杂交兰品种‘韩国小姐’与墨兰品种‘企剑黑墨’杂交坐果及无菌播种研究
2015-04-29曾碧玉常强许传俊
曾碧玉 常强 许传俊


摘 要 以大花蕙兰与墨兰的杂交品种‘韩国小姐为母本,墨兰品种‘企剑黑墨为父本进行杂交,对其杂交坐果、落果情况及种子的无菌播种方法进行研究。结果表明:授粉后0~112 d落果较为严重,150 d时坐果率为62.1%。果荚采收时期及培养基对种子的无菌萌发影响显著,种子在授粉后194 d采收,且于不添加植物生长调节剂的1/2MS培养基中培养时萌发最佳。
关键词 杂交兰;墨兰;杂交;坐果率;无菌播种
中图分类号 S682.31 文献标识码 A
Abstract Cymbidium‘Miss Korea, the hybrid of Cymbidium hybridium and Cymbidium sinense, was crossed with Cymbidium sinense‘Qijian Heimo as male parent. The fruit set and drop and the aseptic seeding method of this cross combination were studied. The results showed that the fruit set was 62.1% on 150 days after pollination. The fruits dropped seriously during 0-112 days after pollination. Fruit harvesting time and nutrient media significantly influenced the germination frequency. Seeds, which harvested on 194 days after pollination, germinated best on half strength Murashige and Skoog(1/2MS) medium without plant growth regulators.
Key words Hybrid;Cymbidium sinense;Interspecies cross;Furit set;Aseptic seeding
doi 10.3969/j.issn.1000-2561.2015.03.011
大花蕙兰(Cymbidium hybridium)与墨兰(C. sinense)均属兰科(Orchidaceae)兰属(Cymbidium)植物。大花蕙兰为兰属植物中一部分大花附生种类及其杂交种[1]。株型高大优美,花朵数多且排列整齐、花大色艳,具极高的观赏价值。墨兰为兰属地生兰类植物,五大类传统国兰之一,其株型典雅、叶形花姿优美、幽香四溢,深受人们喜爱。由于大多数大花蕙兰多原产于热带、亚热带高海拔地区,气温较冷凉。在国内低海拔地区度夏较困难,且不具香味。而墨兰原产于中国、越南和缅甸,具有清香典雅、株型及叶型优美、易于栽培种植开花、耐热且抗性强等优点。因此,常用墨兰作亲本用于培育优良大花蕙兰品种。
由于大花蕙兰品种遗传背景及其倍性复杂化,有二倍体、三倍体、四倍体及非整数倍体[2-4],大花蕙兰自交、品种间杂交及以大花蕙兰为母本与墨兰的杂交结果率均较低[2,4-9],难以成功获得杂交组合。而曾碧玉[2]研究结果显示大花蕙兰与墨兰的杂交种为母本与墨兰杂交,结实率可达76.2%。从中可推测,利用大花蕙兰与墨兰的杂交种为母本与墨兰杂交,可大幅度提高杂交结实率,利于杂交组合的获得。种子成熟度及生长培养基是种子萌发的重要影响因子,这在兰科植物不同属中均普遍存在,如兰属[9-11]、石斛兰属(Dendrobium)[12-14]、兜兰属(Paphiopedilum)[15-17]、杓兰属(Cypripedium)[18-19]等。而即使在相同的大花蕙兰与墨兰种间杂交中,不同组合间果荚采收适期及生长培养基均有所差异[2,8],因此对不同杂交组合的种子采收适期及培养基均需要进一步的摸索。
