甸、草原和荒漠[3-5].因此,选择适宜在青藏高原环境下生长的耐旱树种具有重要意义.
叶片是高等植物进行光合作用的重要器官,也是植物进行气体交换、水分蒸腾、运输的门户,其形态结构特征直接影响到植物的生理活动和生态功能.叶片对环境因子如降水、温度、海拔等的变化最为敏感,具有较大的变异性和可塑性[6-7],且叶表皮特征和解剖结构已被广泛应用于系统分类学的研究和种属间亲缘关系的确定,并与植物的抗旱性呈一定的相关性[8-11].严酷的气候条件下,由于受环境的胁迫植物叶片的形态、结构以及生理功能等方面,保留了一些与该环境相适应的特殊解剖特征和生理机制[12].近年来,有关植物叶片解剖结构与其抗旱性的研究已有不少,赵庆芳等[13]利用扫描电镜技术和石蜡切片法对青藏高原东部嵩草属13种植物叶结构进行了研究,对其生态适应特性进行了探讨.何涛等[14]从形态和解剖结构两方面对青藏高原高山植物的研究进展进行了综述,重点阐述了高山植物的形态、解剖结构及其与环境的适应性关系.马建静等[15]对青藏高原高寒草地和内蒙古高原温带草地65种双子叶植物叶片解剖特征进行了研究,分析了植物对气候的适应特征.但以青藏高原灌木为对象的研究在植物种类和研究宽度上都有很大的不足,且在叶片解剖和气孔特征两方面的共同分析较为缺乏.因此,本试验以红砂Reaumuria songarica、唐古特白刺Nitraria tangutorum、俄罗斯大果沙棘Hippophae rhamnoides等25种青藏高原灌木为试材,对其叶片的解剖结构和气孔特征进行观察分析,从叶片的结构特征角度对树种的抗旱性、生态适应性进行系统研究和评价,为青藏高原干旱地区灌木树种抗旱评价指标的建立及抗旱树种的筛选提供可靠的基础数据和科学依据.
1 材料与方法
1.1 苗木种植试验地概况
苗木种植试验地设在甘肃省天水市小陇山林业科学研究所,其位于秦岭北坡,渭河支流川台区,地理纬度105°54'37″E、34°28'50″N,平均海拔1 160 m.年降水量600~800 mm,年均蒸发量1 290 mm,年均气温10.7℃,≥10℃年积温3 359.0℃,极端高温39℃,极端低温-19.2℃.无霜期约190 d.土壤中性偏碱,pH7.2.
1.2 研究材料
选择青藏高原25种灌木:红砂、西北沼委陵菜Comarum salesovianum、俄罗斯大果沙棘、唐古特白刺、胡颓子Elaeagnus pungens、唐古特莸 Caryopteris tangutica、西北小檗Berberiskawakamii、肋果沙棘Hippophae neurocarpa、鲜卑花Sibiraea laevigata、树锦鸡儿Caragana arborescens、珍珠梅Sorbaria kirilowii、金露梅Potentilla fruticosa、锦鸡儿Caragana sinica、尼泊尔锦鸡儿Caragana nepalensis、鲜黄小檗Berberis diaphana、绣线菊Spiraea salicifolia、三颗针Berberis julianae、霸王 Sarcozygium xanthoxylon、红花岩黄芪Hedysarum multijugum、柳叶沙棘Hippophae salicifolia、白柠条Caragana korshinskii、蔷薇Rosa multiflora、江孜沙棘 Hippophae rhamnoides、西藏沙棘 Hippophae thibetana、短叶锦鸡儿Caragana brevifolia为试材,于2010年4月播种,每种灌木播种10株,种植于小陇
山林业科学研究所苗圃大田试验地,并定期对其进行浇水、除草等日常养护管理.
1.3 测定指标与方法
1.3.1 叶片解剖结构 2012年7月下旬对25种树种的2年生播种苗进行取样,每个树种选取长势相近的3株,取树冠中上部南向枝条上的功能叶20片进行试验.取样时间为晴天上午9:00—11:00,便于观察气孔结构.剪取叶片主脉中间0.5 cm×0.5 cm的小块用FAA(Formalin-acetic acid-alcohol,标准固定液)固定,采用常规石蜡切片法[16].每个树种3株重复,每个重复制成永久装片10张,在 Olympus DP72显微镜下观察,并用Motic Image Advanced3.0软件测量叶片总厚度、上下表皮厚度、角质层厚度、栅栏组织厚度、海绵组织厚度等指标,每个视野重复30次.

