结缕草克隆生长对生境资源梯度水平和次序的响应
2015-04-11赖苏雯李德志陈惠娟耿松陈青青贠小涛樊治华欧阳义黄瑶瑶王明祖
赖苏雯,李德志,2,3,陈惠娟,耿松,陈青青,贠小涛,樊治华,欧阳义,黄瑶瑶,王明祖
( 1. 华东师范大学生态与环境科学学院,上海200241; 2. 华东师范大学上海市城市化生态过程与生态恢复重点实验室,上海200241;3. 华东师范大学浙江天童森林生态系统国家野外科学观测研究站,上海200241)
克隆植物(Clonal Plant)是指具有克隆性的一类植物[1]。这类植物在自然条件下能够自发地通过营养生长过程形成多个在遗传上一致、在形态和生理上独立或者潜在独立的个体[2]。由于克隆植株常常具有水平空间上的移动性[3],因此,克隆植物通常可以像动物一样“觅食”,从而增加其在异质生境中生长的概率。克隆整合又称为生理整合,是指物质(如水分、矿质养分、光合产物等)或信号通过克隆分株之间的间隔子(如匍匐茎、根茎等)进行传输的过程[4]。克隆整合可以通过调节生物量分配[5]的方式实现形态可塑性变化,从而有效响应生境的异质性。克隆植物如何响应生境异质性并使自身生长获益最多,已经成为很多学者研究的焦点问题之一。
环境资源异质性是指包括土壤、水分、温度、光照等各种生态因子的不均匀分布,它普遍存在于自然环境中[6-8]。在自然状态下,异质性生境常常以斑块形式或梯度形式呈现[9]。生境异质性格局对于克隆植株相连分株源-库关系的建立往往起决定作用[10-11]。有关克隆植物对于生境异质性斑块的尺度和对比度的响应方面的研究,此前已开展较多,如一些国外学者早期的经典试验[12-15]以及国内学者近期的一些研究结果,如Qian 等[16]研究发现,野牛草(Buchloe dactyloides)在土壤异质性尺度更大的高养分斑块中的可塑性响应更显著;王九中等[17]在研究土壤异质性对克隆植物香菇草(Hydrocotyle vulgaris)的生长和竞争关系影响时,发现在异质性环境中,虽然香菇草在高养分斑块中会产生更多的根,但只有生长在高对比度生境中,才会产生更多的生物量和分株。此外,张丽丽等[18]在揭示土壤养分斑块对比度对活血丹(Glechoma longituba)克隆整合影响时,发现养分斑块对比度可以通过修饰克隆整合的强度和方向来改变其表型可塑性。然而,关于克隆植物在资源呈现梯度分布的异质生境中的响应方面的研究,迄今鲜有报道。而实际上,资源呈现梯度分布的异质生境在自然界也经常出现,比如因泥沙长期淤积而形成的河口滩涂生境即属此类,其他如光照、温度、水分、养分等生态因子呈现渐变趋势的生境也均属于此类。
生境资源梯度在总体水平上会呈现不同,在渐变顺序上或空间先后次序上也会表现出差异,而生长于其中的克隆植物势必会产生不同的生态响应。这可能涉及到不同生境位点的资源供应能力与分布于其中的克隆分株的生长需求之间是否精确匹配的问题,以及分株之间和克隆植株整体如何对此进行协调、整合以及生长收益如何等诸多复杂的生态学问题。对此进行研究,无疑在理论上和实践上均具有重要意义和价值。鉴于此,本研究选择克隆植物结缕草(Zoysia japonica)为研究对象,通过精确控制试验,探索在生境土壤资源呈现梯度变化时,其梯度的总体水平和增降次序对其生长收益的影响。
1 材料与方法
1.1 试验对象
研究对象为多年生禾本科匍匐茎型克隆植物结缕草。结缕草的基本组成单位是复合节间,是由两个缩短的节间和一个伸长的节间组成[2,19](图1)。A、B 分株或分蘖交错着生在复合节两侧,更靠近基端的称为A 分株,靠近顶端或梢端的称为B 分株。在一个复合节的A、B 分株和匍匐茎上可以分别形成各自的根系,其中A、B 分株的生长是结缕草植株向周围生境进行扩展的基础,在土壤养分供应充足时,克隆分株便进一步发育形成分枝(此为二级分枝)[2]。分枝具有和主匍匐茎相同的形态结构(即分枝也会形成复合节和分株及根系等),当分枝的复合节遇到土壤养分时,也可生根,进一步获取新的土壤资源,从而不断扩展,占领更广阔的生境。

