凤凰单丛茶树资源遗传多样性的ISSR分析
2015-04-29吴清等
吴清等
摘 要 采用ISSR分子标记技术对48份凤凰单丛茶品种(系)进行了遗传多样性分析。选用10个多态性高、分辨力强的ISSR引物分别对供试材料基因组DNA进行扩增,共获得121个位点,其中多态位点带98个,多态位点比率(PPB)为81.0%。品种(系)之间的遗传相似系数变化范围在0.590 9~0.967 4之间。UPGMA聚类分析结果表明,在GS值为0.792的水平上供试材料可划分为7大类,其亲缘关系远近与地理来源关系不大且与香味类型没有必然的联系。
关键词 ISSR分子标记;凤凰单丛茶;遗传多样性
中图分类号 S571.1 文献标识码 A
Abstract ISSR molecular markers were used to analyze the genetic polymorphism of 48 Fenghuang-Dancong tea plant germplasms. Using 10 primers, we amplified 121 bands, of which 98 bands are polymorphic, and the average percentage of polymorphic bands was 81.0%. The genetic similarity among all the tested tea plant germplasms ranged from 0.590 9 to 0.967 4. Based on the genetic similarity and the UPGMA cluster, all the 48 accessions were clustered to 7 groups on the genetic similarity level of 0.792. The dengrogram indicated that the genetic of 48 tested tea plant germplasms had no certain relation with the geographical origin and aroma type.
Key words ISSR markers; Fenghuang-Dancong Tea; Genetic diversity
doi 10.3969/j.issn.1000-2561.2015.03.009