1.3.2 气孔特征 采样方法同1.3.1,并用FAA固定液固定.之后将小叶浸泡在离析液中(体积分数为30%的过氧化氢与乙酸的体积比为1∶1的混合液),于60℃恒温箱中放6~9 h后,待叶片变白,叶肉组织与表皮细胞分离后,用蒸馏水冲洗干净,剥取叶表皮.再用质量分数为1%的番红染色5~10 min,依次经乙醇(体积分数分别为 70%、85%、95%和100%)、1/2无水乙醇+1/2二甲苯、二甲苯逐级脱水,最后用中性树胶封片.在Olympus DP72显微镜下观察,并用Motic Image Advanced3.0软件测量气孔长度、气孔宽度、气孔密度等指标.每个视野重复15次[17].
1.3.3 数据处理 对25种灌木的若干项典型指标的隶属值进行累加,求平均值.如果指标与抗旱性呈正相关:X(μ) =(X-Xmin)/(Xmax-Xmin);如果指标与抗旱性呈负相关:1-X(μ)=1-(X-Xmin)/ (Xmax-Xmin).式中μ为树种代号,X(μ)表示该树种的隶属函数值,X表示各指标的测定值,Xmax和Xmin分别表示各灌木种对应指标的最大和最小测定值.
试验中的数据统计由Microsoft excel完成,数据处理由SPSS9.0软件中Duncan's法的方差分析、多重比较、主成分分析方法完成.
2 结果与分析
2.1 叶片解剖结构特征比较
叶片是植物暴露在地面环境中最多的器官,其形态结构特征最能体现植物对环境的适应,叶片小而厚有利于减少叶片的蒸腾面积并提高其控水能力.由表1可以看出,25种灌木的叶片厚度107.63~323.00μm,平均为186.25μm,变异系数与其他指标相比较小,为32.81%.其中霸王的叶片最厚,为323.00μm,其次是俄罗斯大果沙棘(299.65μm),有11个树种达到200μm以上.短叶锦鸡儿叶片最薄,为107.63μm,其次是江孜沙棘,为116.30μm.叶片上表皮细胞壁外附有1层0.78~4.83μm的角质层,是1层不透水的脂肪性物质,主要防止水分散失,其平均厚度为1.71μm,角质层的变异系数与其他指标相比最大,为59.72%,可见角质层厚度对树种间抗旱能力的比较具有重要意义,通常角质层厚度越大,表明其抗旱性越强.其中,尼泊尔锦鸡儿的角质层最厚,为4.83μm,其次是锦鸡儿(3.15μm)、珍珠梅(3.11μm),其余树种的角质层厚度均低于3.00μm,西北小檗最薄,为0.78μm.
表皮对植物起到了保护、贮水的作用,植物表皮越厚抗旱性越强.25个树种中除唐古特莸、鲜卑花、树锦鸡儿、锦鸡儿、红花岩黄芪、白柠条外,其余树种的上表皮均厚于下表皮.上表皮的平均厚度为14.74 μm,细胞较厚,下表皮的平均厚度为11.34μm,相对较薄,各树种上、下表皮厚度之间存在极显著差异(P<0.01).其中,上表皮最厚的为唐古特白刺(28.60μm),最薄的为白柠条(7.63μm),其相差2.75倍,只有俄罗斯大果沙棘、唐古特白刺、金露梅3个树种的上表皮厚度大于20.00μm.下表皮方面,唐古特白刺最厚,为18.09μm,霸王最薄,只有5.73 μm,其次为短叶锦鸡儿、俄罗斯大果沙棘、蔷薇,均低于7.00μm.金露梅的上、下表皮差异最大,达到15.11μm,胡颓子上、下表皮厚度最接近,上表皮仅比下表皮厚0.99μm.
栅栏组织有助于植物减少水分的蒸腾,提高植物的光合效能以及抗旱能力,海绵组织的增加是植物对低温环境的反应.本试验中,各供试材料的栅栏组织、海绵组织厚度差异较大,部分树种的海绵组织比栅栏组织厚.其中俄罗斯大果沙棘的栅栏组织最厚,为165.71μm,江孜沙棘最薄,为39.62μm.海绵组织中霸王最厚(205.20μm),树锦鸡儿最薄(35.57μm).叶肉中的栅栏组织与海绵组织的比值(简称栅海比)也是评价植物抗旱性的重要指标之一.25个树种中栅海比平均为1.21,树锦鸡儿最大,为2.22,其次为锦鸡儿(2.13),且仅有此2个树种的栅海比大于2.0,唐古特莸最小,仅为0.40,大部分树种的栅海比集中在1.90~0.70.