图1 结缕草植株的典型形态结构示意图[20]Fig.1 The morphological structure of Z. japonica stolon[20]
1.2 试验设计
试验所用结缕草材料购自上海馨香绿化工程公司。选择形态较完整的结缕草克隆植株,切断匍匐茎,得到单个分株,并分别种植在塑料槽中培育。约30 d 后,从中挑选大小、根系和形态特征均匀的植株作为试验材料。把试验材料移植到直径为2. 5 cm、高15 cm 的透明塑料管中。在新形成的主匍匐茎的每个复合节的下方,设置同型号的装满河沙的透明塑料管以接住新复合节的根系,对于其他新复合节,以此类推。由主匍匐茎复合节形成的分枝则让其生长在空气中(分枝上的每个复合节下方不再设置装土的塑料管)。培养基质均为河沙。河沙过0.5 mm 筛后,多次漂洗至水完全清澈,烘箱中高温灭菌24 h 后自然干燥备用。在每个处理中,每个塑料管内填充等量河沙,并施以指定浓度的营养液,即每个克隆植株的分株所生长的生境是由一系列内装河沙并施以指定浓度营养液的塑料管组成的。
试验所用营养液为Hoagland 溶液。生境养分梯度设置有高、低养分水平和高低水平贯通3 个梯度系列。每个资源梯度水平内,沿着结缕草克隆植株的分株生长发育顺序均分别设有增长和降低两种次序生境。用Hoagland溶液稀释倍数的不同模拟生境资源变化的梯度,随着稀释倍数越高,养分浓度越低;反之,养分浓度越高。在高养分水平梯度系列中,增长次序生境为沿着结缕草生长方向施加营养液为Hoagland 溶液稀释33 倍→3 倍,降低次序生境为Hoagland 溶液稀释3 倍→33 倍;低养分水平梯度和高低水平贯通梯度系列的处理与上述类似,具体方法见表1。高养分水平、高低水平贯通和低养分水平各梯度总的养分含量比值约为2.1∶ 1.2∶ 1,其中高低水平贯通梯度的总养分介于高、低养分水平之间。3 个梯度水平不同次序处理共6 个处理,每个处理重复3 次,总共18 组结缕草克隆植株。为精确控制植株生长条件,试验在智能光照培养箱内进行。培养箱的白天设定为12 h/28 ℃、全光照,黑夜设定为12 h/20 ℃、零光照。

表1 试验中资源梯度次序处理方法Table 1 The treatment methods for the resource gradient orders in the experiment
每隔1 d,对生长于塑料管中的各复合节施以3 mL 指定浓度的等量营养液,不施加营养液时,适量浇水,即按天交替施加指定稀释倍数的营养液或水。试验过程中精心管理确保植株能够正常生长。每新长一个复合节,就下置一个新的塑料管并记录时间。当每个试验植株的主匍匐茎长到30 +1(母株)个复合节时收获。整个试验过程进行了5 个月左右。收获时,尽量不破坏根系以及根系与地上部分间的连接关系。用蒸馏水对根部进行彻底清洗,用滤纸将多余水分吸干。收获后的植株放入烘箱,在105 ℃条件下杀青30 min,在80 ℃条件下烘干至恒重,测量总生物量和各部分生物量。
1.3 数据分析
采用SPSS 20.0 对所测数据进行统计分析,用平均值和标准误表示各数值,用Duncan 法对各测定数据进行多重比较,对结缕草各器官生物量及其分配进行双因素方差分析;采用Excel 2013 作图。
2 结果与分析
2.1 生境养分梯度水平和次序对结缕草A、B 分株数量和A、B 分株总生物量的影响

图2 3 个养分梯度水平和增长、降低次序生境内结缕草的分株总数量和分株总生物量Fig.2 The total number of ramets A,the total number of ramets B,the total biomass of ramets A,the total biomass of ramets B of Z. japonica in three nutrient levels and increasing,decreasing order habitats
