新型排胶调节剂对橡胶树无性系热研8—79的增产效应
2015-04-29李言史敏晶陈月异田维敏
李言 史敏晶 陈月异 田维敏


中国热带农业科学院橡胶研究所农业部橡胶树生物学与遗传资源利用重点实验室省部共建国家重点实验室培育基地-海南省热带作物栽培生理学重点实验室 海南儋州 571737
摘 要 以橡胶树无性系热研8-79为材料,分析一种新型排胶调节剂对胶乳产量、干胶产量、茎围增长、树皮结构和死皮发生的影响。结果表明:新型排胶调节剂在高温季节有明显的增产效应,胶乳产量和干胶产量分别增长23.38%和16.01%;新型排胶调节剂能有效降低死皮发生率,仅为2.22%,而且能促进乳管分化,对茎围增长没有明显的抑制效应。结果说明新型排胶调节剂具有高效性和安全性。
关键词 巴西橡胶树;新型排胶调节剂;天然橡胶产量;死皮;树皮结构
中图分类号 S794.1 文献标识码 A
Effect of a Novel Latex Flow Regulator on
Increasing Rubber Yield of Rubber
Tree Clone Reyan8-79
LI Yan, SHI Minjing, CHEN Yueyi, TIAN Weimin*
Rubber Research Institute, CATAS / Key Laboratory of Biology and Genetic Resources of Rubber Tree, Ministry of Agriculture /
State Key Laboratory Incubation Base for Cultivation & Physiology of Tropical Crops, Danzhou, Hainan 571737, China
Abstract The effect of a novel latex flow regulator on the latex production, rubber yield, girth increment, bark structure and occurrence of tapping panel dryness(TPD)were analyzed. Results showed that the regulator caused the latex production and rubber yield to be increased by 23.38% and 16.01% in the moths with relatively high temperature, respectively. It also reduced the TPD occurrence. The rate of TPD occurrence was 2.22%, the lowest in comparison with 0.5% ethrel and that without any treatments. And moreover, it could induce the secondary laticifer differentiation and had little influence on stem girth increment. Therefore, the novel regulator is characterized by its high efficiency and safety.
Key words Hevea brasiliensis; Novel latex flow regulator; Rubber yield; Tapping panel dryness(TPD); Bark structure
doi 10.3969/j.issn.1000-2561.2015.10.010