本研究选取大花蕙兰与墨兰杂交选育出的优良品种‘韩国小姐(Cymbidium‘Miss Korea)为母本,与广东四大墨兰之一的‘企剑黑墨(C. sinense ‘Qijian Heimo)为父本进行杂交,并对其杂交坐果、果荚生长发育动态及种子萌发进行研究,为兰属植物的杂交育种提供参考依据。
1 材料与方法
1.1 材料
杂交兰品种‘韩国小姐(Cymbidium‘Miss Korea)与墨兰品种‘企剑黑墨(C. sinense‘Qijian Heimo)均从福建省厦门市亚热带花卉中心购买,然后种植保存于福建省亚热带植物研究所兰花资源圃内。‘韩国小姐每盆假鳞茎4~6个,每个假鳞茎带叶6~9片, 叶色翠绿; 花枝直立,长53.0~58.0 cm; 每个花枝具13~16朵花,花径5.0~6.0 cm; 花为桃红色具纵纹线, 淡香。‘企剑黑墨每盆5~8个假鳞茎,每个假鳞茎带3~4片叶,叶色墨绿;花枝直立,长34.0~56.0 cm; 每枝含花约6~11朵,花径4.8~5.3 cm; 花为棕褐色并带纵纹线,清香。
1.2 方法
1.2.1 人工授粉及果荚大小测量 以 ‘韩国小姐为母本, ‘企剑黑墨 为父本进行杂交。授粉时,母本取开花后3~4 d的花朵,先用镊子去掉唇瓣及柱头上的花粉块,取待开放的父本花粉块放置于母本的柱头蕊腔内;以上所用工具均用95%酒精消毒。授粉后,观察记录果荚的生长发育动态,并按一定时间间隔测量果荚大小,果长计量为果荚基部至花萼着生处长度,用软尺测量。果宽计量为果荚最大直径,用数显游标卡尺测量。
1.2.2 无菌播种 取授粉后120、194、232 d果荚进行无菌播种。果荚先用流水冲洗干净,然后用75%酒精表面擦拭消毒,0.1%氯化汞消毒约10 min,最后用无菌水冲洗4~5次。将果荚内种子均匀地撒播于培养基中。同时将授粉后120、194 d果荚内种子分成4份,均匀撒播在4种培养基中,即Hyponex、KC[20]、1/2MS[21](大量元素减半)、添加植物生长调节剂(BA 0.5 mg/L+NAA 2 mg/L)的1/2MS培养基。每个处理播种10瓶。以上所用培养基均添加蔗糖30 g/L、蛋白胨3 g/L、椰子汁10%及琼脂6 g/L,并于高压灭菌前将培养基中pH调至5.8。培养室温度为(25±2)℃,光强为30~40 μmol/(m2·s),每天光照12 h。定期观察并记录种子萌发情况,以肉眼可见球状体为启始萌发,并于启动萌发后1个月统计萌发率。
1.3 数据处理
数据分析采用SPSS 11.5 for windows统计分析软件包中的one-way ANOVA(LSD和Duncan检验)软件进行分析。
2 结果与分析
2.1 杂交坐果情况
共授粉了145朵花,分3个区组进行。结果表明,随着授粉后时间的延长,坐果率不断下降,而落果率不断上升。从总的杂交坐果与落果情况来看(表1),授粉后第150天尚存果荚90个,坐果率达62.1%;脱落果荚55个,落果率37.9%。至第259天时,尚存果荚80个,坐果率达55.2%;果荚成熟脱落或开裂,脱落果荚65个,落果率为44.8%。至第328天时,大部分果荚开裂,脱落果荚达130个,落果率达89.7%(表1)。‘韩国小姐ב企剑黑墨 结实挂果周期较长,可长达295 d,甚至更长。从区组杂交坐果与落果情况来看,授粉后至第112天时,坐果率与落果率分别为65.9%与31.5%,与0、295、328 d的坐果率与落果率相比较差异显著,但与150、259 d的坐果率与落果率相比较差异不显著(表1)。可见,授粉后0~112 d落果较为严重。