叶片组织结构紧密度和疏松度是表示叶片栅栏组织、海绵组织于叶片厚度的重要指标,对抗旱性也有重要的意义.叶片组织结构紧密度平均为42.52%,疏松度为41.11%,两者较接近.其中叶片组织结构紧密度最大为俄罗斯大果沙棘(55.64%),最小为唐古特莸24.68%,二者相差1.25倍.其余树种中,西北沼委陵菜、胡颓子、树锦鸡儿、珍珠梅、锦鸡儿的紧密度也超过50%;而叶片组织结构疏松度最大为霸王(63.36%),其叶片厚度中海绵组织已过半,栅栏组织较不发达,锦鸡儿的疏松度最小,为25.29%,此外只有树锦鸡儿、金露梅的疏松度低于30.00%.
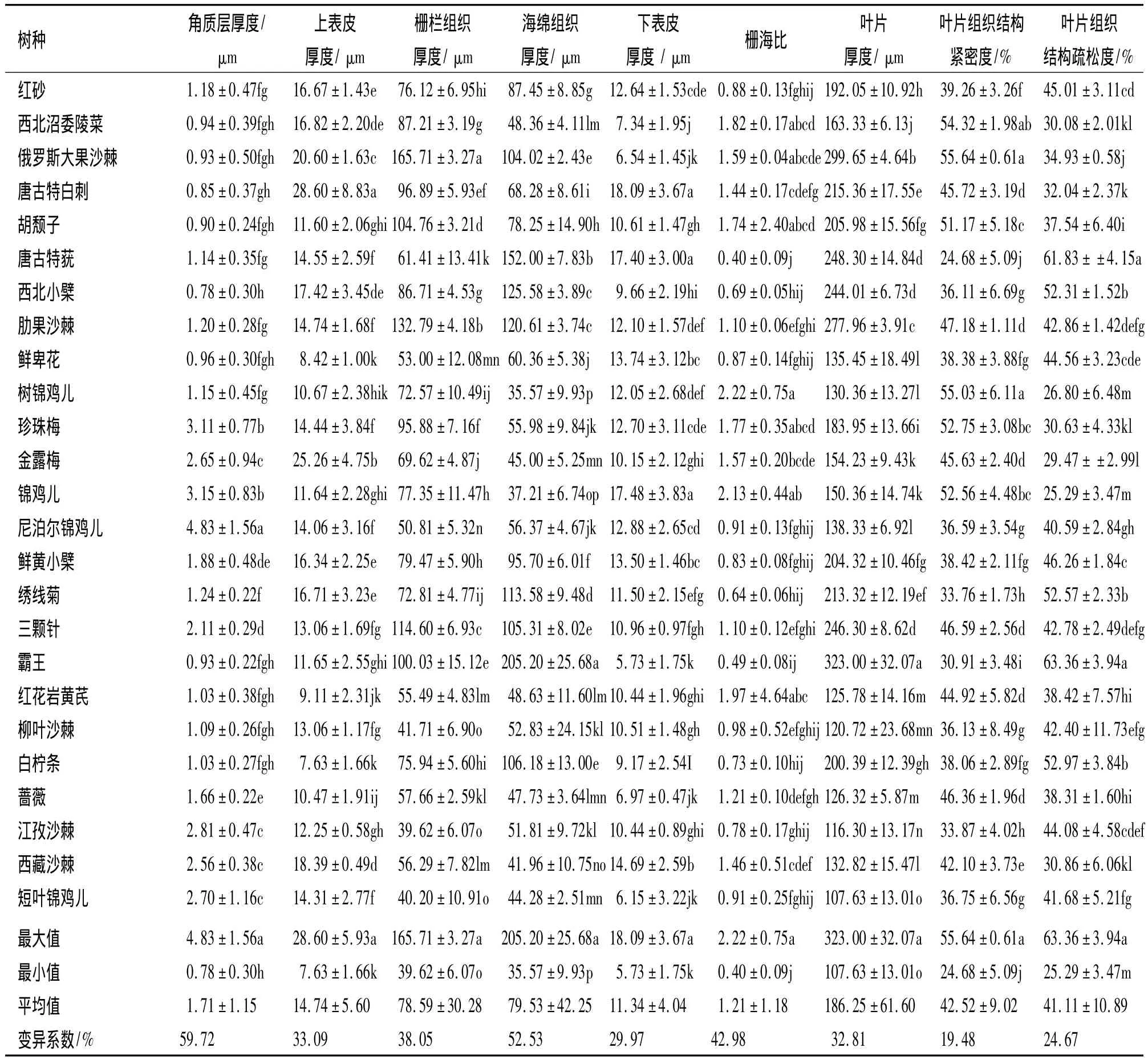
表1 25种灌木叶片解剖结构特征1)Tab.1 Leaf anatom ical structure characteristics of 25 shrub species
2.2 叶片气孔特征比较
叶片上控制蒸腾耗水的主要是气孔,一般认为气孔较小密度较大的植物在干旱条件下能有效地减少水分散失.由表2可知,25种灌木的气孔长度为6.47~24.29μm,平均为14.37μm,三颗针的气孔长度最大,其次为鲜黄小檗(22.26μm)、西北小檗(22.03μm),且仅有此3个树种的气孔长度大于20 μm,柳叶沙棘的气孔长度最小,只有三颗针的26.63%,金露梅(6.83μm)和江孜沙棘(8.21μm)的气孔长度也很小.气孔宽度方面,鲜黄小檗的气孔宽度最大(18.65μm),金露梅最小(2.37μm),平均为8.31μm,变异系数达到52.55%.其中气孔宽度大于10.00μm的共有7个树种,分别为鲜黄小檗、西北小檗(15.80μm)、树锦鸡儿(15.34μm)、蔷薇(14.33μm)、三颗针(12.24μm)、尼泊尔锦鸡儿(12.04μm)、唐古特白刺(11.03μm).种间平均气孔密度为145.77个·mm-2,范围57.27~360.05个·mm-2,变异系数高达52.09%.其中超过300个·mm-2的仅有胡颓子,超过200个·mm-2的有鲜卑花、蔷薇、红花岩黄芪和鲜黄小檗.气孔密度相对较小的是西北沼委陵菜(62.58个·mm-2)、唐古特莸(57.27个·mm-2)、树锦鸡儿(62.50个·mm-2).