凤凰单丛茶属乌龙茶类,产于“中国乌龙茶之乡”的广东省潮安县凤凰镇,由茶农历代沿习单株优选、单株培育、单株采制的传统栽培方式和独特的加工工艺制作而成的具有天然花香和特殊韵味品质的乌龙茶,现已形成了具有区域特色的品种(系)体系,享有“中国的国宝”之美誉[1]。茶业产量及品质提高依赖于茶树优良品种的选育,而凤凰单丛品种(系)资源繁多,且主要以香型为主线将其归类,存在同一香型中包含多个品种(系),同型品种(系)间的品质参差不齐的现象,加之茶树通常是通过杂交育种,从而使各品种(系)之间的亲缘关系混乱,本底不清,遗传背景不明确,极大地妨碍了品种(系)之间的亲缘归类、育种亲本的选配和种质资源的创新和利用。
ISSR(Inter-Simple Sequence Repeat)是一种以基因组中常出现的SSR本身设计成锚定引物为单引物并基于PCR技术的简单序列重复区间扩增多态性分子标记,具有无需预先知道基因组序列信息、快速、稳定、多态性丰富、重复性强及成本低等优点[2]。该技术广泛应用于植物的种质资源鉴定和亲缘关系分析,在茶树种质资源上的应用也有相关报道[3-8]。本研究利用ISSR分子标记技术,从DNA分子水平对所收集的48份凤凰单丛茶树品种(系)进行遗传多样性及亲缘关系研究,为凤凰单丛茶树品种(系)的亲缘归类、保护及利用提供参考依据。
1 材料与方法
1.1 材料
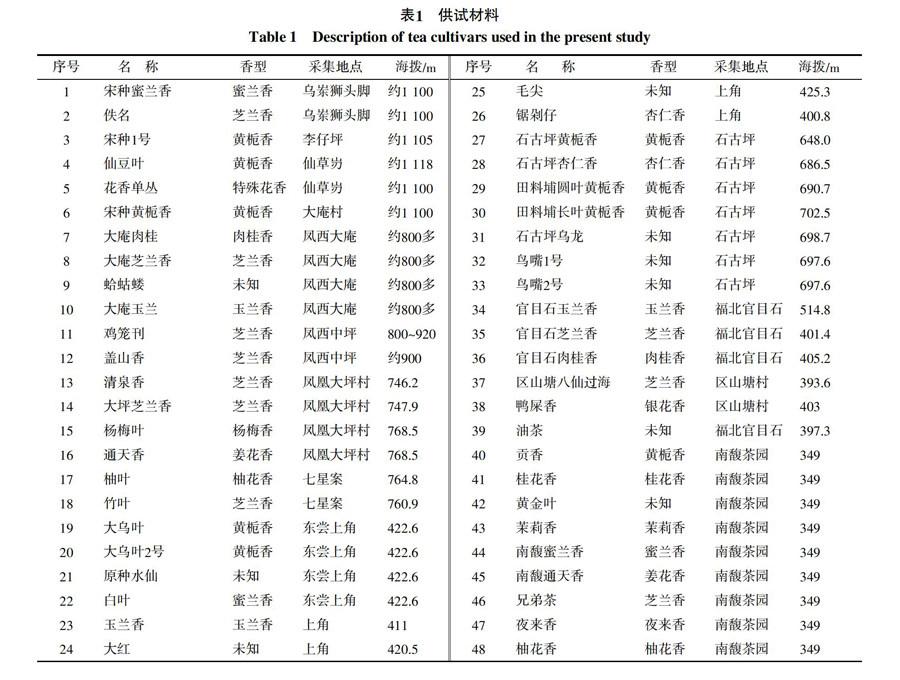
供试的48份凤凰单丛茶树样品及来源详见表1。取一芽两叶置于盛有硅胶的封口袋中,-20 ℃保存备用。ISSR引物由上海生工生物工程技术服务有限公司合成,Taq聚合酶、dNTPs等购自大连宝生物TaKaRa公司。
1.2 方法
1.2.1 DNA的提取 参照Sue Porebski[9]、陈昆松[10]的方法,采用改良的CTAB(2% β-巯基乙醇、3%聚乙烯吡咯烷酮)法进行基因组DNA提取。
1.2.2 ISSR-PCR扩增和产物检测 ISSR-PCR扩增反应在TaKaRa TP600梯度PCR仪上进行,经过比较和优化,确定反应体积为20 μL,PCR组分如下:2 μL 10×PCR buffer、40 ng/μL模板DNA、150 μmol/L dNTPs、1.5 mmol/L MgCl2、引物0.3 μmol/L、1.0 U Taq酶。扩增程序为:94 ℃预变性7 min;94 ℃变性1 min,特定温度退火45 s,72 ℃延伸120 s,38个循环;最后72 ℃延伸10 min。扩增产物用含有0.5 μg/mL EB的1.8%琼脂糖凝胶(1×TAE缓冲液)以5 V/cm电泳3 h,电泳结束用VILBER LOURMAT公司凝胶成像系统照相并保存。
1.3 数据处理
