Inf l uence of topographic aspect on f l oristic diversity,structure and treeline of afromontane cloud forests in the Bale Mountains, Ethiopia
2015-06-09•••
•••
ORIGINAL PAPER
Inf l uence of topographic aspect on f l oristic diversity,structure and treeline of afromontane cloud forests in the Bale Mountains, Ethiopia
Eshetu Yirdaw1•Mike Starr1•Mesele Negash2•Fantaw Yimer2
©Northeast Forestry University and Springer-Verlag Berlin Heidelberg 2015
We studied the inf l uence of east and west aspects on f l oristic composition,diversity,structure and treeline of afromontane cloud forests at Rira in the Bale Mountains,southeast Ethiopia.In addition,we studied how aspect relates to and/or interacts with other topographic and edaphic factors in inf l uencing vegetation diversity.Stratifi ed systematic plot sampling was used to survey the fl oristic composition,diversity and structure of forests on east-and west-facing slopes.The sample plot size was 20×20 m and a total of 36 plots were inventoried.A total of 72 composite soil samples were collected and analysed. Woody species richness of the forest on the east-facing slope was 1.7 times higher than on the west-facing slope. Shannon,Simpson and Log-series alpha diversity indices and evenness of forests on the east-facing slope were signif i cantly higher than on the west-facing slope.NMDS ordination indicated that the east-and west-facing slopes formed two clusters of species and aspect explained 55.2% and 10.4%of the variation in species richness and abundance,respectively.There was no signif i cant difference between aspects in stand structure except in dominant height,which was higher on the east-facing slopes.There was signif i cant interaction between aspect and elevation in inf l uencing woody species diversity.The four plant community groups,which were identif i ed using cluster and indicator species analysis were represented differently on the east and west aspects.The treeline on the east-facing slope(3352 m)was located about 110 m higher than on the west-facing slope(3240 m).Soil moisture def i ciency was unlikely to be a limiting factor on either site.Near the equator,east–west aspect was shown to have considerable impact on f l oristic composition,diversity,structure,and treeline position of montane forests.Diurnal cloud movement patterns and its impact on microclimate of slope aspect should be taken into account in future studies of cloud forest diversity,structure,and treeline position.
Afromontane⋅Aspect⋅Cloud forests⋅Ethiopia⋅Floristic diversity
Introduction
Apart from elevation,topographic aspect exerts a major inf l uence on the structure and f l oristic composition of forests,and may generate differences in ecosystem characteristics,particularly in mountainous areas(Bale et al. 1998).By affecting the solar radiation budget,aspect inf l uences air and soil temperature,humidity,and soil moisture(Fekedulegn et al.2003;Sariyildiz et al.2005). The inf l uence of aspect on microclimatic condition affects woody species recruitment,plant assemblage,diversity, and growth(Holland and Steyn 1975;Brunori et al.1995; Bale et al.1998;Badano et al.2005).Moreover,by inf l uencing light regime and temperature,aspect has a strong role in controlling landscape-scale variations in thealpine treeline elevation(Danby 2003;Dalen and Hofgaard 2005).
In general,in the northern hemisphere south-and southwest-facing slopes receive more intense solar radiation and as a result tend to be drier than north-and northeast-facing slopes(Barnes et al.1998;Fekedulegn et al.2003).However,near the equator the variation in solar irradiance received by north-and south-facing slopes is less pronounced than at higher latitudes(Moore 2008). East-and west-facing slopes show similar but less extreme variation.East-facing slopes are exposed to direct sunlight in the cool of the morning and are normally cooler and moister than west-facing slopes(Barnes et al.1998;Yadav and Gupta 2006).Generally,slopes with afternoon sun (west-facing slopes)have higher maximum temperature than an equivalent slope with morning sun(McCune and Dylan 2002).In addition to aspect,slope steepness and diurnal cloud movement patterns also affect the magnitude of solar radiation received at any given site and this affects spatial patterns of plant association and diversity(Smith 1977;Bruce and Dylan 2002;Badano et al.2005).The fi ne-scale variation in aspect and slope gradient affects the local slope heterogeneity of mountainous landscapes that results in marked differences in biota and ecosystem function(Barnes et al.1998).
The Bale Mountains are one of largest massifs that form the south-eastern highlands of Ethiopia and are located approximately at 6°45′N,39°45′E(Yimer et al.2006a). This mountain block is formed of ancient volcanic rocks dissected by rivers and streams that have cut deep gorges. The Bale Mountains rise from the surrounding plateau at about 2500 m and reach 4377 m at the peak of Mt.Tullu Deemtu on the Sanetti plateau(Hillman 1988;Uhlig and Uhlig 1991).The Bale Mountains,lying southeast of the rift valley,are the largest area of Afro-alpine habitat in Africa with over 4000 km2above 3000 m.This is a high and extensive massif that comprises a wide range of habitats across vegetation belts,from montane forests to short Afro-alpine meadows and harbours many species of endemic f l ora and fauna.The Bale Mountains are included within the Eastern Afromontane biodiversity hotspot (Tallents and Macdonald 2011).The Bale Mountains National Park covers about 2200 km2,and includes the Harenna forest which is one of the largest remnant natural afromontane forests in Ethiopia(Hillman 1988;Tesfaye et al.2002).
In general,there is little scientif i c information available on the spatial ecology of montane cloud forests in Ethiopia, which are unique and threatened ecosystems.Therefore, the overall objective of this study was to investigate the inf l uence of east and west aspects,on the f l oristic composition,diversity,structure,and treeline of afromontane cloud forests at Rira,Bale Mountains,Ethiopia.The confounding effects of elevation,slope and edaphic factors were taken into account in the study.
Materials and methods
Study sites
The study was carried out at Rira in the Bale Mountains National Park about 400 km southeast of Addis Ababa (Fig.1).Two research sites were selected,one on eastfacing slope(6°46′N,39°41′E)and the other on west-facing slope(6°44′N,39°44′E).The research sites were characterized by gentle slopes to very steep mountain slopes with slope gradients ranging from 15 to 80%and elevations ranging from 3074 to 3274 m above sea level (a.s.l.).Because of the lack of long-term climatic data for the Rira area,the climate estimator software tool,New LocClim(FAO 2005;Grieser et al.2006)was used to generate long-term monthly rainfall and temperature data for each sample plot.Mean annual temperature(MAT)and mean annual precipitation(MAP)at the east and west aspect were estimated to be 14.3 and 14.6°C and 894 and 905 mm/year,respectively.Rainfall at Rira has a bimodal pattern with a long rainy season lasting from June to October and a short rainy season from March to May (Fig.2).Cloud or mist is quite common at the study sites because of the prevalence of moist south-western air currents,which are intercepted by the Harenna escarpment (Hillman 1988;Nigatu and Tadesse 1989).The soils are of volcanic origin welded with volcanic ash materials, weathered to mainly black to very dark brown,silt loams rich in organic matter,loam-loamy sand and silt clay loam (Yimer et al.2006a,2006b).Andosols are the most prevalent soil types found at higher elevations of the Bale Mountains(Yimer 2006b;Amare et al.2013).