2.2 果荚的生长发育变化
果长及果宽走势大致相同。授粉后2~3 d可见柱头膨大,萼瓣片开始失水皱缩或卷曲;10~12 d子房开始膨大。而后很长一段时间,果荚不断膨大,属快速增长期。直至授粉后160~180 d时,果荚膨大逐渐趋于平稳,而后出现下降后上升再下降上升的波动(图1)。至授粉后180 d时,果荚大小基本趋于平稳,果长(7.17±0.03)cm,果宽(20.57±0.19)mm。成熟果荚呈棱形,近柱头端稍钝;果皮呈翠绿或翠绿并稍泛黄,果皮表面渐变粗糙,果棱皱缩。
2.3 果荚采收时期及培养基对种子萌发的影响
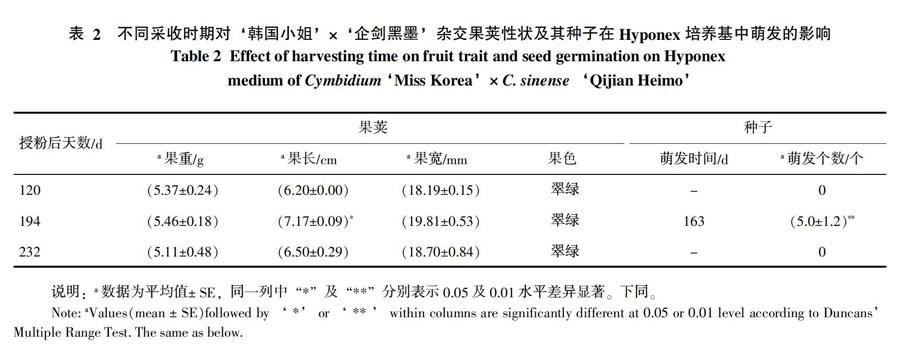
不同采收时期(授粉后120、194、232 d)果荚性状见表2。3个不同时期果荚中,以194 d果荚最大,果重为(5.46±0.18)g,果长为(7.17±0.09)cm,果宽为(19.81±0.53)mm。虽然3个时期果荚色泽均翠绿,但232 d 果荚果棱明显呈现皱缩状态。不同采收时期对种子萌发影响差异显著。目前(播种后180 d)为止,未见120、232 d种子萌发;仅见194 d种子萌发,并在0.01水平达极显著差异(表2)。可见,以授粉后194 d果荚采收进行无菌播种较佳。
由表3可知,不同培养基对种子萌发影响差异显著。在4种培养基中,以1/2MS为基本培养基,萌发较快,最早萌发所需时间为132 d。而在Hyponex及KC培养基中萌发较晚,为163 d。在4种培养基中均可观察到原球茎与根状茎,且均健壮。但以1/2MS中培养萌发个数最多,平均达25.3个,在0.01水平达极显著差异(表3);且绿色原球茎迅速伸长继而形成多分支状根状茎,表面着生大量白色根毛状物质。而其他3种培养基中观察到的根状茎短且分支少或无分支,表面白色根毛状物质着生少(图2);另外,形成的根状茎具有成苗快的优点,具杂种优势。本研究中,以1/2MS为基本培养基,添加植物生长调节剂(6-BA 0.5 mg/L+NAA 2 mg/L)未能缩短启动萌发所需的时间,也未能提高萌发个数。可见,4种培养基中,‘韩国小姐ב企剑黑墨杂交种子在不添加植物生长调节剂的1/2MS培养基中萌发为佳。此外,本研究结果中还发现,萌发较佳的培养基比萌发欠佳或无萌发的培养基褐化严重。
3 讨论与结论
张东旭[9]的研究结果显示以大花蕙兰和墨兰的杂交后代为母本与大花蕙兰进行杂交,是提高杂交结果率且得到杂交后代的有效途径。本研究中,利用大花蕙兰和墨兰的杂交种‘韩国小姐与墨兰‘企剑黑墨进行杂交,具较高的亲和性,至授粉后第150天时,果荚坐果率达62.1%。结果证明了大花蕙兰与墨兰的杂交种为母本与墨兰杂交能大幅度提高杂交坐果率,并且有利于杂交组合的获得进而改良兰属植物种质。整个果荚生长发育期间,授粉后0~112 d落果较为严重,可能由于无法完成双受精或是胚败育,具体成因还须结合胚胎学进一步深入研究。此外,果荚生长后期出现下降后上升的波动,可能是由于果荚养分吸收变缓或是失水萎缩所致[2,4,7-11,22]。