对保存的图片进行人工方法读带,将电泳图上可重复的、清晰的条带赋值为“1”,同一电泳迁移位置无带或不易分辨的弱带赋值为“0”,建立原始数据矩阵。使用NTSYspc2.10e软件,根据Nei等[11]采用DICE系数计算供试材料间的遗传相似系数,并按SAHN邻接法对供试材料进行UPGMA遗传相似性聚类分析,并绘制树状聚类图[5]。
2 结果与分析
2.1 ISSR扩增产物多态性
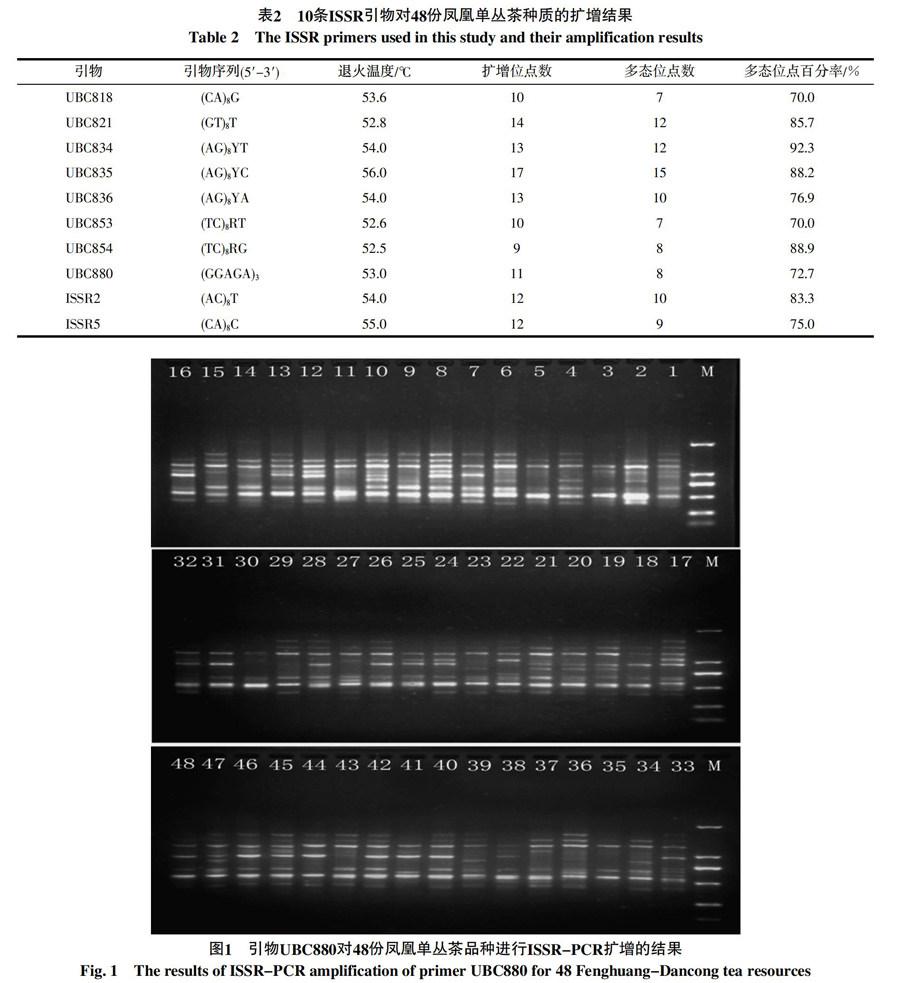
从103条ISSR引物中筛选出10条能在特定的退火温度下扩增条带具有多态性高、重复性强、背景清晰的引物用于ISSR-PCR反应,10条引物共扩增出121个位点,其中多态性位点数为98个,平均多态性位点百分率(PPB)为81.0%(表2)。不同引物扩增的位点数7~15个不等,平均每个引物扩增12.1个位点和9.8个多态位点,多态位点百分率最高为92.3%,最低为70%(表2)。扩增产物片段大小介于250~2 000 bp之间,图1为引物UBC880扩增的结果。
2.2 品种(系)间亲缘关系和聚类分析
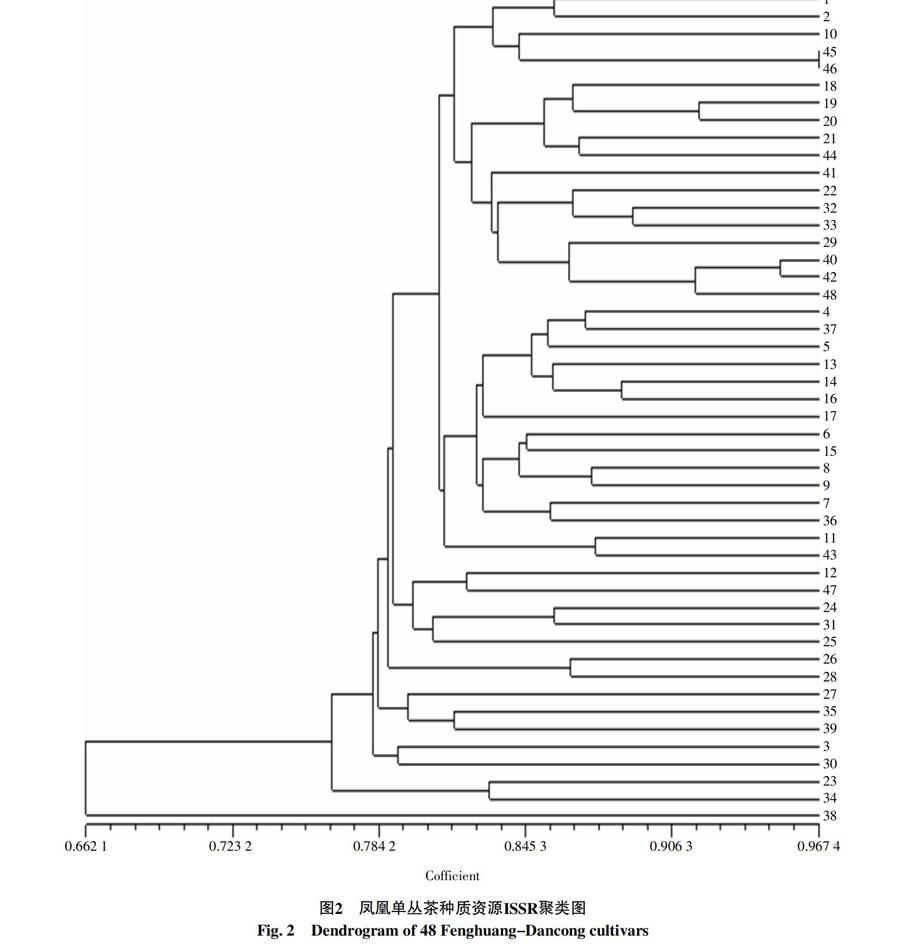
根据条带统计的数据矩阵,分析供试材料间的遗传相似性系数,结果表明,48份凤凰单丛茶树资源品种(系)之间的遗传相似性系数范围介于0.590 9~0.967 4之间,平均遗传相似性系数为0.795 0。其中南馥通天香(45号)与兄弟茶(46号)之间的相似系数最高(0.967 4),亲缘关系最近。而玉兰香(23号)与鸭屎香(38号)之间的相似系数最低(0.590 9),亲缘关系最远,显示它们彼此存在较大的遗传差异。
依据种质间的Neis遗传相似性系数对48份凤凰单丛茶样品进行UPGMA聚类分析,结果(图2)显示,在相似性系数阈值为0.792时,可将48份样品分为7大类群,第Ⅰ类群包括宋种蜜兰香等33份品种(系),第Ⅱ类群包括盖山香、夜来香、大红、石古坪乌龙、毛尖5个品种(系),第Ⅲ类群包括锯剁仔、石古坪杏仁香2个品种(系),第Ⅳ类群包括石古坪黄栀香、官目石芝兰香、油茶3个品种(系),第Ⅴ类群包括宋种1号、田料埔长叶黄栀香2个品种(系),第Ⅵ类群包括玉兰香、官目石玉兰香2个品种(系),第Ⅶ类群包括鸭屎香1个品种(系)。其中第Ⅰ类群还可以分为两个亚类群,第Ⅰ亚类群包括宋种蜜兰香、佚名、大庵玉兰、南馥通天香、兄弟茶、竹叶、大乌叶、大乌叶2号、原种水仙、南馥蜜兰香、桂花香、白叶、鸟嘴1号、鸟嘴2号、田料埔圆叶黄栀香、贡香、黄金叶和柚花香18个品种(系),第Ⅱ亚类群包括仙豆叶、区山塘八仙过海、花香单丛、清泉香、大坪芝兰香、通天香、柚叶、宋种黄栀香、杨梅叶、大庵芝兰香、蛤蛄蝼、大庵肉桂、官目石肉桂香、鸡笼刊和茉莉香15个品种(系)。根据聚类树状图结果表明,供试的凤凰单丛茶种质之间的亲缘关系与地理来源、海拔高度及香味类型没有必然的对应关系,例如同一香型、命名相似的大坪芝兰香、大庵芝兰香、官目石芝兰香3个种质并不完全聚为同一类群,大庵玉兰、玉兰香、官目石玉兰香3个种质也并不完全聚为同一类群,田料埔圆叶黄栀香、田料埔长叶黄栀香、宋种黄栀香、石古坪黄栀香4个种质分别聚类于不同类群中。