巴西橡胶树(Hevea brasiliensis Müll. Arg.)是天然橡胶的主要来源。割胶是天然橡胶生产的必须环节,占天然橡胶生产成本的50%~60%[1-2]。乙烯利刺激割胶技术的发明和应用显著提高了每刀次的胶乳产量,降低了割胶频率,给天然橡胶生产带来了革命性的变革[3-5]。但是,乙烯利使用不当带来的负面效应也日益明显,如排胶时间过度延长、树皮衰老和死皮等[6-10]。因此,天然橡胶生产迫切需要高效安全的新型排胶调节剂。虽然从上世纪五十年代以来人们即在这方面做了大量工作[11],但迄今为止,乙烯利依然是一种最有效的排胶调节剂。最近,笔者们基于乳管伤口堵塞机制的研究结果[12-14],研发出一种排胶调节剂配方。本研究以大田定植的橡胶树无性系热研8-79为材料,分析该配方对胶乳产量、茎围增长、树皮结构和死皮防控等方面的效应,旨在评价该配方的大田增产效应和安全性。
1 材料与方法
1.1 材料
橡胶树无性系热研8-79于2001年定植在海南省儋州市中国热带农业科学院试验场七队。该林段的定植规格3 m×7 m,2009年开割,采用S/2d/3非刺激割制。到2014年试验开始时,该林段进入第6割年。
1.2 方法
1.2.1 材料处理 2014年6月,测定试验林段健康树5刀的胶乳产量,作为分组的依据。试验树分3组,其中不作任何处理的为阴性对照组,用0.5%乙烯利处理的为阳性对照组,用新型排胶调节剂(主要成分是细胞松弛素D和茉莉酸甲酯)处理的为处理组,每组45株。试验从2014年7月1日开始,当天割胶后,下午在割线处涂施相应试剂。按照S/2d/3割制,每处理1次割5刀,即一个刺激周期。测量每株每刀的胶乳产量。用于其它分析的胶乳样品为每5株的混合样,也就是每个处理的每刀有9个混合样,即9个生物学重复。收集的胶乳样品在冰浴条件下带回实验室测定。
1.2.2 干胶含量和产量测定 从每个胶乳混合样中取1 g胶乳,用5%的冰醋酸凝固,再用水漂洗过夜,然后放入60 ℃的烘箱中烘干至恒重,测定干胶含量。按照下列公式计算干胶产量。
干胶产量=胶乳产量×干胶含量
1.2.3 死皮调查 按照袁坤等[15]的方法进行死皮调查。根据橡胶树割胶技术规程鉴定死皮级别,0级:无病;一级死皮:死皮长度在2 cm以下;二级死皮:死皮长度在2 cm至全割线长的1/4;三级死皮:死皮长度为割线长的1/4~1/2;四级死皮:死皮长度为割线长的1/2~3/4;五级死皮:死皮长度为割线长的3/4~全线。由于每年的10至12月份是死皮高发期[16],因此,本试验在9至12月份期间,每个月的月末逐株跟踪调查。
1.2.4 茎围测量 试验开始前,测量距地面1.5 m处的茎围。停割后,测量健康植株距地面1.5 m处茎围。年增长量即为停割后茎围与开割前茎围的差值。
1.2.5 光学显微镜技术 为了观测新型排胶调节剂对树皮结构的影响,以定植于实验场五队的5 a生实验树热研8-79为材料。选取9株,分3组。1组不做任何处理,作为阴性对照;另2组分别用新型排胶调节剂和0.5%乙烯利处理。按照S/2d/3割制,每处理1次割3刀,共处理5次。然后,在每株树割线下用直径1.5 cm的打孔器取1块树皮制作标本(标本用80%的酒精固定,经酒精系列脱水,用碘-溴处理显示乳管,石蜡包埋),再对标本进行横切,固绿衬染后,Leica研究显微镜(DMLB,Wetzlar, Germany)观察照相。
1.2.6 月平均温度观测 月平均温度数据由农业部儋州热带作物科学观测实验站提供。
1.3 数据处理
使用SAS8.1软件进行统计分析。采用One-way ANOVA中的Duncan多重对比分析不同数据组间的差异。
2 结果与分析
2.1 新型排胶调节剂对胶乳产量、干胶含量和干胶产量的影响
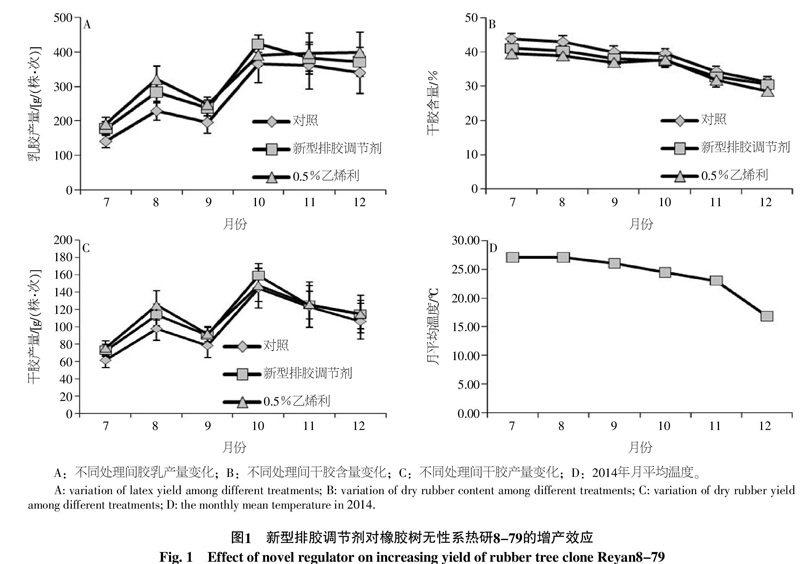