表2 25种灌木叶片气孔特征1)Tab.2 Leaf stomatal structure characteristics of 25 shrub species
2.3 叶片结构指标的抗旱性主成分分析
综合叶解剖特征和表皮形态,25种灌木的12项指标得到的排列顺序均不同.因此,为综合评判各个树种叶片旱生构造的优劣,对以上12项叶片特征指标进行主成分分析,采用量化指标以评定植物的抗旱性[18],主成分系数绝对值较大的指标可作为评价树种抗旱性的重要依据.主成分分析结果(表3)表明,前2个主成分的累计贡献率已达到92.92%,第1主成分中气孔密度、叶片厚度的系数较大,栅栏组织、海绵组织厚度次之,第2主成分中,依旧是气孔密度、叶片厚度的系数较大,栅栏组织、海绵组织厚度次之,因此选择气孔密度、叶片厚度作为评价供试树种抗旱性的主要指标,栅栏组织、海绵组织厚度作为次要指标.
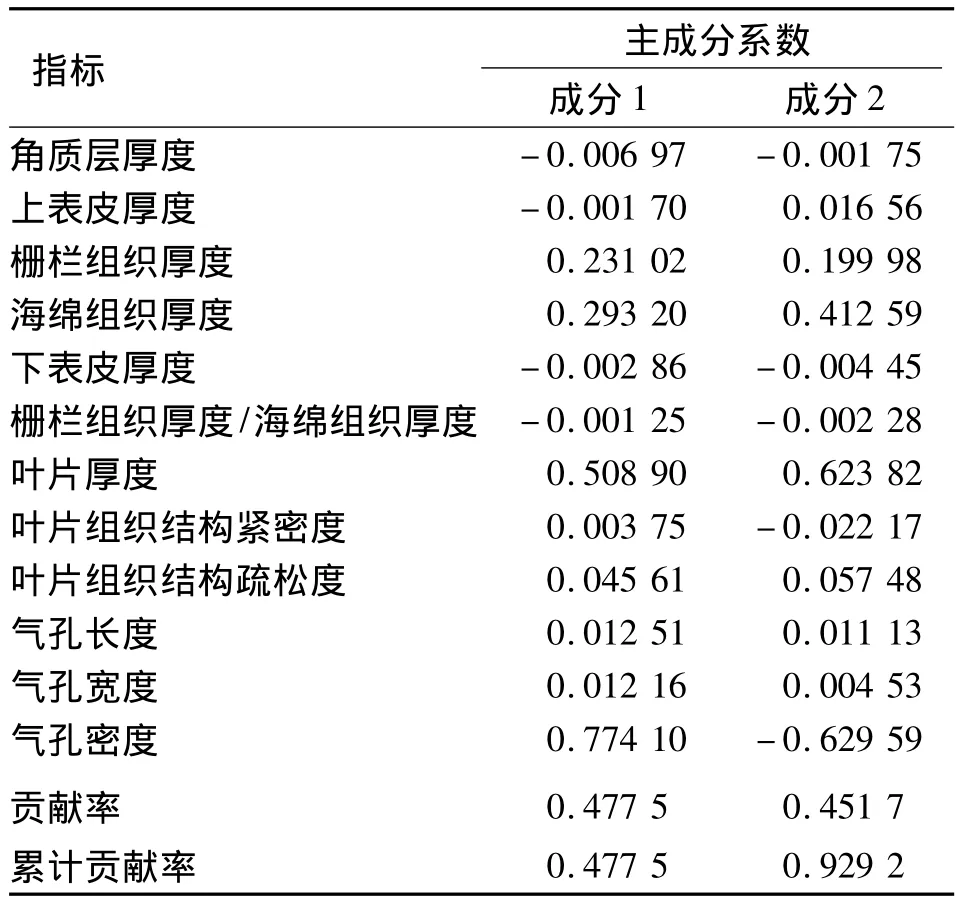
表3 叶片解剖结构特征的抗旱性主成分分析Tab.3 Results of a p rincip le com ponent analysis of leaf characteristics of drought resistance
2.4 种间抗旱性比较
从表1、2中可以看出各指标的变异系数范围19.48%~59.72%,变异系数最大的为角质层厚度,一般在一组指标中,变异系数大的指标对分析结果具有重要的意义[19],因此,结合主成分分析结果,选取叶片厚度、气孔密度、栅栏组织厚度、海绵组织厚度、角质层厚度5项指标,运用隶属函数法进行比较[20].对25种灌木的5项典型指标的隶属值进行累加求平均值,平均值越大抗旱性越强.