The forests at Rira are cloud forests located at high elevation on the northern fringes ofthe Harenna forest,which is classif i ed as moist evergreen afromontane forest(Friis et al. 2010).The Harenna forest is situated on the southern slopes of the Bale Mountains at elevations of 1300–3500 m asl (Bussmann 1997;Senbeta and Denich 2006;Friis et al. 2010).Floristically,the Harenna forest is closely related to the south-western Ethiopian afromontane rainforest and is thought to be one of the major centers of endemism containing diverse gene reservesincluding wild coffee(Hillman 1988;Yimer et al.2006a,2006b;Friis et al.2010).The vegetation formations in the Harenna forest are mainly of the afromontane and afroalpine types,showing marked elevational vegetation zonation.The Harenna forest is stratif i ed by vegetation belts,such asErica-Hypericum-Hagenia,Arundinaria alpinaK.Schum.,Scheff l era-Hagenia,Aningeria,Podocarpusfrom highest to lowestelevations,respectively(Hillman 1988;Yimer et al.2006a, 2006b).At the upper elevation limit,the Harenna forest gives way to a sub-Afroalpine vegetation type,characterized byEricabushland(Nigatu and Tadesse 1989).
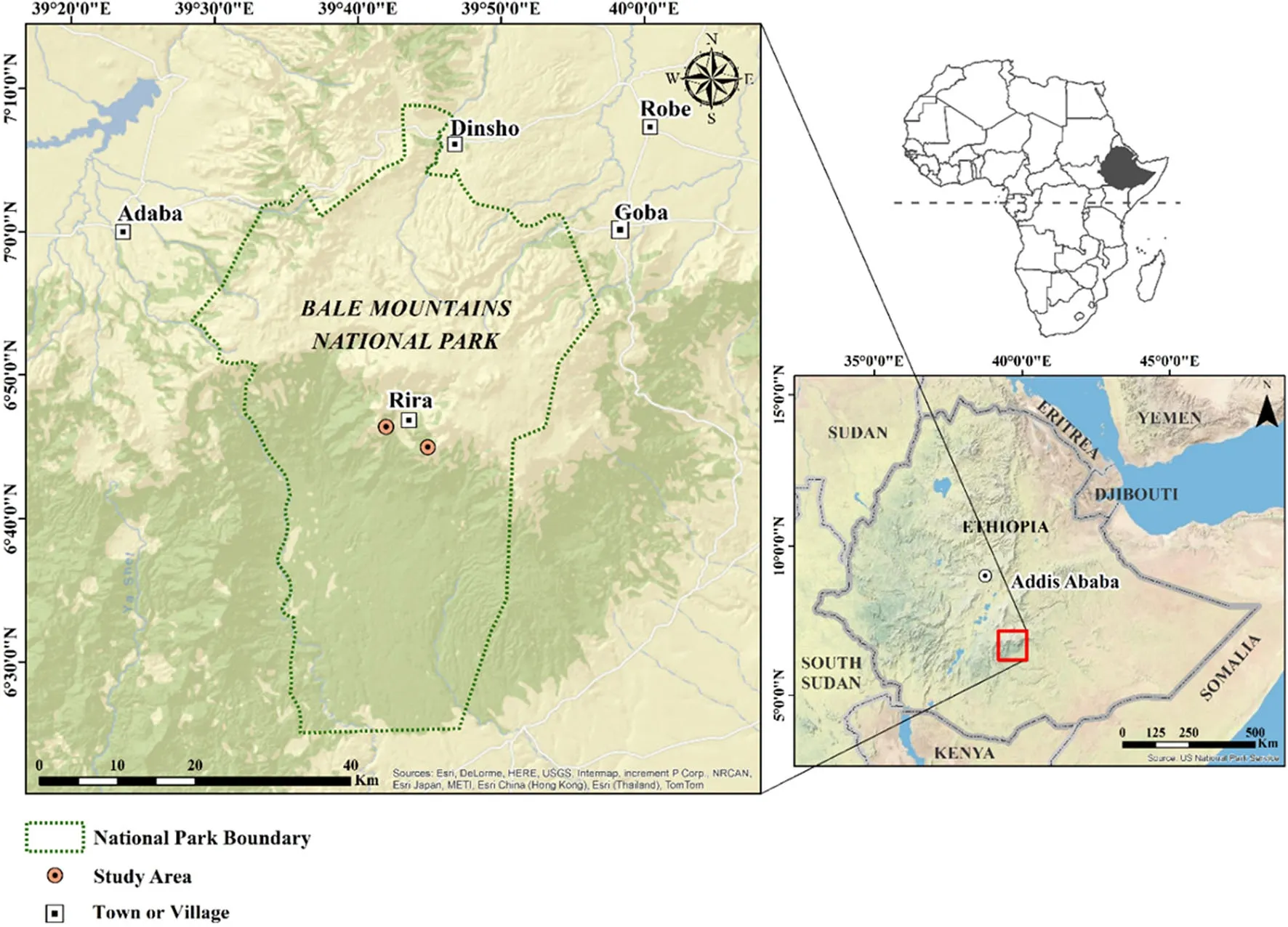
Fig.1 Maps of Africa and Ethiopia and the location of study sites at Rira,Bale Mountains,Ethiopia

Fig.2 Long-term mean monthly temperature(T),rainfall(P),and potential evapotranspiration(PET)at the study sites(values generated using New LocClim.See text)
Human disturbances and encroachment into the Harenna forest for crop cultivation,animal grazing,selective logging and settlement expansion has led to forest degradation and deforestation(Tesfaye et al.2004;Senbeta and Denich 2006;Yimer et al.2007).Furthermore,at higher elevations (above~3300 m)f i re is used by montane pastoralists to burn theEricascrub vegetation to promote the regeneration of grasses,herbs,and youngEricashoots,which are grazed or browsed by livestock(Johansson et al.2012).In general,the increasing population within and in the vicinity of the Harenna/Rira forest has been the main driver of increased extraction of wood and non-wood forest products (Hillman 1988;Tesfaye et al.2002;Teshome et al.2011).
Inventory design
Woody species
Stratif i ed systematic sampling of 36 plots,each measuring 20×20 m(18 plots per aspect),was used to survey the fl oristic composition,diversity and structureofthe forestson east-and west-facing slopes.For each aspect,six plots were placed at 100 m intervals along three horizontal line transects separated by an elevation interval of 100 m(3074 m a.s.l.lower;3174 m asl,middle;and 3274 m asl,higher). The totalsampled area was1.44 ha,which constituted 5.3% of the total study area.The initial plots at all the elevation levels were selected randomly.The elevation,slope,aspect and GPS location of each plot were determined.