3 讨论与结论
本研究选用10条多态性高、分辨力强的ISSR引物对收集到的48份凤凰单丛茶种质DNA进行扩增,得到凤凰单丛茶ISSR带型表现出较高的多态性,多态性带型占81.0%,遗传相似系数介于0.590 9~0.967 4间,遗传相似性变幅为0.376 5,而萧力争[12]等采用AFLP标记法分析34份凤凰单丛古茶树资源遗传多样性得出的多态性比率为79.3%,遗传相似性系数变化范围在0.13~0.49之间,遗传相似性变幅为0.36,这表明ISSR分子标记是评估凤凰单丛茶遗传多样性的有效方法,同时也验证了ISSR分子标记具有快速、稳定、多态性丰富、重复性强等特点[13-15]。Liu[3]等采用ISSR标记分析134云南茶树资源遗传多样性,分析得出其相似系数范围为0.445~0.819,变幅为0.374,说明潮州凤凰镇凤凰单丛茶树资源之间具有较宽的遗传基础,但也存在品种(系)间的遗传距离较小、亲缘关系较近,只有少数品种(系)表现出较为显著的遗传分化,如采摘于南馥茶园的南馥通天香与兄弟茶之间的遗传相似性最高,可能是种植时品种(系)弄混,可将这两个品种(系)作为复份材料保存,而在聚类上,鸭屎香与其他品种(系)单独分开,则有待于进一步鉴定分析。造成这种现象的原因可能是因为长期以来,凤凰单丛茶主要以香型归类,茶农藉此在不同地域之间经常相互引种栽培,出现许多同名异物或同物异名的现象,最终导致品种(系)的名称和分类十分混乱。然而本研究采用ISSR标记方法分析得出,凤凰单丛茶种质间的亲缘关系与其地理来源及香味类型没有必然的联系,此研究结果跟萧力争等[12]采用AFLP标记方法分析得出来结果一致。在类聚上,大乌叶和大乌叶2号,鸟嘴1号和鸟嘴2号,这种遗传相似性高,却体现出微小的遗传分化,这可能是因为是茶树本身为高度杂合体且是异花授粉,在长期杂交演化过程中能产生很多不连续的变异,加上茶树芽叶本身具有突变的可能,同时长时间移植后新环境对品种(系)的遗传信息也可能发生一定的影响[3,16]。
因此,为了保证凤凰单丛茶种质的优良品质,必须明确凤凰单丛茶种质间的遗传背景,避开原来香型归类的影响,根据ISSR分子标记分析结果有助于指导选配工作,选择亲缘关系较远的品种(系),增加其子代的遗传变异,选育出具有杂种优势、综合性状好的优良品种(系)。
参考文献
[1] 叶汉钟, 黄柏梓. 凤凰单丛[M]. 上海: 上海文化出版社, 2010.