运用隶属函数法综合评价了25种树种的抗旱能力并对其进行排序(表4),平均值越大的树种抗旱性越强.因此25种灌木中俄罗斯大果沙棘的抗旱性最强,唐古特莸的抗旱性最弱.由于树种较多,所以依据隶属函数的均值以及植物在叶片解剖结构和气孔结构的特征,特别是叶片厚度、气孔密度、栅栏组织厚度、海绵组织厚度、角质层厚度5项指标,人为地将所有树种分为四大类,以便归纳、对比和总结.第1类为抗旱性极强的树种,具有叶片较厚、气孔密度大的特点,包括俄罗斯大果沙棘、胡颓子、三颗针;第2类为抗旱性较强的树种,主要表现为栅栏组织较厚,包括:肋果沙棘、珍珠梅、锦鸡儿、鲜黄小檗、唐古特白刺、尼泊尔锦鸡儿;第3类为抗旱性一般的树种,各项指标都表现为一般水平,且树种最多,包括:蔷薇、金露梅、鲜卑花、西藏沙棘、绣线菊、红花岩黄芪、西北小檗、红砂、江孜沙棘、白柠条、西北沼委陵菜、霸王、短叶锦鸡儿;第4类为抗旱性较差的树种,其海绵组织较厚,栅栏组织较薄,且气孔密度较低,平均为78.94个·mm-2,树种有:树锦鸡儿、柳叶沙棘、唐古特莸.
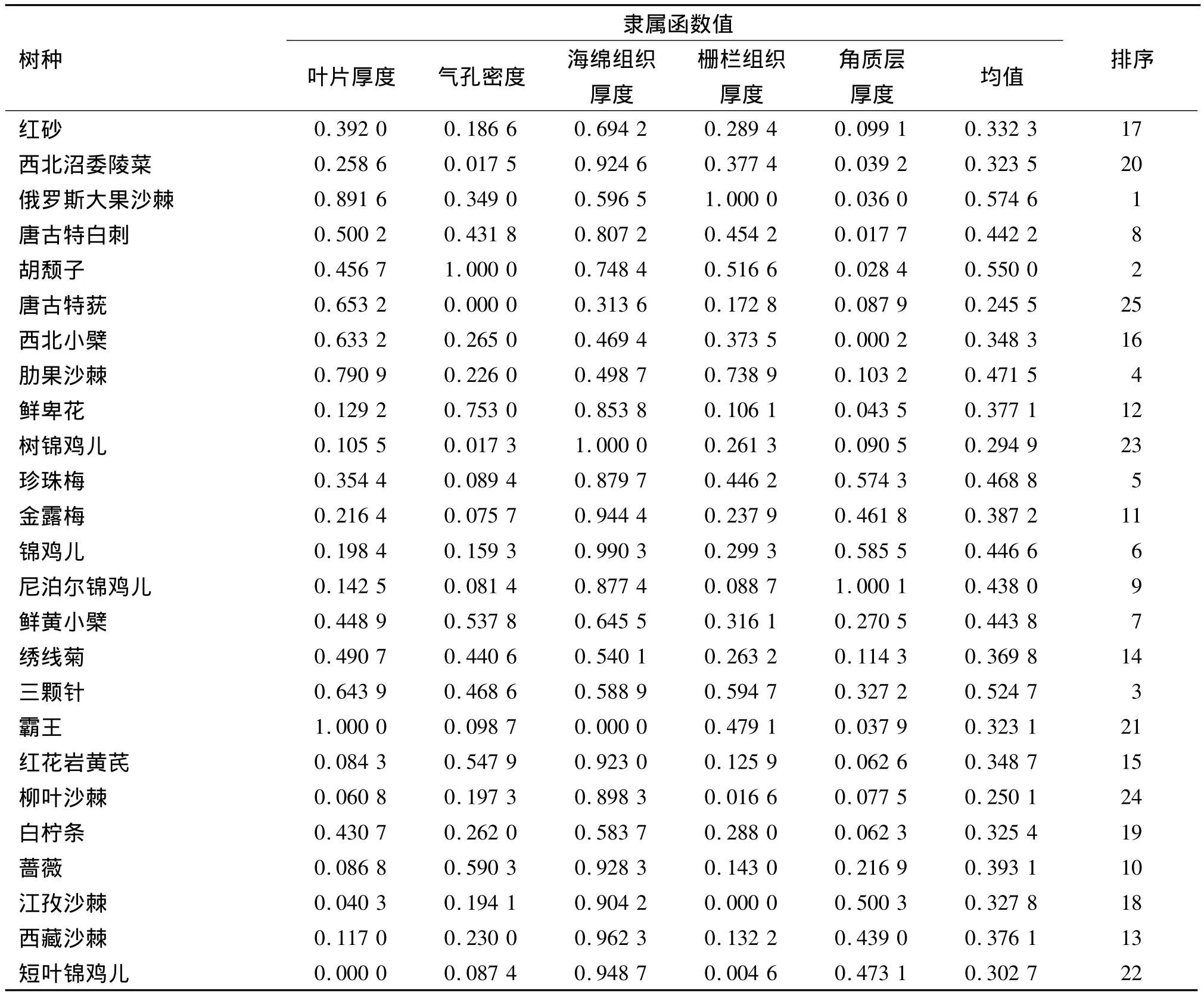
表4 25种灌木5项指标的隶属函数值及抗旱性综合评价结果Tab.4 Subordinate function values of 25 shrub species and comprehensive appraisals of drought resistance
3 讨论与结论
叶片是植物进化中对环境变化较敏感、可塑性较大的器官,其结构特征最能体现环境因子的影响或植物对环境的适应[21],因而叶片的解剖结构特征是反映植物对干旱生境适应能力非常重要的一个方面,是树木抗旱性综合评定必不可少的一部分[22].青藏高原昼夜温差大、太阳辐射强、空气干燥,植物极易受到生理性干旱的胁迫,其叶片必然形成与之相适应的形态结构[23].如表皮、角质层较厚,有的植物有表皮毛,叶片气孔小而密度大等特征,具有隔热、保水的功能,是植物对强光、低温、缺水等环境的适应[24-26].