In each plot,the number of tree and shrub species(including woody lianas),number of individuals of each species,dominant height,and basal area were measured.Dead trees and shrubs were also counted.In addition,the diameter at breast height(DBH)of all trees and shrubs(≥1.3 m height)were measured.Woody plants were grouped by size classes as:germinants(height≤0.1 m),seedlings (0.1<height<1.3 m),and saplings(height≥1.3 m and DBH<2.5 cm).The different strata of the forest(lower, middle,upper and emergent)and their average heights were determined for each of the six horizontal transect lines.
Herbaceous layer
Five 1-m2quadrats were randomly placed in each sample plot for estimating the average ground cover percentage of the herbaceous layer.The average of the f i ve values was used to estimate the ground cover percentage.The dominant species of the herbaceous layer were identif i ed.
Canopy closure and leaf area index(LAI)
Hemispherical photographs were used to measure the canopy closure and LAI of the forest.Hemispherical photographs were taken with a Canon 7.5 mm(objective)f i sheye lens camera and Kodak 200 ASA black-and-white f i lm. Pictures were taken from a height of 1 m from the ground when the sky was overcast,early in the morning or late in the afternoon.The negatives of the hemispherical photographs were scanned using a f l at-bed scanner.The digitised pictures were then analysed using the Gap Light Analyzer(Version 2)hemispherical photograph analysis computer programme(Frazer et al.1999).
Species identif i cation
Most of the plant species were identif i ed in the f i eld using botanical keys and local knowledge.The few species that could not be identif i ed in the f i eld were identif i ed(except one species)using various botanical keys and books (Hedberg and Edwards 1989;Edwards et al.1997;Hedberg et al.2003).
Treeline
Eight points were randomly selected directly upwards from the transect line at elevation of 3274 m to determine the elevation(using altimeter and GPS)of the treeline on both the east and west-facing slopes.The eight points were located within a horizontal distance of 500 m in length.In this study,the treeline was def i ned as the location,whereErica arboreatrees more or less abruptly changed to shrub size,i.e.<1.5 m in height.
Soil sampling
Soil samples were taken from the four corners and center of each 20×20 m sample plot with a gouge soil auger(8 cm diameter).Samples of the 0–20 and 20–40 cm layers were taken for analysis.The 0–40 cm layer represents the top soil where most of the plant roots and nutrients are concentrated. The samples from each plot were composited by layer resulting in a total of 72 composite samples(36 from each slope aspect).In addition,undisturbed soil cores for the determination of bulk density were taken with steel cylinders of5 cm diameterand height(98.2 cm3).Priorto soilphysical and chemical analyses,the auger samples were air-dried at room temperature and passed through a 2 mm soil sieve,the<2 mm soilbeing retained foranalysis.Particle size fractions of the<2 mm soil were determined by hydrometer after dispersion in a mixerwith hexametaphosphate.Bulk density was calculated from the volumetric soil core samples after drying at 105°C for 24 h.Soil pH was measured with a combined electrode in a 1:2.5 soil to water suspension.Soil organic carbon was determined according to the Walkley and Black method(Schnitzer 1982).Available water capacity(AWC) was calculated as the difference between the amountof water (mm)held at f i eld capacity(FC)and permanent wilting point (PWP)computed using the pedotransfer function developed by Ritchieetal.(1999).Particlesize fraction,bulk density and organic carbon content were used as input data.
Data analysis
In addition to species richness and abundance,Shannon, Simpson and Log-series alpha diversity indices were used to measure woody species diversity on the east-and westfacing slopes.Shannon evenness was used to separately calculate the species evenness.Sørensen’s quantitative index was used to measure the similarity in woody species composition of the forests on east-and west-facing slopes. The R version 3.0.2 and PC-ORD version 6.15 software packages were used to calculate diversity and similarity indices(R Core Team 2013;McCune and Mefford 2011).
Two-way ANOVA(analysis of variance)was used to detect differences in species richness,abundance,Shannon, Simpson,Log-series alpha,indices and evenness in relation to aspect,elevation and their interaction.Signif i cantly different means were compared by Tukey multiple comparison tests.Similarly,one-way ANOVA and Tukey’s HSD test were used to detect variations in stand structure attributes.
A non-metric multidimensional scaling(NMDS)ordination based on Sørensen’s quantitative similarity index(as a distance measure)was used to investigate the similarity/ dissimilarity of the forests on east-and west-facing slopes in terms of woody species composition.NMDS was also used to study the inf l uence of environmental variables,such as elevation,slope and soil parameters on species distribution.The NMDS was carried out using PC-ORD for Windows software(version 6.15).A random starting confi guration with stability criterion of 10-5was used and 200 iterations were conducted,which resulted in a two-dimensional solution.A Monte Carlo test was conducted to evaluate whether NMDS extracted stronger axes than expected by chance(McCune and Grace 2002).For the Monte Carlo test,50 randomized runs were carried out and the probability that a similar f i nal stress could have been obtained by chance was very low(P<0.05).Two-tailed Pearson correlation was conducted to determine the type and strength of relationship between the ordination axes and species richness,abundance,aspect,slope,elevation and the soil parameters.
Hierarchical cluster analysis was carried out using PCORD for Windows software in order to def i ne plant community groups.In the cluster analysis Ward’s method was used as a linkage method,while relative Euclidean was used as a distance measure.
Multi-response permutation procedures(MRPP)was used to test differences in species composition between the plant community groups and to test within-group homogeneity.The Sørensen(Bray-Curtis)distance measure was used in the MRPP analysis.
An indicator species analysis was conducted to identify the indicatorspeciesforthe plantcommunity groupson eastand west-facing slopes.In this study,plant community groups were named by the two species which had the highest indicator values(IV).In the indicator species analysis,the Monte Carlo test of signif i cance of observed maximum IV for each species was based on 4999 randomizations.
Rare species were determined for both the east-and west-facing slopes.In this study,rare species are def i ned as species that fell in the lower quartile of the species distribution(Gaston 1994).
Soil data were grouped and summarized according to the elevation zones,aspects and soil depths.Statistical differences were tested using three-way ANOVA following the general linear model(GLM)procedure of SPSS Version 12.0.1 for Windows(Pallant 2001).Tukey’s honest signif i cance difference(HSD)test was used for post hoc tests of group differences(P<0.05).
Results
Floristic composition
Woody species
Sixteen tree and shrub species belonging to 14 families were recorded in the 36 sample plots.The three most abundant species on both east and west aspects in decreasing order wereErica arborea(tree size),Rappanea simensis(Hochst.ex A.DC.)Mez,andHypericum revolutumVahl,accounting for 54,25,10%and 40,17,11% of the total tree population,respectively.On the east-facing slopes,R.simensiswas the most frequent(94%of the plots),whileMaytenus ovatus(Wall.ex W.&A.)Loes was the least frequent species.On the west-facing slopes,Erica arborea,R.simensisandH.revolutum(56%each) were the most frequent,whileHagenia abyssinica(7%) was the least frequent species.Scheff l era abyssinica(A. Rich.)Harms was the second most frequent tree species on both east and west aspects and it was present on 83 and 50%of all plots,respectively.The rare woody species found on both east and west aspects were:Cassipourea malosana(Bak.)Alston,DiscopodiumpenninerviumHochst.,Arundinaria alpina,andPittosporum viridif l orumSims.