生长于高、低水平的降低次序生境内的结缕草植株,其A 分株数均显著多于增长次序生境(P <0.05),但在高低水平贯通的增长和降低次序生境内,A 分株数没有显著差异(图2)。生长于高水平梯度的结缕草植株B 分株数量多于低水平梯度,也多于高低水平贯通梯度。生长于高水平和低水平梯度的增长次序生境内的B 分株数都少于同一梯度的降低次序生境内的B 分株数,而生长于高低水平贯通梯度的增长次序生境内的B 分株数则多于同一梯度降低次序生境内的B 分株数(图2)。
在各个处理生境内的的结缕草A、B 分株总生物量和A、B 分株数的变化趋势并非完全一致。尽管在高低水平贯通梯度生境内的增长次序生境内的B 分株数多于同一梯度的降低次序生境内的B 分株数,但前者的B 分株生物量却小于后者。A 分株和B 分株的总生物量均表现为在高水平梯度生境>低水平梯度生境>高低水平贯通梯度生境,且A 分株和B 分株的总生物量在各个梯度的降低次序生境内都要大于同一梯度的增长次序生境,其中,A 分株和B 分株总生物量在高水平梯度的降低次序生境内显著高于其他处理组。各处理生境内的A 分株数量平均值为31 个,是B 分株数量平均值14 个的2.2 倍,且A 分株生物量平均值为1.67 g,是B 分株生物量平均值0.29 g 的5.7 倍,可见,A 分株的长势明显优于B 分株。
2.2 生境养分梯度水平和次序对结缕草A 分枝数、A 分枝长度、A 分枝总生物量的影响
在本试验的各个处理生境内,着生于结缕草各个复合节上的B 分株都未能发育成B 分枝,而部分复合节上的A 分株长成了A 分枝,因此,这里仅对生境营养梯度水平和次序对结缕草A 分枝数、A 分枝长度、A 分枝总生物量的影响进行分析。各水平梯度的增长次序生境内的A 分枝数多于同一梯度水平的降低次序生境(图3)。A 分枝总长度在高、低养分水平梯度的降低次序生境内均大于同一梯度的增长次序生境,而在高低水平贯通梯度生境内的趋势则相反,A 分枝总长度在该梯度的增长次序生境内显著大于该梯度的降低次序生境内(P <0.05)(图3)。A 分枝的总生物量在高养分水平梯度的增长次序生境内和高低水平贯通梯度的增长次序生境内,均大于同一梯度的降低次序生境,而A 分枝的总生物量在低水平梯度的降低次序生境内大于同一梯度的增长次序生境(图3)。总体而言,在各个处理生境内的结缕草A分枝总数量、总长度和总生物量的变化趋势并不完全一致,各种梯度水平和次序生境处理间的A分枝各参数差异较显著,高低水平贯通梯度的增长次序生境明显有利于A 分枝生长,在数量、总长度、总生物量上都大于同一梯度降低次序生境。

图3 3 个养分梯度水平和增长、降低次序生境内的结缕草植株的A 分枝的总数量、总长度和总生物量Fig.3 The total number of branches A,the total length of branches A,the total biomass of branches A of Z. japonica in three nutrient levels and increasing,decreasing order habitats
2.3 生境养分梯度水平和次序对结缕草的主匍匐茎总长度和匍匐茎总生物量的影响
结缕草主匍匐茎的总长度在高养分水平梯度、低养分水平梯度和高低水平贯通梯度生境内依次降低,且在高养分水平梯度的增长次序生境内最大(图4),并与其他处理生境间存在显著差异(P <0.05)。主匍匐茎总长度在各水平梯度的增长次序生境内均大于同一梯度的降低次序生境,但只在高水平梯度生境内差异显著,而在低养分水平梯度和高低水平贯通梯度生境内差异并不显著(P >0.05)。
结缕草主匍匐茎的生物量在各养分水平梯度的增长次序生境内依次降低,而在同一梯度的降低次序生境内则先降低后增加。主匍匐茎的生物量在高养分和低养分水平梯度的增长次序生境均大于同一梯度的降低次序生境,而在高低水平贯通梯度的降低次序生境显著大于同一梯度的增长次序生境(P <0.05)。