本试验对25种灌木的叶解剖特征研究表明,供试灌木叶面积较小,上表皮角质层平均厚度为1.71 μm,最大值高达4.83μm,叶表皮气孔小且密集,平均密度为141.85个·mm-2,这样的叶片结构能够有效减少水分散失,增强植物叶片控水力,通气组织发达,将白天产生的氧气和夜晚放出的二氧化碳进行充分利用.李春阳等[27]也得出旱生植物具有机械组织较为发达、表皮往往有多层细胞、角质层发达、表皮毛密集分布及气孔下陷等特征以减少水分散失的结论.但张晓艳等[28]也提出气孔密度并不随着干旱程度的加剧而一直增加.试验的25种植物上表皮平均比下表皮厚30.0%.叶肉组织普遍较厚,叶厚度变异系数较小,有利于减少叶片蒸腾面积并提高贮水性,这也可能与植物所生长的海拔高度有关.叶绿体体积大且数量多,这与崔宏安等[29]对一球悬铃木叶结构的解剖研究结果一致.
由于青藏高原每年适合植物生长的时间有限,所以青藏高原的植物进化出高光合效率的叶片结构.本试验中植物叶肉细胞体积较小,叶绿体多,增加了光合反应的效率和反应面积.栅栏组织层数多、排列紧密,在光合作用中提高了对水分、光能的利用效率,弥补叶片狭小的负面影响,这一点在崔秀萍等[30]对浑善达克沙地不同生境下黄柳Salix gordejevii叶片解剖结构的比较试验中已得到证实.同时海绵组织可能也是对青藏高原低温环境适应的进化结果.寇建村等[31]对不同紫花苜蓿Medicago sativa品种叶片旱生结构的比较中提出,栅海比、紧密度越高,植物抗旱能力越强,这也与本试验结果类似.本试验的25种植物栅海比平均为1.21,栅海比0.8以下的仅有4种;叶片组织结构紧密度平均为42.52%,低于30.00%的仅有1种,表明青藏高原植物已形成了适应强光照、低温、干旱环境的叶片组织结构.海绵组织的变异系数高达52.53%,这可能是由于不同种植物对低温、生理干旱采取了不同的响应对策的结果.