Shrub-sizeErica arboreaandPittosporum viridif l orumwere found only on west-facing slopes,whereasMaytenus ovatuswas found only on east-facing slopes.Clumps of mountain bamboo(A.alpina)were found also only on east aspects at elevations up to 3174 m.H.abyssinicawas present at all elevations on east aspects,whereas it was only found at lower elevations on west aspects.S.abyssinicawas found at all elevations on both aspects except at the higher elevation on west-facing slopes.
On east aspects,the upper canopy at elevation of 3074 m was dominated byH.abyssinica,at 3174 m it was dominated byR.simensisandS.abyssinica,while at 3274 m it was dominated by tree-sizeE.arborea.On west aspects,the upper canopy at 3074 m was dominated byR. simensis,S.abyssinicaandH.abyssinica,at 3174 m itwasdominated byR.simensis,S.abyssinicaand tree-sizeE. arborea,while at 3274 m it was dominated by shrub-sizeE.arborea.
Climbers
A total of six climber species were recorded in the forests on both east-and west-facing slopes.The species were:Ipomea kituiensisVatke,Stephania abyssinica(Dillon and A.Rich.)Walp.,Clematis brachiata(Thunb.),Zehneria scabra(L.f.)Sond.,Mikaniopsis clematoides(Sch.Bip.ex A.Rich.)Milne-Redh,and Miesa(local name).
Herbacous species
There was no signif i cant difference in the estimated ground herbaceous cover percentage between east and west aspects.The east and west aspects had average herbaceous ground cover of 94 and 95%,respectively.The main herbaceous species were:Mimulopsis solmsiiSchweinf.,AlchemillaabyssinicaFresen.,VeronicaglandulosaBenth.,Sedum cf churchillianumRobyns and Boutique, andImpatiensspp.
Epiphytic f l ora
The tree trunks and main branches at the two study sites were covered with a thick mat of mosses and lichens(up to 20 cm in thickness).Some epiphytic ferns and woody species(Ex.Hagenia abyssinica)were anchored and growing on the thick layer of mosses and lichens.
Stand structure
The stand characteristics of the afromantane forest for each aspect and elevation zone are described in Table 1.Except for dominant height,there was no signif i cant(P>0.05) difference between aspects in terms of stand structure,but stand structure varied by elevation on both aspects.Tree DBH on the east aspect and both dominant height and basal area on the west aspect showed a declining trend from lower to higher elevation.The highest basal area was recorded(47 m2/ha)on the east aspect,and the lowest (17 m2/ha)was on the west aspect at higher elevations.On the west aspect,basal area was higher by 16%at lower elevation,but lower by 7 and 64%for middle and higher elevations,respectively,than on the east aspect.This implies that the west-facing slope had lower DBH of trees at middle and higher elevations.Stem number varied signif i cantly(P<0.001)by elevation.Stem number increased substantially from lower(172 stems/ha)to higher elevation(1388 stems/ha)on the east aspect,but showed an inverted parabolic trend on the west aspect.Stem number on the east aspect was higher by 56%at higher elevations but lower by 25 and 62%at lower and middle elevations,respectively,than on the west aspect.Canopy closure and leaf area index(LAI)did not differ signif icantly between the two aspects,but differed by elevation. The tree canopy cover was higher on west aspects (67–79%)than on east aspects(50–64%)except at higher elevations.The highest canopy tree cover and LAI were recorded on east aspect at higher elevation(92.5%and 3.28,respectively),while the lowest was at lower elevation (50%and 0.58,respectively)on east-facing slopes.
On the eastern aspect,R.simensiswas the most abundant species at lower and middle elevations(280 and 633 stems/ ha,respectively),whereas tree-sizeE.arboreawas the most abundant at these elevations(600 and 1717 stems/ha, respectively)on the western aspect.Athigherelevation,treesizeE.arboreaaccounted for the highest stem density (4050 stems/ha,70%of the total tree population)on the eastern aspect,whereas shrub-sizeE.arboreaandH.revolutumwere the only woody vegetation on the western aspect at this elevation(2167 stems/ha).Shrub-sizeE.arboreahad a multi-stemmed growth form and occurred as a clump.Its mean height and DBH were 1.46 m and 109 cm,respectively.On the average,the number of dead trees and shrubs was nearly f i ve(4.8)per sample plot.
Forest stands diversity and environmental factors
Woody species diversity
Woody species richness of the forest on the east-facing slope was 1.74 times higher than that on the west-facing slope(Table 2).Shannon,Simpson and Log-series alpha diversity indices and evenness values were also signif icantly higher on the east-facing than on the west-facing slope.Species richness,diversity indices and evenness on both east-and west-facing slopes were highest at the middle elevation followed by the lower and higher elevations.Except for abundance,all diversity indices andevenness had the lowest values at the highest elevation. The difference in woody species diversity between the lower and middle elevations was not signif i cant,while the difference between the higher elevation and the other elevation zones was signif i cant.In general,species richness on both aspects declined sharply when approaching the Ericaceous belt.Contrary to species richness,abundance was highest at higher elevation,mainly due to numerous and denseEricabushes.A 2-way ANOVA showed that there was a signif i cant interaction effect between aspect and elevation on woody species diversity(Table 3).Floristic similarity of the sample plots was mainly determined by elevation and aspect(Table 4).

Table 1 Mean DBH,dominant height(DOH),basal area(BA),stem numbers,canopy closure and leaf area index(LAI)for each elevation zone and aspect of the afromontane cloud forests at Rira,Ethiopia

Table 2 The mean woody species richness,abundance and biodiversity indices of east-and west-facing slopes at Rira,Ethiopia

Table 3 Summary of two-way ANOVA results for species richness,abundance,Shannon,Simpson,Log alpha and Evenness in relation to topographic aspect(AS)and elevation zones(EZ)
Ordination
NMDS ordination yielded a two-dimensional solution,with a reduced stress value of 11.1(Fig.3).NMDS axes 1 and 2 explained 22.5 and 62.3%of the total variation,respectively(cumulative R2=0.848).Compared to axis 1,axis 2 had a stronger and signif i cant correlation with abundance (r=0.388,P<0.05 with axis 1 andr=0.824,P<0.01 with axis 2)and species richness(r=-0.033,NS with axis 1 andr=-0.701,P<0.01 with axis 2).
Aspect was a signif i cantly correlated with both axis 1 (r=-0.340,P<0.05)and axis 2(r=0.507,P<0.01). The strongest correlation was,however,shown by elevation;axis 1(r=0.578,P<0.01)and axis 2(r=0.740,P<0.01).Plot slope-angle and soil clay content were not signif i cantly correlated with axis 1,but they were signif icantly correlated with axis 2.