[2] Zietkiweica E, Rafalski A, Labuda D. Genome fingerprinting by simple sequence repeat(SSR)-anchored poly-merase chain reaction amplification[J]. Genomics, 1994, 20(2): 176-183.
[3] Liu B Y, Li Y Y, Tang Y C, et al. Assessment of Genetic Diversity and Relationship of Tea Germplasm in Yunnan as Revealed by ISSR Markers[J]. Acta Agronomica Sinica, 2010, 36(3): 391-400.
[4] Modal T K. Assessment of genetic diversity of tea(Camellia sinensis(L.)O.Kuntze)by inter-simple sequence repeat polymerase chain reaction[J]. Euphytica, 2002, 128: 307-315.
[5] Yao M Z, Chen L, Liang Y R. Genetic diversity among tea cultivars from China, Japan and Kenya revealed by ISSR markers and its implication for parental selection in tea breeding programmes[J]. Plant Breeding, 2008, 127: 166-172.
[6] 郭春芳, 唐玉海, 孙 云, 等. 茶树种质资源遗传多样性的ISSR分析[J]. 热带作物学报, 2008, 29(2): 181-186.
[7] 林郑和, 陈荣冰, 陈常颂, 等. ISSR分子标记在茶树遗传关系分析中的初步应用[J]. 茶叶科学, 2007, 27(1): 45-50.
[8] 季鹏章, 汪云刚, 张 俊, 等. 茶组植物亲缘关系的ISSR分析[J]. 西南农业学报, 2009, 22(3): 584-588.
[9] Porebski S, Bailey L G, Baum B R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components[J]. Plant Molecular Biology Reporter, 1997, 15(1): 8-15.
[10] 陈昆松, 李 方, 徐昌杰, 等. 改良CTAB法用于多年生植物组织基因组DNA的大量提取[J]. 遗传, 2004, 26(4): 529-531.
[11] Nei M, Li W H. Mathematical model for studying genetic variation in terms of restriction endonucleases[J]. Proc Natl Acad Sci, 1979, 76(10): 5 269-5 273.
[12] 萧力争, 晏嫦妤, 李家贤, 等. 凤凰单丛古茶树资源的遗传多样性AFLP分析[J]. 茶叶科学, 2007, 27(4): 280-285.
[13] Lan D Godwin, Elisabeth A B Aitken, Lawrence W. Smith. Application of inter simple sequence repeat(ISSR) markers to plant genetics[J]. Electrophoresis, 1997, 18: 1 524-1 528.
[14] McGregor C E, Lambert C A, Greyling M M, et al. A comparative assessment of DNA fingerprinting techniques(RAPD, ISSR, AFLP and SSR)in tetraploid potato(Solanum tuberosum L.)germplasm[J]. Euphytica, 2000, 113: 135-144.
[15] Pradeep Reddy M, Sarla N, Siddiq E A. Inter simple sequence repeat(ISSR)polymorphism and its application in plant breeding[J]. Euphytica, 2002, 128: 9-17.
[16] 赵尹强, 陈朝进, 刘志刚, 等. 茶树芽叶的变异[J]. 中国茶叶, 2011(2): 8-9.