试验中25种植物总体抗旱性较强,能较好地适应干旱地区的干旱胁迫逆境.通过主成分分析法和对比变异系数可知,叶片厚度、气孔密度、栅栏组织厚度、海绵组织厚度、角质层厚度是反映抗旱能力较为敏感的解剖性状,说明这些叶片结构形态指标均可作为研究植物叶片抗旱性的有效指标.但植物的抗旱性是其对干旱长期适应的一种复杂的综合遗传特性,不仅与自身形态解剖结构特征有关,而且取决于内部的多种生理生化活动,也受到外界自然环境条件的影响,而且在植物的发育阶段或生长季节里,其表现出来的水分生理特性可能也不尽相同.具体评价某种树木的抗旱性还需考虑多方面的因素,从而作出科学的解答.本试验采用隶属函数值法对各树种的抗旱能力进行综合评价,并根据各树种得分将25种灌木树种分为抗旱性极强树种、抗旱性较强树种、抗旱性一般树种和抗旱性较弱树种.
致谢:感谢华南农业大学林学院王博、郑舒鹏、杨思伟、谭锦芬、万紫荣、刘珊、杨宛源同学在石蜡切片试验中给予的大力帮助,特致谢忱!
[1] 孙景宽,张文辉,陆兆华,等.沙枣(Elaeagnus angustifolia)和孩儿拳头(Grewia biloba G.Don var.parvif lora)幼苗气体交换特征与保护酶对干旱胁迫的响应[J].生态学报,2009,29(3):1330-1340.
[2] 张金霞,曹广民,周党卫,等.高寒矮嵩草草甸大气-土壤-植被-动物系统碳素储量及碳素循环[J].生态学报,2003,23(4):627-634.
[3] 刘军会,高吉喜,王文杰.青藏高原植被覆盖变化及其与气候变化的关系[J].山地学报,2013,31(2): 234-242.
[4] 李东明,郭正刚,安黎哲.青藏高原多年冻土区不同草地生态系统恢复能力评价[J].应用生态学报,2008,19(10):2182-2188.
[5] 于格,鲁春霞,谢高地.青藏高原草地生态系统服务功能的季节动态变化[J].应用生态学报,2007,18 (1):47-51.
[6] 李全发,王宝娟,安丽华,等.青藏高原草地植物叶解剖特征[J].生态学报,2013,33(7):2062-2070.
[7] 逯永满,姜彦成.中国海罂粟属(Glaucium L.)叶片特征及其抗旱性[J].新疆农业科学,2010,47(10): 2063-2067.
[8] 董蕾,曹洪麟,叶万辉,等.5种喀斯特生境植物叶片解剖结构特征[J].应用与环境生物学报,2011,17 (5):747-749.
[9] KONG H Z.Comparativemorphology of leaf epidermis in the Chloranthaceae[J].Bot J Linn Soc,2001,136(3): 281-296.
[10]陆嘉惠,李学禹,周玲玲,等.甘草属植物叶表皮特征及其系统学意义[J].云南植物研究,2005,27(5): 525-533.
[11]钟义,夏念和.国产润楠属植物的叶表皮特征及其系统学意义[J].热带亚热带植物学报,2010,18(2): 109-121.
[12]BOSABALIDISA M,KOFIDISG.Comparative effects of drought stress on leaf anatomy of two olive cultivars[J].Plant Sci,2002,163(2):375-379.
[13]赵庆芳,崔艳,马世荣,等.青藏高原东部嵩草属植物叶解剖结构的生态适应研究[J].广西植物,2007,27 (6):821-825.
[14]何涛,吴学明,贾敬芬.青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展[J].生态学报,2007,27(6):2574-2583.
[15]马建静,吉成均,韩梅,等.青藏高原高寒草地和内蒙古高原温带草地主要双子叶植物叶片解剖特征的比较研究[J].中国科学:生命科学,2012,42(2):158-172.
[16]周仪.植物形态解剖实验[M].修订版.北京:北京师范大学出版社,1993:37-68.
[17]孙同兴,江幸山.简便有效的叶表皮离析方法:过氧化氢-醋酸法[J].广西植物,2009,29(1):44-47.
[18]何平.数据统计与多元统计[M].西安:西安交通大学出版社,2004:176.
[19]韩刚,李少雄,徐鹏,等.6种灌木叶片解剖结构的抗旱性分析[J].西北林学院学报,2006,21(4):43-46.
[20]王彩华,宋连天.模糊论方法学[M].北京:中国建筑工业出版社,1998:138-203.
[21]王勋陵,马骥.从旱生植物叶结构探讨其生态适应的多样性[J].生态学报,1999,19(6):787-792.
[22]董建芳,李春红,刘果厚,等.内蒙古6种沙生柳树叶片解剖结构的抗旱性分析[J].中国沙漠,2009,29 (3):480-484.