图4 3 个养分梯度水平和增长、降低次序生境内的结缕草植株主匍匐茎总长度和主匍匐茎总生物量Fig.4 The total length of stolons,the total biomass of stolons of Z. japonica in three nutrient levels and increasing,decreasing order habitats
2.4 生境养分梯度水平和次序对结缕草的总根数量、总根生物量的影响
结缕草植株的总根数量在高水平梯度、高低水平贯通梯度、低水平梯度生境内依次降低,且在降低次序生境内的总根数量均大于同一梯度的增长次序生境(图5)。结缕草根的总生物量和总根数量的变化趋势并不完全一致。结缕草的总根生物量在高水平梯度、低水平梯度、高低水平贯通梯度生境内依次降低,且在同一梯度的增长次序生境大于同一梯度的降低次序生境,但只在高水平梯度生境内差异显著(P <0.05),而在低水平梯度和高低水平贯通梯度的两种次序生境内总根生物量差异不显著(P >0.05)(图5)。
2.5 生境养分梯度水平和次序对结缕草的总生物量、根冠比的影响
结缕草植株总生物量在高养分水平梯度生境明显大于低养分水平梯度和高低水平贯通梯度生境。低养分水平梯度生境内的总生物量虽大于高低水平贯通梯度生境内的总生物量,但差异不显著(P >0.05)。各个梯度的增长次序生境内的总生物量均小于同一梯度的降低次序生境内的总生物量(图6)。
根冠比在低养分水平梯度生境内均大于高养分水平梯度和高低水平贯通梯度生境(图6)。在高低水平贯通梯度生境内,根冠比在增长次序生境显著大于降低次序生境(P <0.05)。在高养分水平梯度生境内根冠比最小,且根冠比在该梯度的增长次序生境也显著大于降低次序生境。总体而言,在各处理间,根冠比均表现差异:根冠比在低养分水平梯度的增长次序生境显著大于其他各处理生境(P <0.05),而根冠比在高养分水平梯度的降低次序生境明显小于其他各处理;根冠比在高养分水平梯度的增长次序生境与低养分水平梯度的降低次序生境、高低水平贯通梯度的降低次序生境间差异不显著(P >0.05)。
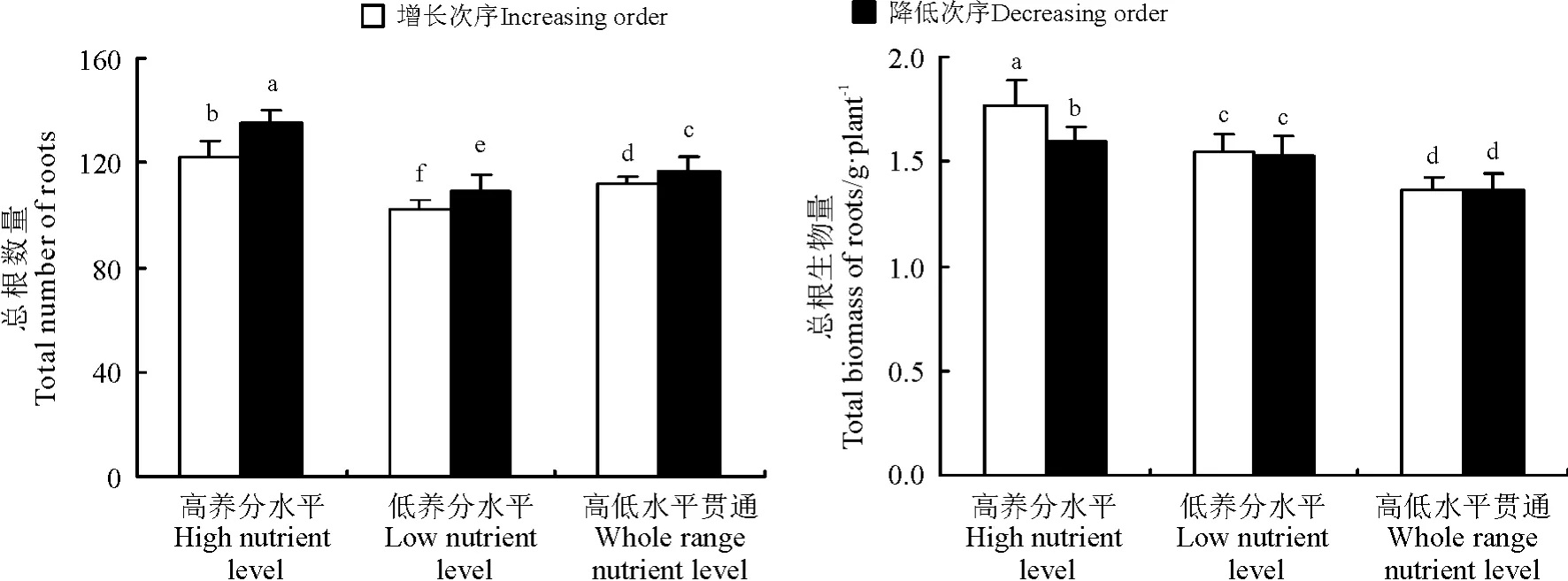
图5 3 个养分梯度水平和增长、降低次序生境内的结缕草植株总根数量和总根生物量Fig.5 The total number of roots,the total biomass of roots of Z. japonica in three nutrient levels and increasing,decreasing order habitats
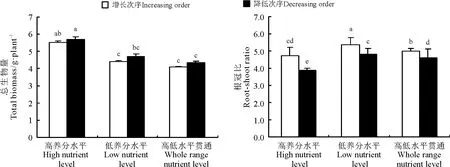
图6 3 个养分梯度水平和增长、降低次序生境内的结缕草植株总生物量和根冠比Fig.6 Total biomass,the root-shoot ratio of Z. japonica in three nutrient levels and increasing,decreasing order habitats
2.6 生境养分梯度水平和次序对结缕草植株的生物量分配的影响
图7 所示为不同养分水平梯度和次序对结缕草植株的生物量分配的影响,其中A 分株生物量包括所有的A 分株和A 分枝。在所有处理中,结缕草植株A 分株的生物量分配呈现出高养分水平梯度生境>低养分水平梯度生境>高低水平贯通梯度生境的趋势,且A 分株生物量分配在高养分和低养分水平梯度的降低次序生境大于同一梯度的增长次序生境,而在高低水平贯通梯度生境的增长次序生境略微大于降低次序(P >0.05)。B 分株生物量分配在高养分水平梯度的降低次序生境内显著(P <0.05)高于其他各处理生境,其他各处理生境的B 分株生物量分配在4.52% ~6.00%,且相互间差异不显著(P >0.05)。