总体上,新型排胶调节剂处理的株次胶乳产量月变化趋势与阴性和阳性对照相似(图1-A)。除9月份外,新型排胶调节剂、阴性和阳性对照的胶乳产量都在逐月增加。胶乳产量在9月份显著下降很可能是台风“海鸥”的影响所致。7~9月份,与阴性对照相比,新型排胶调节剂处理的株次胶乳产量增长率为23.38%,而阳性对照的增长率为34.14%,绝对株次胶乳的增长量分别为43.73 g和64.8 g。10月份,无论是新型排胶调节剂,还是阴性和阳性对照,株次胶乳产量都较7~9月份显著增加,而且新型排胶调节剂处理的株次胶乳产量为422.8 g,超过了阳性对照的390.7 g。11~12月份,新型排胶调节剂、阴性和阳性对照的株次胶乳产量虽然有所下降,但仍显著高于7~9月份的胶乳产量。统计分析可知:7~10月份,新型排胶调节剂的株次胶乳产量都显著高于阴性对照,但在11~12月份,两者差异不显著;在7~9月及12月份,阳性对照的株次胶乳产量都显著高于阴性对照,但在10和11月份,两者差异不显著。
与株次胶乳产量逐月增加不同,干胶含量表现出逐月降低的趋势,其中在11~12月份,干胶含量大幅度下降(图1-B)。新型排胶调节剂的株次干胶含量介于阴性和阳性对照之间。在7~10月份,虽然新型排胶调节剂的株次干胶含量均显著低于阴性对照,但在11~12月份,两者没有明显差异。而在7~12月份,阳性对照的株次干胶含量都显著(10月份)和极显著(其他月份)低于阴性对照。
在7~10月份,株次干胶产量月变化趋势与株次胶乳产相似(图1-C)。虽然11~12月份的株次胶乳产量维持在较高水平上,但是由于干胶含量的大幅度降低,使得干胶产量明显降低(图1-B、C)。7~9月份,无论是新型排胶调节剂还是阳性对照的株次干胶产量均显著高于阴性对照,株次干胶产量较阴性分别增加16.01%和 22.59%。但三者在10~12月份的株次干胶产量没有显著差异。株次干胶含量的月变化趋势与气温的月变化趋势相吻合(图1-D)。随着气温的降低,排胶时间延长,株次干胶含量降低。
2.2 新型排胶调节剂对树皮结构的影响
长期使用乙烯利刺激对橡胶树树皮结构有不良的影响,主要表现在有输导功能次生韧皮部的厚度变小和外层乳管衰老死亡2个方面[17]。在本研究中,只是在5 a生的幼树上做短期试验,而且使用低浓度乙烯利(0.5%),因此,乙烯利刺激的负面效应不明显。但是阴性对照、新型排胶调节剂和阳性对照的树皮结构仍表现出一定程度的差异:在有功能次生韧皮部厚度方面,阴性对照和阳性对照之间没有明显差异(图2-A、C),但是新型排胶调节剂处理的有功能次生韧皮部厚度明显增厚(图2-B);在乳管分化方面,阴性对照在挨近形成层的次生韧皮部中没有出现次生乳管(图2-A),而在新型排胶调节剂处理和阳性对照挨近形成层的次生韧皮部中都出现了次生乳管(图2-B、C),而且新型排胶调节剂处理的更为明显(图2-B)。
2.3 新型排胶调节剂对茎围增长的影响
新型排胶调节剂处理的植株茎围平均增长量虽然大于阴性和阳性对照,但是三者没有显著差异(表1)。其原因主要有2个方面:一是时间不长,仅6个月;二是株间差异较大,导致标准差较大。
2.4 新型排胶调节剂对死皮发生的影响
9~12月份,新型排胶调节剂处理的仅在10月份出现1株死皮树,死皮发生率为2.22%(图3-A),死皮级别为二级(图3-B),在12月份,该植株的死皮程度加重为三级(图3-C);阴性对照在11月和12月分别新增1株一级死皮树,死皮发生率为4.44%(图3-A),死皮级别为一级;阳性对照在10月份出现1株死皮树,在12月份又出现2株死皮树,死皮发生率为6.66%(图3-A)。其中,在10月份出现的死皮树的死皮级别为四级,在12月份加重为五级(图3-D),而12月份出现的2株死皮树的死皮级别为二级。
3 讨论与结论
乙烯利刺激增产的主要原因是显著延长橡胶树的排胶持续时间[18]。橡胶树的排胶持续时间长短取决于2个因素,即排胶动力和排胶阻力。通常认为,乙烯利一方面通过促进水分从乳管周围组织进入乳管,维持一定的膨压和胶乳的稀释效应而增大排胶动力[19],另一方面通过提高黄色体的稳定性而延缓乳管伤口末端橡胶凝块的形成[20-24],减小排胶阻力。但是,也有证据表明,在使用高浓度乙烯利或气刺微割条件下,黄色体破裂指数显著增加,而排胶时间显著延长[25-26]。这说明乙烯利刺激的这种效果不能用延缓橡胶凝块的形成来解释。