[23]江川,罗大庆,王立辉.西藏半干旱区5种灌木叶片结构的抗旱特征研究[J].西北林学院学报,2011,26 (4):13-17.
[24]蔡永立,宋永昌.浙江天童常绿阔叶林藤本植物的适应生态学:Ⅰ:叶片解剖特征的比较[J].植物生态学报,2001,25(1):90-98.
[25]ROCASG,BARROSC F,SCARANO FR.Leaf anatomy plasticity of Alchornea triplinervia(Euphorbiaceae)under distinct light regimes in a Brazilian montane Atlantic rain forest[J].Trees:Struct Funct,1997,11(8):469-473.
[26]ROCASG,SCARANO FR,BARROSCF.Leafanatomical variation in Alchornea triplinervia(Spreng)Müll.Arg.(Euphorbiaceae)under distinct light and soil water regimes[J].Bot JLinn Soc,2001,136(2):231-238.
[27]李春阳,TUOMELA K.桉树的抗旱性研究进展[J].世界林业研究,1998,1(3):23-28.
[28]张晓艳,杨惠敏,侯宗东,等.土壤水分和种植密度对春小麦叶片气孔的影响[J].植物生态学报,2003,27 (1):133-136.
[29]崔宏安,白红霞,丁虹茹,等.一球悬铃木叶结构的解剖研究[J].西北林学院学报,2008,23(6):66-68.
[30]崔秀萍,刘果厚,张瑞麟.浑善达克沙地不同生境下黄柳叶片解剖结构的比较[J].生态学报,2006,26 (6):1842-1847.
[31]寇建村,杨文权,贾志宽,等.不同紫花苜蓿品种叶片旱生结构的比较[J].西北农林科技大学学报:自然科学版,2008,36(8):67-72.
【责任编辑 李晓卉】
Drought resistance evaluation based on leaf anatom ical structures of 25 shrubs on the Tibetan Plateau
PAN Xin1,2,QIU Quan1,LIJiyue1,WANG Junhui3,HE Qian1,SU Yan1,MA Jianwei4,DU Kun4
(1 College of Forestry,South China Agricultural University,Guangzhou 510642,China; 2 Fuzhou Forestry Bureau,Fuzhou 350007,China;3 Research Institute of Forestry,Chinese Academy of Forestry/Key Laboratory of Tree Breeding and Cultivation,State Forestry Administration,Beijing 100091,China; 4 Xiaolongshan Forestry Science and Technology Research Institute,Tianshui741022,China)
【Objective】This study reported the leaf anatomical structure,stomatal characteristics,and the responsemechanism ofwater adaptability of25 shrubs on Tibetan Plateau,with an aim to provide a reference beneficial to afforestation and selection of drought-resistant plants on the Tibetan Plateau and an attempt to outline how to screen excellent drought-resistant plants.【Method】Conventional paraffin section technique was used tomake comparison with 12 indexes such as the thickness of leaf cuticle,upper epidermis,lower epidermis,palisade tissue,spongy tissue,and leaf thickness.The principal component analysis and subordinate function were used to develop a system to analyze and evaluate the plant drought resistance.【Resultand conclusion】The result showed that the leaves of all the 25 shrubs had typical xeromorphic structures,and the maximum leaf thickness was up to 323.00μm,186.25μm on average,and the maximum epidermal layer thickness was up to 4.83μm,with an average of 1.71μm,which presented that upper epidermiswas30.00%thicker than lower epidermis averagely,proving that the leafstomata were small and dense as the result of long-term adaptive evolution in a drought plateau environment.Moreover,12 indicator species differed from each other significantly.The five representative drought resistance indexes,including stomatal density,leaf thickness,palisade tissue thickness,spongy tissue thickness and stratum corneum thickness had been screened through a principal component analysis combined with the variation coefficient of each index.By using the method of subordinate function values,this study provides a sketch of content on sorting drought performance into different levels,including highly drought-resistant species,drought-tolerant species,drought-resistant in general species,and drought-sensitive species.
shrub;leaf anatomical structure;stomatal characteristic;drought resistance;drought stress; Tibetan Plateau
S718.43
A
1001-411X(2015)02-0061-08
潘 昕,邱 权,李吉跃,等.基于叶片解剖结构对青藏高原25种灌木的抗旱性评价[J].华南农业大学学报,2015,36(2):61-68.
2013-10-23 优先出版时间:2015-01-21
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20150121.0934.009.html
潘 昕(1989—),女,硕士研究生,E-mail:xiaowandoujiajia@126.com;通信作者:李吉跃(1959—),男,教授,博士,E-mail:ljyymy@vip.sina.com;王军辉(1972—),男,研究员,博士,E-mail:wangjh@caf.ac.cn
林业公益性行业科研专项(200904033);科学技术部农业科技成果转化资金项目(2011GB24320010)