主匍匐茎的生物量分配在高养分水平梯度、低养分水平梯度、高低水平贯通梯度生境内依次升高,且在同一梯度的降低次序生境内升高幅度大于同一梯度的增长次序生境。在高低水平贯通梯度的降低次序生境中,主匍匐茎生物量分配最高;在高养分水平梯度的降低次序生境中,主匍匐茎生物量分配显著低于其他各处理生境。增长次序生境和降低次序生境内的根生物量分配,均分别在低养分水平梯度最大、高低水平贯通梯度居中、高养分水平梯度最小,且增长次序生境内的根生物量分配均大于同一梯度的降低次序生境。
资源梯度水平和次序对结缕草植株的A 分株生物量、B 分株生物量、匍匐茎生物量、根系生物量和总生物量都分别具有极显著影响(P <0.01);其中资源梯度水平和次序的交互作用对A 分株生物量、B 分株生物量、匍匐茎生物量、根系生物量亦有极显著影响,对总生物量影响显著(P <0.05)(表2)。资源梯度水平的不同对结缕草植株的A 分株、匍匐茎、根系的分配比有极显著影响,对B 分株的生物量分配影响不显著;不同的资源梯度次序对A分株、根系的分配有极显著影响,对B 分株生物量分配有显著影响,对匍匐茎生物量分配影响不显著;资源梯度水平和次序的交互作用对A 分株、B 分株、匍匐茎、根系的生物量分配影响均不显著。整体看来,相比于资源梯度次序,资源梯度水平对各参数的影响更大。
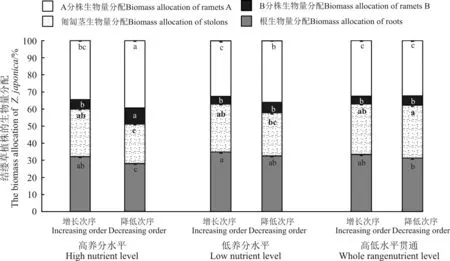
图7 3 个养分梯度水平和增长、降低次序生境内结缕草植株的生物量分配Fig.7 The biomass allocation of Z. japonica in three nutrient levels and increasing,decreasing order habitats
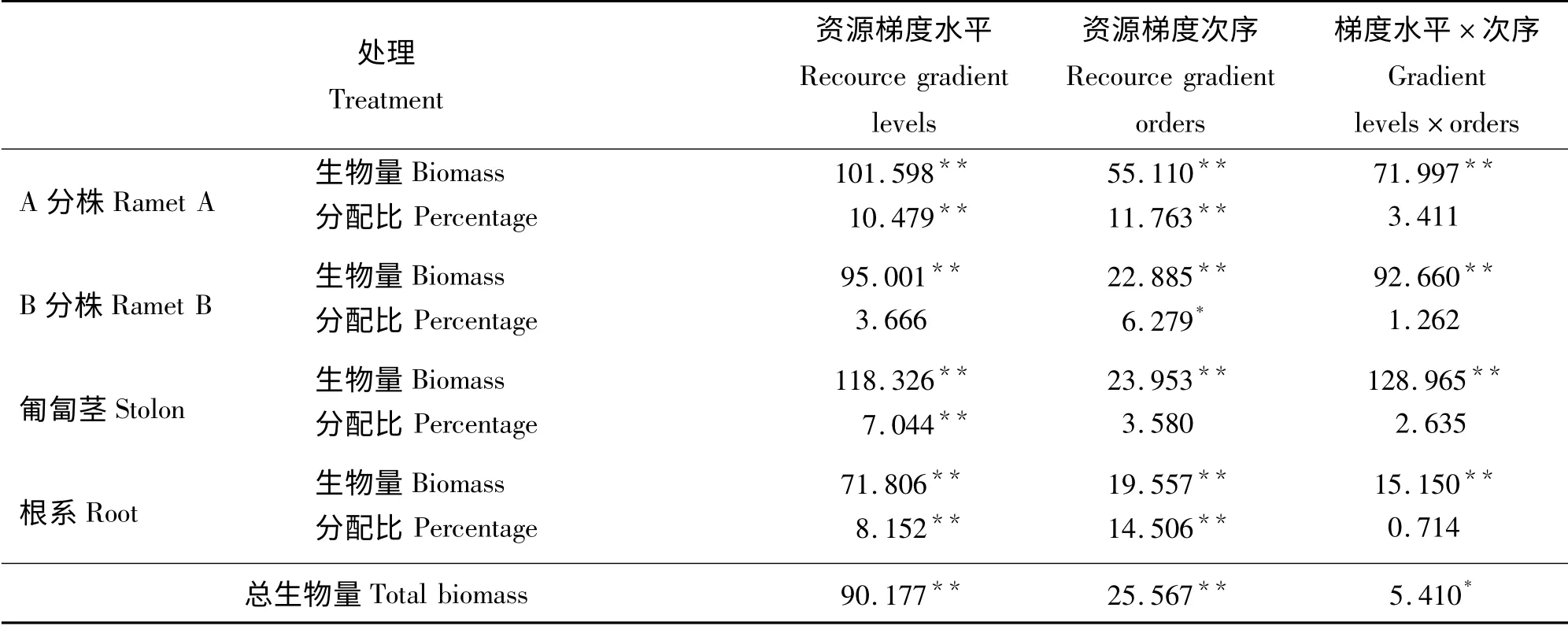
表2 资源梯度水平和次序对结缕草各器官生物量及其分配影响的F 检验Table 2 F significance test of effect of recource gradient levels and orders on biomass of ramet A,ramet B,stolon,root and their allocation of Z.japonica
3 讨论
氮和磷都是植物生长所需的主要矿质元素,也是陆地生态系统内植物生长的主要影响元素[21]。有研究表明,一定程度的氮、磷增加会引起植物总生物量及营养器官生物量的增加,进而促进植物生长[22]。本研究通过精确控制试验,探索了在生境土壤资源呈现梯度变化时,其梯度的总体水平和增降次序对结缕草生长收益的影响。生物量及其分配格局是植物表型差异的综合反映[23],通过F 检验发现资源梯度的总体水平和增降次序对各部分器官的生物量有显著影响,分配格局亦互有不同,这些结果都表现了结缕草在响应不同梯度水平和次序时较强的可塑性。本研究表明,结缕草植株的总生物量呈现出高养分水平梯度生境>低养分水平梯度生境>高低水平贯通梯度生境的趋势,且降低次序生境的结缕草植株总生物量均高于增长次序生境。虽然各养分水平梯度内的两种次序生境的总体养分相同,但结缕草的生长收益却有明显差异。在降低次序生境内,母株所得到的土壤养分供应最多,而在整个生长过程中,母株生长发育时间最长,对土壤养分的需求也最高,母株以后的其他分株的生长发育时间依次减少,他们对于土壤养分的需求也相应依次减少,而与此同时,母株以后的其他分株所得到的土壤养分供应也相应减少,因此,降低次序生境的土壤养分供应的递减趋势与克隆植株上各分株依次生长需求的递减趋势基本协调,所以,结缕草克隆植株的总生长收益也更大。与此对应的增长次序生境的土壤养分供应递增的趋势与克隆植株各分株的生长需求递减趋势不相协调,因此,生境土壤资源未能被生长于其中的结缕草克隆植株充分有效地利用,总生长收益也较小。Li 等[24]在研究生境土壤氮素资源异质性格局对结缕草生长的影响时发现,当结缕草克隆植株较早地从肥沃土壤斑块中获取资源时,其总生物量较大,这与本研究结果的趋势相一致。