兰科植物种子成熟度及培养基是种子萌发的重要影响因子。通常种子过于幼嫩或成熟均不利萌发,且成熟种子无菌播种萌发比未成熟种子困难[15-16,19,23-24]。关于其内在休眠机制尚不明确,但目前普遍认为是成熟种子进入休眠期,由于在其发育过程中不断累积萌发抑制物质,同时种胚抗渗性增强,不利于养分吸收[9,15-16, 24-25]。但亦有研究认为是由于内珠被萎缩形成一层致密的膜状物并包围种胚,即‘壳(carapace),影响种胚水分及养分的吸收;壳中还可检测到各种化学成分,诸如木质素、纤维素、木栓质及角质层物质等,增加种子的抗渗性进而影响种子萌发[17-19,24, 26]。本研究中,种子采收时期及培养基对种子萌发影响显著,可见采收时期及培养基均为种子萌发的重要影响因子。授粉后194 d种子培养可萌发,而授粉后120、232 d种子培养未见萌发。可能由于授粉后120 d种子过于幼嫩而授粉后232 d种子已发育成熟进入休眠期而难于萌发,但其具体的休眠机制还有待于进一步的研究验证。
大花蕙兰与墨兰的杂种胚从具有萌发能力后一直保持着这种萌发能力,直至种子完全成熟时仍可正常萌发[8]。大花蕙兰与墨兰杂交,无论是正交还反交,其杂交种子均以原球茎方式萌发[2, 8, 27]。并认为大花蕙兰的原球茎再生性状由显性基因控制,而墨兰的根状茎再生方式由隐性基因控制[27]。本研究中,杂交种子至完全成熟时则无法萌发。种子萌发方式与大花蕙兰与墨兰杂种萌发方式不同,表现为原球茎而后根状茎继而快速成苗的过程,具杂种优势;虽经根状茎,但分化成苗容易且快,与传统兰属地生性兰如春兰、建兰及墨兰等种子萌发形成的根状茎难于成苗有区别。可能是由于本研究中杂交种子特性更倾向于墨兰,更多地表现出地生兰的特性,而大花蕙兰×墨兰杂交种子更多地倾向于大花蕙兰。此种萌发方式与纹瓣兰(C. aloifolium)、冬风兰(C. dayanum)种子萌发方式相类似[28-30]。此外,以建兰(C. ensifolium)为母本与纹瓣兰杂交、纹瓣兰为母本与银边墨兰(C. sinense ‘Yinbian Molan)杂交及杂交兰 ‘韩国桃花 (大花蕙兰×墨兰)为母本与蕙兰(C. faberi)杂交的种子也以此方式萌发[2,6,31]。Yamazaki等[24]在金兰(Cephalanthera falcata)种子非共生的萌发研究中,结果发现在褐化的培养基中种子萌发率最高。本研究中亦发现相类似的现象,即在褐化较严重的培养基中种子萌发较佳,而在微褐化或无褐化的培养基中种子萌发欠佳或无萌发。这可能归因于种子萌发过程中释放出致培养基褐化的物质,但这种致褐化物质与种子萌发的相关性还需进一步研究。
致谢 感谢福建省亚热带植物研究所张庆美副研究员对本研究提供的帮助!
参考文献
[1] 陈心启,吉占和. 中国兰花全书[C]. 北京:中国林业出版社,1998: 100-102.
[2] 曾碧玉. 兰属、 蝴蝶兰属杂交结实率及种胚培养研究[D]. 广州: 华南农业大学, 2006.
[3] 朱根发, 吕复兵, 王碧青,等. 大花蕙兰品种的染色体数目分析[J]. 园艺学报, 2006, 33(2): 417-421.
[4] 王利民. 大花蕙兰杂交育种研究[D]. 北京:北京林业大学,2007.
[5] 张志胜, 何琼英, 傅雪琳,等. 中国兰花远缘杂交及杂交种子萌发的研究[J]. 华南农业大学学报, 2001, 22(2): 62-65.
[6] 朱根发, 王碧青, 陈明莉,等. 大花蕙兰与兰属植物种间杂交研究[J]. 植物学通报. 2005a, 22(4): 445-448.
[7] 曾碧玉, 朱根发,张文惠. 大花蕙兰与墨兰杂交结实率研究初报[J]. 亚热带植物科学, 2010, 39(1): 14-16.
[8] 凌春英. 大花蕙兰和墨兰杂交育种研究[D]. 北京: 北京林业大学, 2010.
[9] 张东旭. 大花蕙兰杂交及种胚萌发研究[D]. 泰安: 山东农业大学, 2010.
[10] 潘银萍. 部分国兰品种杂交育种及种胚萌发的研究[D]. 泰安:山东农业大学, 2011.
[11] 王郑昊. 多花兰与蕙兰杂交及种胚萌发研究[D]. 泰安: 山东农业大学, 2012.