Aspect explained 55.2 and 10.4%of the variation in species richness and abundance,respectively,whereas, elevation explained 7.4 and 48.2%of the variation in species richness and abundance.Species richness was negatively correlated with elevation.
Plant communities
Based on cluster and indicator species analysis,four communities of woody species were identif i ed on the eastand west-facing slopes(Table 5).After dividing the plant communities into four groups in the cluster analysis about 66%of the information was retained.
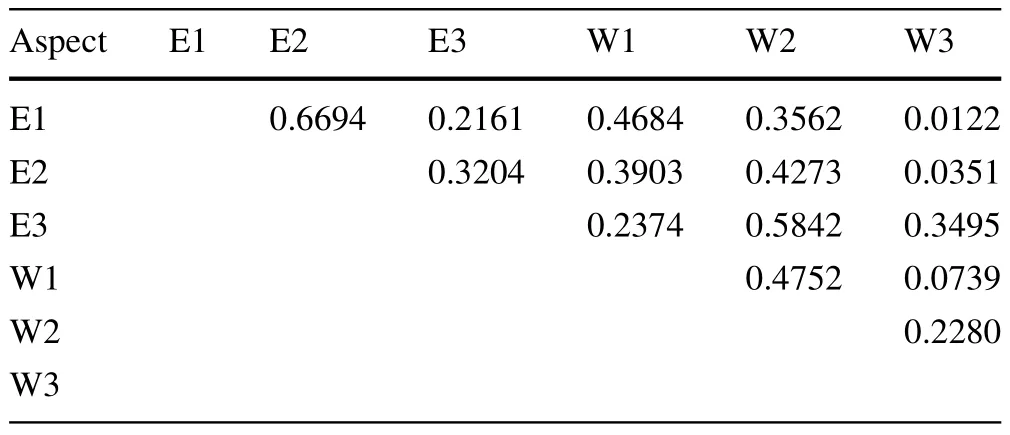
Table 4 Sørensen quantitative similarity index for the east-and west-facing slopes at Rira,Ethiopia
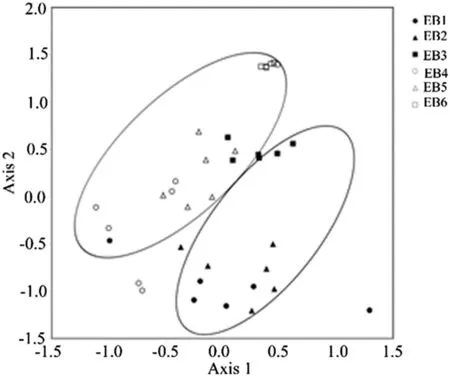
Fig.3 Non-metric multidimensional scaling(NMDS)ordination of 36 plots representing,two aspects and three elevation classes at Rira, Ethiopia.EB1 east lower,EB2 east middle,EB3 east higher,WB1 west lower,WB2 west middle,WB3 west higher
MRPP showed signi fi cant differences in woody species composition between the four communities(P<0.001). The test statistic T and the agreement statistic A had values of-17.1 and 0.38,respectively.The test statistictdescribes the separation between groups(the more negativetvalues,the stronger the separation),while A describes within-group homogeneity.Six combinations of pair-wiset-test comparisons were conducted and showed signi fi cant difference between the four communities(P<0.001).The separation between communities one and two was the highest(t=-12.3),while the separation between one and three(t=-5.8)was the lowest.
Treeline
On the east-facing slope(3352 m)the treeline was located about 110 m higher in elevation than on the west-facing slope(3240 m).By far,the dominant woody species at treeline was the shrub-sizeErica arborea(about 90%of the stems)followed byHypericum revolutum(10%).These are characteristic species of the afromontane Ericaceous belt.
Soil
Plot mean soil properties by aspect,elevation and soil depth are presented in Table 6.Soil texture on both east and west aspects varied from sandy loam to loam.Clay content increased from east to west aspect and soil depth, but decreased with elevation.Clay content differed signif i cantly by aspect,soil depth and elevation(Table 7).As is typical of montane cloud forests,the forest soil at Rira had a high organic carbon content and low bulk density (Hamilton 1995;Yimer et al.2007).Soil organic carbon was signif i cantly higher on the west-facing slope and in the surface layer.Soil bulk density for surface and sub-horizon layers differed signif i cantly(P<0.001)by aspect and elevation but not by soil depth.Soil acidity decreased from east to west aspect and increased with soil depth and elevation.The difference in soil acidity between aspects was signif i cant(P<0.001),but did not signif i cantly differ by elevation(Table 7).
Discussion and conclusions
Comparison of Rira with lower elevation Afromontane forests
Based on the def i nition by Scatena et al.(2010),the lower and higher elevation forests at Rira may be classif i ed as upper montane and sub-Afroalpine cloud forests,respectively.In general,because of the high elevation and nearly ever-present cloud cover of the study sites at Rira,woody species diversity is considerably poorer than at afromontane forests located at lower elevations.For instance,the relatively low-lying part of the Harenna forest (1550–1650 m)was reported to contain on average about 61 woody species(Tadesse and Nigatu 1996),whereas only 16 woody species were recorded at Rira.Similarly, other afromontane forests in Ethiopia located at elevations ranging from 1500 to 3000 m,such as the Jibat,Menagesha,Wofwasha,Wondo Genet,and Mana Angetu forests were reported to contain on average about 52,30,51,72, and 117 woody species,respectively(Bekele 1994;Teketay and Bekele 1995;Lulekal et al.2008;Kebede et al. 2013).Studies conducted at Mt.Elgon and Mau forest in Kenya found 49 and 50 tree species at elevations of 1200–2400 m and 2100–2700 m,respectively(Hitimana et al.2004;Kinjanjui et al.2013).In general,this f i nding is in agreement with the widely accepted general pattern thatspecies richness declines with increasing elevation(Uhlig 1988;Rahbek 1995).Woody species composition at Rira was also noticeably different from lower elevation afromontane forests in that typical sub-Afroalpine species such asEricaspp.andHypericum revolutumwere present. Unlike other afromontane forests,epiphytic f l ora(covering tree trunks,branches and fallen trees and twigs)was abundant at Rira,which is one of the outstanding features of the tropical montane cloud forests.