在本研究的所有处理生境中,结缕草植株上所有复合节上生长的B 分株都未能发育成B 分枝,而部分复合节上的A 分株则发育成为了A 分枝,可见,在养分一定的情况下,结缕草植株优先选择将资源投资给A 分株,从而促进了A 分株的生长发育。在每个养分水平梯度的增长或降低次序生境内,结缕草克隆植株的A 分株数量和总生物量均显著大于B 分株数量和总生物量,而且在高养分水平梯度生境内,A 分株的表现更优。这可能是因为A 分株更靠近基端,而B 分株更靠近顶端,顶端优势促使B分株的生长受抑制。另外,在各个养分水平梯度的降低次序生境都比增长次序生境更能显著促进A分株和B 分株的生长。然而,结缕草的分枝强度变化趋势和A 分株并不一致,表现为在各梯度水平的增长次序生境比降低次序生境更能促进A 分枝数量的增加,尤其是在高低水平贯通生境中差异更为显著。与低养分水平梯度生境相比,高低水平贯通生境的养分梯度更大(Hoagland 营养液浓度从稀释4 倍到稀释64 倍),这意味着分株所处小生境的土壤养分相差大或生境斑块对比度大,在此情况下,克隆植株内分株间发生克隆整合的可能性也越大[25-26]。然而本研究结果显示,虽然高低水平贯通梯度生境的整体养分高于低养分水平梯度生境,但在高低水平贯通梯度的降低次序生境中的A 分枝的生长反而小于低养分水平梯度的增长及降低次序生境;只有在高低水平贯通梯度的增长次序生境中,A 分枝的生长才显著优于低养分水平梯度的增长及降低次序生境。分析原因,可能较为复杂,这需要首先理解结缕草克隆植株生理整合的基本过程:结缕草克隆植株的资源(如氮素[27]、碳素[28]、水分[29]等)整合方向以向顶端传输为主,克隆分株所吸收的资源或合成的物质首先满足自身生长发育的基本需求,然后将多余的一部分向其他分株进行运输和分享,如果尚有多余的其他部分资源或物质,无法被及时传输出去,则被分株自身用于形成分枝结构,进行侧向生长。在本研究中的高低水平贯通梯度的增长次序生境中,系列分株所处小生境的土壤养分逐渐增加且增幅较大。位于克隆植株基部的一些分株,因所处小生境土壤养分较低而难以吸收到充足的养分,因而只是简单地维持自身生长,很少形成A分枝,但随着后续分株所在小生境土壤养分水平逐渐大幅提高,一些分株开始吸收到丰富的土壤养分,在满足自身生长需求之后,还与相邻分株分享资源,并且可能还有富余的没被运出的资源,这部分资源被用于A 分株进一步发育形成A 分枝,由于后发育形成的分株所处小生境的土壤养分逐渐增加,因此,克隆植株上后发育形成的复合节上的分株更易于形成A 分枝(未发表结果),这导致在高低水平贯通梯度的增长次序生境中A 分枝的数量、长度及生物量显著高于低养分水平的增长及降低次序生境。而在本研究中的高低水平贯通梯度的降低次序生境中,系列分株所处小生境的土壤养分逐渐降低且降幅较大。位于克隆植株基部的若干分株,因小生境养分供应充足,而促进了A 分枝的生长,而其余大部分分株,则因小生境养分供应的快速降低,分株所吸收的养分不够丰富,而难以进一步发育形成A 分枝,因此,在高低水平贯通梯度的降低次序生境中A 分枝的数量、长度及生物量显著低于同一养分梯度水平的增长次序生境及其他养分梯度水平的两类次序生境。
在各处理中,高养分水平梯度的两类生境较其他两个水平梯度的各生境更能促进主匍匐茎的伸长生长,且在高养分水平梯度的增长次序生境内,主匍匐茎总长度最长、生物量也更高。主匍匐茎长度在各养分水平梯度的增长次序生境也高于同一梯度的降低次序生境。匍匐茎是结缕草进行水平空间移动的最主要器官,也是分株间进行生理整合的通道。在各水平梯度的增长次序生境中,随着系列分株所处土壤小生境中资源水平的逐渐增加,结缕草相对增加了对匍匐茎伸长生长的投资,这有利于结缕草更有效、更快捷地进行水平空间的移动和探索新的生境资源。在高低水平贯通梯度的降低次序生境内,结缕草植株的主匍匐茎虽然比增长次序生境短,但生物量反而比增长次序生境大。分析原因可能是因为在该梯度的降低次序生境内主匍匐茎较粗,单位长度的匍匐茎生物量更高,所以趋势与其他水平梯度不一致(在高养分水平和低养分水平梯度中,主匍匐茎的生物量在增长次序生境内高于同一梯度的降低次序生境)。