郝秉中等[27]发现,在刚停止排胶的乳管伤口末端有大量蛋白质性质的网状结构(被称为“蛋白质网”)和完整橡胶粒子的聚集物,没有橡胶凝块。这至少说明有相当一部分的乳管在刚停止排胶时不是被橡胶凝块堵塞的。笔者发现“蛋白质网”的主要成分是来自黄色体破裂后释放的3种主要蛋白质:橡胶素、几丁质酶和β-1,3-葡聚糖酶以及这些蛋白质之间及其与橡胶粒子的相互作用形成的[14]。这些蛋白质网和橡胶粒子聚集物形成的堵塞物通过β-1,3-葡聚糖酶锚定在乳管细胞的细胞骨架上[28]。本新型排胶调节剂的主要成分就是具有解聚细胞骨架功能的细胞松弛素D和具有诱导乳管分化作用的茉莉酸甲酯。因此,在理论上,既有通过阻断β-1,3-葡聚糖酶与肌动蛋白的互作,延缓堵塞物在乳管伤口处的积累,从而减少排胶阻力,又可以促进乳管分化,增加乳管数量,从而增大产胶潜力。大田试验结果表明,该调节剂在7~9月份具有明显的增产效应,但是在10~12月份低温季节,新型排胶调节剂的株次干胶产量均与不做任何处理(阴性对照)的株次干胶含量没有显著差异。其主要原因是低温导致橡胶树排胶时间延长[29]。乙烯利刺激的增产效应也主要发生在7~9月份。这些结果表明,在10月份以后,没有必要再施用调节剂或乙烯利,这为生产上合理使用乙烯利提供了科学依据。虽然新型排胶调节剂的大田增产效应与乙烯利相似,但是仍具有以下特点:(1)干胶含量介于阴性对照和阳性对照之间,而且在11~12月份的干胶含量与阴性对照没有显著差异;(2)促进新生乳管发育的效应明显强于乙烯利刺激;(3)增加有输导功能的次生韧皮部的厚度;(4)死皮发生率低。这些特点对橡胶树持续高产稳产至关重要。
参考文献
[1] 张 得. 海南或现弃割现象 天然橡胶成本几何[J]. 世界热带农业信息, 2014(10): 1-4.
[2] 何晓伟. 2015年国产天然橡胶市场分析[J]. 中国橡胶, 2015(2): 18-21.
[3] Abraham P D, P'ng T C, Ng E K. RRIM Ethrel trials: progress report, in Proc RRIM Plant Conf[R]. Rubber Research Institute Malaya, Kuala Lumpur, 1971.
[4] Dickenson P B, Sivakumaran S, Abraham P D. Ethad and other new stimulants for Hevea brasiliensis, Proc Int Rubber Conf[R]. Rubber Research Institute Malaysia, Kuala Lumpur, 1976.
[5] Sivakumaran S, Abraham P D, Ching P T, et al. Comparative performance of Ethad and Ethephon in field trials[R]. Int Rubber Res Dev Board Symp, Kuala Lumpur, 1978.
[6] Sharples A, Lambourne J. Field experiments relating to brown bast disease of Hevea brasiliensis[J]. Malayan Agr J, 1924, 12: 190-343.
[7] 范思伟, 杨少琼. 橡胶树死皮的概念、 假说和发病机理[J]. 热带作物研究, 1991, 15: 73-80.
[8] 许文献, 魏小弟, 校现周, 等. 刺激割胶制度对橡胶树死皮病发生的生理效应[J]. 热带作物学报, 1995, 16(2): 9-14.
[9] 覃宝祥, 胡新文, 邓晓东, 等. 橡胶树死皮病的形成及其机制[J]. 植物生理学通讯, 2005, 41(6): 827-830.
[10] 俞时举, 林位夫. 橡胶树死皮发生机理研究现状及展望[J]. 安徽农业科学, 2008, 36(17): 7 299-7 300.