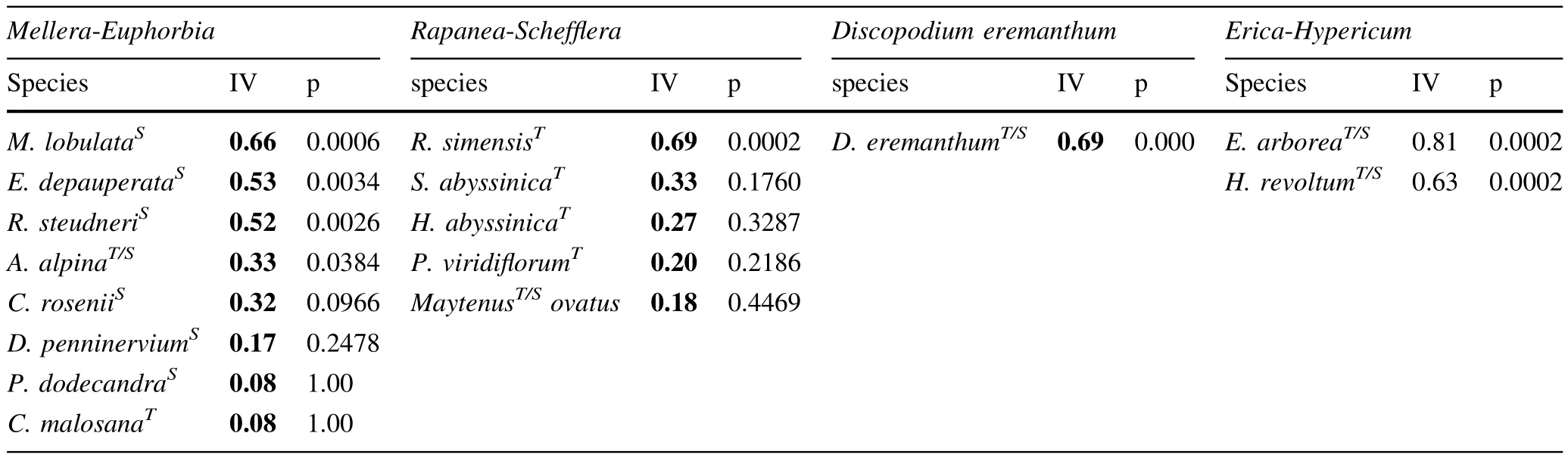
Table 5 Woody species communities on the east-and west-facing slopes

Table 6 Mean(±SD)soil properties for the six plots representing each elevation zone and aspect of the afromontane cloud forests at Rira, Ethiopia
Tree stature(height,DBH,basal area)at Rira tended to be considerably smaller and contains fewer canopy strata than afromontane forests at lower elevations(Bekele 1994; Senbeta and Denich 2006).The lower tree stature and the prevalence of gnarled stems and branches at Rira are in line with the physiognomic characteristics exhibited by upper montane cloud forests(Foster 2001;Scatena et al.2010). Generally,there is a strong thermal advantage of being short,which is why dwarf shrubs succeed at high elevations whilst trees do not(Grace et al.2002).The lower stem number and canopy closure percentage at lower elevation(Table 1)may be a result of cuttings by local people.Particularly,harvesting of bamboo(mainly from lower elevations)for construction purposes is common in the area.
Inf l uence of aspect on diversity,composition, structure and treeline
The forest on the east-facing slope had signif i cantly higher woody species richness,evenness and Shannon,Simpsonand Log-series alpha diversity values than on the westfacing slope(Table 2).The vegetation on the west aspect seemed similar to the vegetation on the east aspect at about 100 m higher in elevation.This was manifested by the higher similarity of the vegetation at E3 and E2 to W2 and W1,respectively;the west aspect plots being 100 m lower in elevation(Table 4).Similarly,the dominant canopy species on the west aspect resembled those on the east aspect at elevations 100 m higher.

?
There was also a difference in the representation (abundance)of plant community groups found on the east and west-facing slopes.For instance,on the west aspect, theMellera-Euphorbiagroup was scarcely represented, whereas theErica-Hypericumgroup was abundant in comparison to the east-facing slope.In addition,shrub-sizeErica arboreawas found only on the west-facing slopes indicating that the Ericaceous belt is located at lower elevation on the west compared to the east aspect.The fact that the treeline on the east aspect was located about 110 m higher than on the west aspect also indicates that there is distinct elevational difference in species assemblage similarity between the two contrasting aspects.In general, second to elevation,aspect was the main physiographic factor that impacted woody species diversity and composition.
In addition to their separate inf l uences,the combined effects of aspect and elevation on woody species diversity was signif i cant.Consequently,the inf l uence of aspect on woody species diversity was more pronounced at higher elevation.
In general,aspect had a more pronounced impact on woody species diversity than on forest structure(Tables 1, 2).However,dominant height was signif i cantly greater on the east-than on the west-facing slope.The lower stature of trees on the west aspect indicates that the microclimatic conditions on the west aspect have characteristics of relatively higher elevation compared to the east aspect.
The higher woody species diversity,tree stature and treeline elevation was most probably due to the higher amount of solar radiation received and consequently the increased air and soil temperature on the east-facing slopes.The east-facing slopes received morning sunshine, while during mid-day and afternoon there was usually a formation of cloud cover,which reduced the solar radiation received by the west-facing slopes.Due to convective uplift of moist air,the afternoons on Bale Mountains tend to have orographic cloud cover and rain,which reduces incident solar radiation that reaches the forest canopy and the ground(Wesche 2003).Hence,microclimate models should take into account the diurnal cloud movement patterns and cloud type,which are important factors in determining the incident radiation and temperature at the canopy and forest f l oor.
The lower temperature(as a consequence of lower solar radiation)on west-facing slopes may be detrimental for some woody species to establish and thrive at such a high elevation and this may result in the lowering of the woody species diversity.Moreover,lower amount of solar radiation and temperature are the most probable factors for the location of the treeline at lower elevation on the westfacing slopes.Increasing precipitation and cloudiness reduce solar radiation and temperature and consequently tend to lower treeline elevation(Grubb 1997;Ko¨rner 1998; Grace et al.2002).However,the human forest disturbance particularly by f i re,which is prevalent in the Afroalpine and sub-Afroalpine zones of the Bale Mountains,may also lower the treeline(Miehe and Miehe 1994;Wesche et al. 2000;Bader and Ruijten 2008;Johansson et al.2012).
The higher organic carbon content of the soil on the west-facing slope compared to the east-facing slope (Table 6)may also indicate among others a lower temperature and as a result lower litter decomposition rate(cf. Zhang et al.2008;Negash and Starr 2013).
As there was no substantial difference in the amount of rainfall,available water capacity and soil texture(Fig.2 and Table 7),it is unlikely that the difference in available soil moisture explains the signif i cant difference in woody species diversity and tree stature between the east and west aspects.In addition to rainfall,the forests at Rira receive precipitation by canopy interception of cloud water or cloud stripping(the Rira rainfall data do not take cloud stripping into account).The amount of precipitation obtained from cloud stripping can be substantial;typically 5–20%of annual rainfall,although values exceeding 100%have been recorded(Foster 2001).Moreover,the high prevalence of cloud cover,canopy wetting,and lower temperature reduces substantially the evapotranspiration from the forests on the east and west aspects and consequently lowers the likelihood of soil moisture def i cit acting as a limiting factor.