虽然在高养分水平梯度生境中,结缕草植株的主匍匐茎总长度最长且总生物量最高,但就生物量分配比而言,结缕草克隆植株对于匍匐茎的生物量投资比例在高低水平贯通梯度生境(相当于中等养分水平)条件下最高,而在高养分或低养分条件下,该比例都倾向于减少。F 检验亦显示,资源梯度水平对匍匐茎的生物量分配的确有显著影响。在de Kroon 和Schieving[30]此前构建的克隆植物在资源异质生境条件下的生物量分配模型中,也预测克隆植物在中等资源水平时对间隔子的生物量分配最高。这一点与本研究结果不谋而合。此外,张淑敏等[31]的研究结果同样显示在中等养分条件下,绢毛匍匐委陵菜(Potentilla reptans var. sericophylla)对匍匐茎的生物量投资倾向最大。
根系作为植物从土壤中获取养分和水分的主要营养器官,其发达程度决定了植株吸收能力的大小[32],具有较大的可塑性[33-34]。在本研究中的高养分水平梯度的增、降次序生境内,结缕草根系数量和生物量均显著增加,但根系数量和根生物量变化趋势不一致,比如与低养分水平梯度生境相比,总养分水平处于中等的高低水平贯通梯度生境内的根系数量多而根生物量小,分析原因可能是因为其新生根较多,而营养累积时间较短,因此,虽在根数量上占优势,但根生物量并不占优势。
分析植物体的地上和地下生物量分配权衡或根冠比是衡量整个植株在特定生境内的生长策略的常用方法[35-36]。本研究发现,当氮磷共同增加(在高养分水平梯度生境内)时,结缕草克隆植株的根冠比降低。这可能是因为当根系所在生境养分充足时,分株会分配更多的生物量到地上部分以增加叶片进行光合作用的能力,进而促进植物的生长[37]。而在低养分水平梯度生境条件下,结缕草克隆植株的根冠比趋向增高。可见,土壤养分贫瘠时,结缕草通过增加根系的生物量以期获取更多的土壤资源,并增加竞争力。克隆植物劳动分工的典型模式是趋富专化,即分株将相对多的生物量投向吸收较丰富资源的器官或部分[1],以增强分株对丰富资源的吸收能力。而本研究结果显示,结缕草分株在低养分水平生境内,对根部的投资加大,即出现趋贫专化。这一结果与Li 等[20]的结果有相似之处。这一现象可能意味着,结缕草克隆分株具有较强的独立性,即分株仅在自身所处的小生境内发生可塑性反应,而受相邻分株的影响不大[37-38],另外,出现这一情况也可能与试验中所设置的养分水平有关,即斑块之间对比度没有达到诱发产生劳动分工的阈值[20,39]。
4 结论
1)在不同梯度水平和次序生境中,结缕草植株A 分株的生长明显优于B 分株:A 分株的总数量平均值约为B 分株的2.2 倍,而总生物量平均值约为B 分株的5.7 倍;部分复合节上的A 分株发育成了A 分枝,而所有复合节上的B 分株均未发育成B 分枝。
2)结缕草克隆植株的总生物量在高养分水平梯度生境>低养分水平梯度生境>高低水平贯通梯度生境,且在同一梯度的降低次序生境的总生物量均高于增长次序生境,表明提高生境中的养分水平能显著增加结缕草克隆整合的生长收益,且当小生境养分供应与分株的生长需求相匹配时,结缕草克隆整合的生长收益更大。
3)结缕草克隆植株的根冠比在低养分水平梯度生境>高低水平贯通梯度生境(相当于中等养分水平)>高养分水平梯度生境,表明结缕草克隆分株具有较强的独立性,分株更倾向于在自身所处小生境内发生可塑性反应,而受相邻分株影响不大。
4)结缕草克隆植株对匍匐茎的生物量投资比例在高低水平贯通梯度生境(相当于中等养分水平)中最大。
[1] 董鸣,于飞海.克隆植物生态学术语和概念[J].植物生态学报,2007,31(4):689-694.