[11] Baptist E D C, De Jonge P. Stimulation of yield in Hevea brasiliensis Müll. Arg. Effect of synthetic growth substances on yield and on bark renewal[J]. J Rubber Res Inst Malaya, 1955, 14: 362-367.
[12] 郝秉中, 吴继林, 谭海燕. 橡胶树乳管切割后的堵塞研究[J]. 热带作物学报, 1996, 17(1): 1-6.
[13] Hao B Z, Wu J L, Meng C X, et al. Laticifer wound plugging in Hevea brasiliensis: The role of protein-network with rubber particle aggregations in stopping latex flow and protecting wounded laticifers[J]. J Rubb Res, 2004, 7(4): 281-299.
[14] Wang X C, Shi M J, Wang D, et al. Comparative proteomics of primary and secondary lutoids reveals that chitinase and glucanase play a crucial combined role in rubber particle aggregation in Hevea brasiliensis[J]. J Proteome Res, 2013, 12: 5 146-5 159.
[15] 袁 坤, 谢贵水, 杨礼富, 等. 不同药剂处理对橡胶树死皮和产量的影响[J]. 西南农业学报, 2013, 26(4): 1 524-1 526.
[16] 叶德林, 王昆成, 刘黔英. 浅析冬季低温割胶与橡胶树死皮的增长[J]. 热带农业科学, 2014, 34(9): 52-54.
[17] 吴继林, 谭海燕, 郝秉中. 乙烯利过度刺激采胶诱导巴西橡胶树割面干涸病的研究[J]. 热带作物学报, 2008, 29(1): 1-9.
[18] dAuzac J. The hormonal stimulation of latex yield[C]. // dAuzac J, Jacob J L, Chrestin H(Eds). Physiology of Rubber Tree Latex. CRC Press, Florida, 1989: 289-293.
[19] Tungngoen K, Kongsawadworakul P, Viboonjun U, et al. Involvement of HbPIP2;1 and HbTIP1;1 aquaporins in ethylene stimulation of latex yield through regulation of water exchanges between inner liber and latex cells in Hevea brasiliensis[J]. Plant Physiol, 2009, 151: 843-856.
[20] Southorn W A. Latex flow study. I. Electron microscopy of Hevea brasiliensis in the region of the tapping cut[J]. J Rubb Res Inst Malaya, 1968, 20: 176-186.
[21] Southorn W A, Edwin E E. Latex flow studies. II. Influence of lutoids on the stability and flow of Hevea latex[J]. J Rubb Res Inst Malaya, 1968, 20: 187-200.
[22] Coupe M, Chrestin H. Physico-chemical and biochemical mechanisms of the hormonal(ethylene)stimulation[C].//Physiology of Rubber Tree Latex. Auzac J D, Jacob J L, Chrestin H. Boca Raton: CRC Press, 1989: 295-319.
[23] Gidrol X, Chrestin H, Tan H L, et al. Hevein, a lectin-like protein from Hevea brasiliensis(Rubber tree)is involved in the coagulation of latex[J]. J Biol Chem, 1994, 269: 9 278-9 283.
[24] Wititsuwannakul R, Pasitkul P, Kanokwiroon K, et al. A role for a Hevea latex lectin-like protein in mediating rubber particle aggregation and latex coagulation[J]. Phytochemistry, 2008, 69: 339-347.
[25] 校现周, 罗世巧, 许闻献, 等. 微割对橡胶无性107产量及生理状况的影响[J]. 热带作物学报, 199, 20(1): 8-12.
[26] 校现周, 蔡 磊. 乙烯利刺激对橡胶树乳管细胞活性氧代谢的影响[J]. 热带作物学报, 2003, 24(1): 1-7.
[27] Hao B Z, Wu J L, Meng C X, et al. Laticifer wound plugging in Hevea brasiliensis: The role of protein-network with rubber particle aggregations in stopping latex flow and protecting wounded laticifers[J]. J Rubb Res, 2004, 7: 281-299.
[28] 邓顺楠. 巴西橡胶树乳管伤口封闭物积累的研究[D]. 海口: 海南大学, 2013.
[29] 史敏晶, 程 成, 田维敏. 低温促进巴西橡胶树排胶的生理基础[J]. 热带作物学报, 2015, 36(1): 92-96.