The result of the present study is in agreement with the fi ndings of Smith(1977),and in contrast to the f i ndings of Bader and Ruijten(2008),Fekedulegn et al.(2004),and Yadav and Gupta(2006).One of the factors causing the contrasting results obtained by different authors may be the variation in the prevalence of cloud cover at the respective study sites.In a study on the effects of topographic aspect on montane cloud forest diversity,composition and structure,diurnal cloud movement patterns and the associated microclimatic conditions should be taken into account. Furthermore,species distribution models should take diurnal cloud movement patterns and the associated microclimatic conditions as inf l uencing factors.
At or near the equator,north-and south-facing slopes receive by and large similar amounts of solar radiation,but it would occur at different times depending on the aspect (Moore 2008;Gallardo-Cruz et al.2009).However,as shown in this study the east and west aspects,which receive morning and afternoon sunlight,can have signif icant differences in f l oristic composition,diversity and treeline location.Unlike mid latitudes,near the equator the impact of east versus west aspects can be more pronounced than the impacts of north versus south aspects in terms of fl oristic diversity and treeline position.
AcknowledgmentsThe authors are very grateful for the logistical support offered by the Wondo Genet College of Forestry and Natural Resources during the f i eld work.The f i nancial support of the Department of Forest Sciences,University of Helsinki is thankfully acknowledged.
Amare T,Hergarten C,Hurni H,Wolfgramm B,Yitaferu B,Gebre Selassie Y(2013)Prediction of soil organic carbon for ethiopian highlands using soil spectroscopy.ISRN Soil Sci 2013:1–11. http://dx.doi.org/10.1155/2013/720589.http://www.hindawi. com/journals/isrn/2013/720589/Accessed 14 Jan 2014
Badano EI,Cavieres LA,Molina-Montenegro MA,Quiroz CL(2005) Slope aspect inf l uences plant association patterns in the Mediterranean matorral of central Chile.J Arid Environ 62:93–108
Bader MY,Ruijten JA(2008)A topography-based model of forest cover at the alpine tree line in the tropical Andes.J Biogeogr 35:711–723
Bale CL,Williams JB,Charley JL(1998)The impact of aspect on forest structure and f l oristics in some Eastern Australian sites. For Ecol Manag 110:363–377
Barnes BV,Zak DR,Denton SR,Spur SH(1998)Forest ecology,4th edn.Wiley,New York,p 792
Bekele T(1994)Vegetation ecology ofremnantAfromontane forestsofthe central plateau of Shewa,Ethiopia.Acta Phytogeogr Suec 79:1–59
Brunori A,Nair PKR,Rockwood DL(1995)Performance of two Eucalyptus species at different slope positions and aspects in a contour-ridge planting system in the Negev desert of Israel.For Ecol Manag 75:41–48
Bussmann R(1997)The forest vegetation of the Harenna escarpment (Bale province,Ethiopia)—syntaxonomy and phytogeographical aff i nities.Phytocoenologia 27(1):1–23
Dalen L,Hofgaard A(2005)Diffrentail regional tree dynamics in the Scandes mountains.Antarct Alp Res 37(3):284–296
Danby R(2003)A multiscale study of tree-line dynamics in southwestern yukon.Arctic 56(4):427–436
Edwards S,Demissew S,Hedberg I(1997)Flora of Ethiopia and Eritrea:Hydrocharitaceae to Arecaceae,vol 6.The National Herbarium,Addis Ababa University,Addis Ababa,p 586
FAO(2005)New_LocClim:local climate estimator(version 1.10). Environment and natural resources,working paper number 20 (CD-ROM).FAO,Rome
Fekedulegn D,Hicks RR,Colbert JJ(2003)Inf l uence of topographic aspect,precipitation and drought on radial growth of four major tree species in an Appalachian watershed.For Ecol Manag 177:409–425
Fekedulegn D,Colbert JJ,Rentch JS,Gottschalk KW(2004)Aspect differences in vegetation,soil and microclimatic characteristics of an Appalachian watershed.Castanea 69(2):92–108
Foster P(2001)The potential negative impacts of global climate change on tropical montane cloud forests.Earth-Sci Rev 55:73–106
Frazer GW,Canham CD,Lertzman KP(1999)Gap light analyzer (GLA),version 2.0:imaging software to extract canopy structure and gap light transmission indices from true-colour f i sheye photographs,users manual and program documentation.Millbrook,Simon Fraser University,Burnaby,British Columbia,and the Institute of Ecosystem Studies
Friis I,Demissew S,Breugel P(2010)Atlas of the potential vegetation of Ethiopia.Det Kongelige Danske Videnskabernes Selskab,Copenhagen,p 306
Gallardo-Cruz JA,Perez-Garcia EA,Meave JA(2009)β-diversity and vegetation structure as inf l uenced by slope aspect and altitude in a seasonally dry tropical landscape.Landscape Ecol 24:473–482
Gaston KJ(1994)Rarity.Population and community biology series 13.Chapman and Hall,London,p 205
Grace J,Berninger F,Nagy L(2002)Impacts of climate change on tree line.Ann Bot 90:537–544
Grieser J,Gommes R,Bernardi M(2006)New LocClim—the local climate estimator of FAO.Geophys Res Abstr 8:2
Grubb PJ(1997)Control of forest growth and distribution on wet tropical mountains:with special reference to mineral nutrition. Annu Rev Ecol Syst 8:83–107
Hamilton LS(1995)Mountain cloud forest conservation and research: a synopsis.Mt Res Dev 15(3):259–266
Hedberg I,Edwards S(1989)Flora of Ethiopia,vol 3.Pittosporaceae to Araliaceae.The National Herbarium,Addis Ababa and Uppsala University,Uppsala,p 659
Hedberg I,Edwards S,Nemomissa S(2003)Flora of Ethiopia and Eritrea,vol 4(2).Part 1:Apiaceae to Dipsacaceae.The National Herbarium,Addis Ababa and Uppsala University,Uppsala
Hillman JC(1988)The Bale mountain national park area,southeast Ethiopia,and its management.Mt Res Dev 8(2/3):253–258
Hitimana J,KiyiapiJL,Njunge JT(2004)Foreststructure characteristics in disturbed and undisturbed sitesofMt.Elgon moistlowermontane forest,western Kenya.For Ecol Manag 194:269–291
Holland PG,Steyn DG(1975)Vegetational responses to latitudinal variations in slope angle and aspect.J Biogeogr 2:179–183
Johansson MU,Fetene M,Malmer A,Granstro¨m A(2012)Tending for cattle:traditional f i re managment in Ethiopian montane heathlands.Ecol Soc 17(3):19
Kebede M,Kanninen M,Yirdaw E,Lemenih M(2013)Vegetation structural characteristics and topographic factors in the remnant moist Afromontane forest of Wondo Genet,south central Ethiopia.J For Res 24(3):419–430