[12] Alam M K, Rashid M H, Hossain M S, et al. In vitro seed propagation of Dendrobium(Demdrobium transparens)Orchid as influenced by different media[J]. Biotechnology, 2002, 1(1):111-115.
[13] 王 伟,靳晓翠,金荷仙,等. 大苞鞘石斛(Dendrobium wardianum)种子离体萌发培养[J]. 种子, 2011, 30(4): 45-49.
[14] Vasudevan R, van Staden J. Fruit harvesting time and corresponding morphological changes of seed integuments influence in vitro seed germination of Dendrobium nobile Lindl[J]. Plant Growth Regulation, 2010, 60(2): 237-246.
[15] 陈之林, 叶秀群, 梁承邺,等. 杏黄兜兰和硬叶兜兰的种子试管苗培养[J]. 园艺学报, 2004, 31(4): 540-542.
[16] 刘其府, 傅燕艳, 曾宋君,等. 同色兜兰种子非共生萌发试验[J]. 广东农业科学, 2012, 39(12): 47-49.
[17] 张娟娟, 严 宁,胡 虹. 三种兜兰属植物种子发育过程及其与无菌萌发的关系[J]. 植物分类与资源学报, 2013, 35(1): 33-40.
[18] Lee Y I, Lee N, Yeung E C, et al. Embryo development of Cypripedium formosanum in relation to seed germination in vitro[J]. Journal of the American Society for Horticultural Science, 2005, 130(5): 747-753.
[19] 邓 莲, 张 毓, 王苗苗,等. 濒危兰科植物大花杓兰种子非共生萌发的研究[J]. 种子, 2012, 31(6): 31-34, 39.
[20] Knudson L A. New nutrient solution for the germination of orchid seed[J]. American Orchid Society Bulletin, 1946, 14(2): 214-217.
[21] Murashige T, Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures[J]. Physiol Plantarum,1962, 15(3): 473-497.
[22] 曾碧玉, 朱根发, 刘海涛. 蝴蝶兰红花系品种间杂交结果率研究[J]. 华南农业大学学报, 2007, 28(1): 117-119.
[23] Kitsaki C K, Zygouraki S, Ziobora M, et al. In vitro germination, protocorm formation and plantlet development of mature versus immature seeds from several Ophrys species(Orchidaceae)[J]. Plant Cell Report, 2004, 23(2): 284-290.
[24] Yamazaki J, Miyoshi K. In vitro asymbiotic germination of immature seed and formation of protocorm by Cephalanthera falcata(Orchidaceae)[J]. Annals of Botany, 2006, 98(6): 1 197-1 206.
[25] Lee Y I, Lu C F, Chung M C, et al. Developmental changes in endogenous abscisic acid concentrations and asymbiotic seed germination of a terrestrial orchid, Calanthe tricarinata Lindl[J]. Journal of the American Society for Horticultural Science,2007, 132(2): 246-252.
[26] 张 毓, 张启翔, 赵世伟,等. 濒危植物大花杓兰胚与珠被发育的研究[J]. 园艺学报, 2010, 37(1): 72-76.
[27] 朱根发, 陈明莉, 罗智伟,等. 墨兰与大花蕙兰种间杂种原球茎的诱导及增殖研究[J]. 园艺学报, 2004, 31(5): 688-690.
[28] Nayak N R, Chand P K, Rath S P, et al. Influence of some plant growth regulators on the growth and organogenesis of Cymbidium aloifolium(L.)Sw. seed-derived rhizomes in vitro[J]. In Vitro Cellular & Developmental Biology-Plant, 1998, 34(3): 185-188.
[29] Chang C, Chen Y C, Yen H F. Protocorm or rhizome? The morphology of seed germination in Cymbidium dayanum Reichb[J]. Botanical Bulletin of Academia Sinica. 2005, 46:71-74.
[30] 朱根发, 王碧青,吕复兵. 建兰与纹瓣兰种间杂种胚培养研究[J]. 热带亚热带植物学报, 2005b, 13(5): 447-450.
[31] 陈瑶瑶, 张 燕, 张 琛, 等. 杂交兰 ‘韩国桃花×蕙兰种间杂交种子无菌萌发特征研究[J]. 园艺学报, 2009, 36(3): 441-446.