Kinjanjui JM,Karachi M,Ondimu KN(2013)Natural regeneration and ecological recovery in Mau forest complex.Kenya.Open J Ecol 3(6):417–422
Ko¨rner C(1998)A re-assessment of high elevation tree-line positions and their explanation.Oecologia 115:445–459
Lulekal E,Kelbessa E,Bekele T,Yineger H(2008)Plant species composition and structure of the Mana Angetu moist montane forest,south-eastern Ethiopia.J East Afr Nat Hist 97(2):165–185
McCune B,Dylan K(2002)Equations for potential annual direct incident radiation and heat load.J Veg Sci 13:603–606
McCune B,Grace BJ(2002)Analysis of ecological communities. MJM Softw Des,Oregon,p 300
McCune B,Mefford MJ(2011)PC-ORD.Multivariate analysis of ecological data,version 6.MjM Software,Gleneden Beach
Miehe G,Miehe S(1994)Zur oberen Waldgrenze in tropischen Gebirgen.Phytocoenologia 24:53–110
Moore PD(2008)Tropical forests.Facts on File Inc,New York,p 246
Negash M,Starr M(2013)Litterfall production and associated carbon and nitrogen f l uxes of seven woody species grown in indigenous agroforestry systems in the south-eastern Rift Valley escarpment of Ethiopia.Nutr Cycl Agroecosyst 97:29–41
Nigatu L,Tadesse M(1989)An ecological study of the vegetation of the Harenna forest,Bale,Ethiopia.Sinet 12(1):63–93
Pallant J(2001)SPSS survival manual:a step by step guide to data analysis using SPSS for windows.Open University Press, Buckingham,p 334
R Core Team(2013)R:A language and environment for statistical computing.R Foundation for Statistical Computing,Vienna. http://www.R-project.org
Rahbek C(1995)The elevational gradient of species richness:a uniform pattern?Ecography 18(2):200–205
Ritchie JT,Gerakis A,Suleiman A(1999)Simple model to estimate fi eld-measured soil water limits.Trans ASAE 42(6):1609–1614
Sariyildiz T,Anderson JM,Kucuk M(2005)Effects of tree species and topography on soil chemistry,litter quality,and decomposition in Northeast Turkey.Soil Biol Biochem 37:1695–1706
Scatena FN,Bruijnzeel LA,Bubb P,Das S(2010)Setting the stage. In:Bruijnzeel LA,Scatena FN,Hamilton LS(eds)Tropical montane cloud forests:science for conservation and management.Cambridge University Press,Cambridge,pp 3–13
Schnitzer M(1982)Organic matter characterization.In:Page AL, Miller RHA,Keeney DR(eds)Methods of soil analysis.Part 2: Chemical and microbiological properties.American Society of Agronomy,Inc.and Soil Science Society of America,Inc., Madison,pp 581–594
Senbeta F,Denich M(2006)Effects of wild coffee management on species diversity in the Afromontane rainforests of Ethiopia.For Ecol Manag 232:68–74
Smith JMB(1977)Vegetation and microclimate of east and westfacing slopes in the grasslands of MT Wilhelm,Papua New Guinea.J Ecol 65(1):39–53
Tadesse M,Nigatu L(1996)An ecological and ethnobotanical study of wild or spontaneous coffee,Coffea arabicain Ethiopia.In: van der Maesen JG,van der Burgt XM,van Medenbach de Rooy JM(eds)The biodiversity of african plants.Kluwer,Dodrecht, pp 277–294
Tallents LA,Macdonald DW(2011)Mapping high-altitude vegetation in the bale mountains.Walia,Ethiopia,pp 97–117.doi:10. 1111/j.1365-2656.2011.01911.x
Teketay D,Bekele T(1995)Floristic composition of Wof-Washa natural forest,Central Ethiopia:implications for the conservation of biodiversity.Feddes Repert 106:127–147
Tesfaye G,Teketay D,Fetene M(2002)Regeneration of fourteen tree species in Harenna forest,southeastern Ethiopia.Flora 197:461–474
Tesfaye G,Teketay D,Assefa Y,Fetene M(2004)The impact of f i re on the soil seed bank and regeneration of Harenna forest, southeastern Ethiopia.Mt Res Dev 24(4):354–361
Teshome E,Randall D,Kinahan A(2011)The changing face of the Bale Mountains National Park over 32 years:a study of land cover change.Walia,Ethiopia,pp 118–130
Uhlig SK(1988)Mountain forests and the upper tree limit on the southeastern plateau of Ethiopia.Mt Res Dev 8(2/3):227–234
Uhlig SK,Uhlig K(1991)Studies on the altitudinal zonation of forests and alpine plants in the central Bale Mountains,Ethiopia. Mt Res Dev 11(2):153–156
Wesche K(2003)The importance of occasional droughts for afroalpine landscape ecology.J Trop Ecol 19:197–208
Wesche K,Miehe G,Kaeppeli M(2000)The signif i cance of f i re for afroalpine ericaceous vegetation.Mt Res Dev 20(4):340–347
Yadav AS,Gupta SK(2006)Effect of micro-environment and human disturbance on diversity of woody species in the Sariska tiger project in India.For Ecol Manag 225:178–189
Yimer F,Ledin S,Abdelkadir A(2006a)Soil property variations in relation to topographic aspect and vegetation community in the south-eastern highlands of Ethiopia.For Ecol Manag 232:90–99
Yimer Y,Ledin S,Abdelkadir A(2006b)Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains,Ethiopia.Geoderm 135:335–344
Yimer Y,Ledin S,Abdelkadir A(2007)Change in soil organic carbon and total nitrogen content in three adjacent land use types in the Bale Mountains,south-eastern highlands of Ethiopia.For Ecol Manag 242:337–342
Zhang D,Hui D,Luo Y,Zhou G(2008)Rates of litter decomposition in terrestrial ecosystems:global patterns and controlling factors. J Plant Ecol 2:85–93
17 December 2014/Accepted:31 January 2015/Published online:7 October 2015
Project funding:This study was f i nancially supported by the Department of Forest Sciences,University of Helsinki.
The online version is available at http://www.springerlink.com
Corresponding editor:Zhu Hong
✉Eshetu Yirdaw
eshetu.yirdaw@helsinki.f i
1Department of Forest Sciences,University of Helsinki, P.O.Box 27,00014 Helsinki,Finland
2Wondo Genet College of Forestry and Natural Resources, Hawassa University,P.O.Box 128,Shashemene,Ethiopia
杂志排行

Journal of Forestry Research的其它文章
- Drone remote sensing for forestry research and practices
- Life cycle environmental impact assessment of biochar-based bioenergy production and utilization in Northwestern Ontario, Canada
- Growth rates of Eucalyptus and other Australian native tree species derived from seven decades of growth monitoring
- Effect of f i rst thinning and pruning on the individual growth of Pinus patula tree species
- The inf l uence of selective cutting of mixed Korean pine(Pinus koraiensis Sieb.et Zucc.)and broad-leaf forest on rare species distribution patterns and spatial correlation in Northeast China
- Modeling forest f i res in Mazandaran Province,Iran