[2] 李德志,申芳芳,王长爱,柯世朕,王超华,范旭丽,宋云,李红,周燕.克隆植物结缕草在两种环境中的生长发育特征[J].华东师范大学学报,2006(6):117-124.
[3] 葛俊,邢福.克隆植物对种间竞争的适应策略[J].植物生态学报,2012,36(6):587-596.
[4] 王长爱,李德志,朱志玲,王旭平,盛丽娟,石强.克隆植物生理整合作用的研究方法及其应用[J].应用与环境生物学报,2006(4):581-588.
[5] Wang N,Yu F H,Li P X,He W M,Liu F H,Liu J M,Dong,M.Clonal integration affects growth,photosynthetic efficiency and biomass allocation,but not the competitive ability,of the alien invasive alternanthera philoxeroides under severe stress[J].Annals of Botany,2008,101(5):671-678.
[6] Jackson R B,Caldwell M M.The scale of nutrient heterogeneity around individual plants and quantification with geostatistics[J].Ecology,1993,74:612-614.
[7] Caldwell M M,Pearcy R W.Exploitation of Environmental Heterogeneity by Plants:Ecophysiological Processes above-and below-Ground[M].San Diego:Academic Press,1994:98-106.
[8] Gross K L,Pregitzer K S,Burton A J.Spatial variation in nitrogen availability in three successional plant communities[J].Journal of Ecology,1995,83(3):357-367.
[9] Kotliar N B,Wiens J A.Multiple scales of patchiness and patch structure:A hierarchical framework for the study of heterogeneity[J].Oikos,1990,59:253-260.
[10] Pitelka L E,Ashmun J W.Physiology and integration of ramets in clonal plants[A].Jackson J B C,Buss L W,Cook R E.Population Biology and Evolution of Clonal Organisms[M].New Haven:Yale University Press,1985:399-435.
[11] Marshall C.Source-sink relations of interconnected ramets[A].van Groenendael J,de Kroon H eds.Clonal Growth in Plants:Regulation and Function[M].The Hague:SPB Academic Publishing,1990:23-41.
[12] Birch C P D,Hutchings M J.Exploitation of patchily distributed soil resources by the clonal herb Glechoma hederacea[J].Jounral of Ecology,1994,82:653-664.
[13] Wijesinghe D K,Hutchings M J.Enviromental heterogeneity and the growth of Glechoma hederacea:The effects of scale of heterogeneity on performance[J].Jounral of Ecology,1997,85:17-28.
[14] Wijesinghe D K,Handel S N. Advantages of clonal growth in heterogeneous habitats-an experiment with Potilla simplex[J].Jounral of Ecology,1994,82:495-502.
[15] Wijesinghe D K,Hutchings M J.The effects of environmental heterogeneity on the performance of Glechoma hederacea:The interactions between patch constrast and patch scale[J].Jounral of Ecology,1999,87:860-872.
[16] Qian Y Q,Luo D,Gong G,Han L,Ju G S,Sun Z Y.Effects of spatial scale of soil heterogeneity on the growth of a clonal plant producing both spreading and clumping ramets[J].Journal of Plant Growth Regulation,2014,33(2):214-221.
[17] 王九中,董必成,罗芳丽.土壤异质性对香菇草生长和竞争关系的影响[J].江苏农业科学,2014,42(6):302-306.
[18] 张丽丽,董鸣,李仁强,王艳红,崔清国,何维明.土壤养分斑块对比度改变活血丹克隆整合强度和方向[J].植物生态学报,2007,31(4):619-624.
[19] Shoji S.Ecological studies on the Zoysia type grassland:Development and growth of the stolon of Z.japonica Steud[R].Report of Institute of Agriculture Research,Tohoku University,1976,27(4):49-59.
[20] Li D Z,Takahashis,Zhu T C.Effects of soil nitrogen availability and clonal integration on the branching behaviors of Zoysia japonica[J].Acta Prataculturae Sinica,2006,15(5):135-146.
[21] 刘洋,张健,陈亚梅,陈磊,刘强.氮磷添加对巨桉幼苗生物量分配和C∶ N∶ P 化学计量特征的影响[J].植物生态学报,2013,37(10):933-941.
[22] 全国明,谢俊芳,章家恩,毛丹鹃.氮、磷养分对飞机草营养器官表型可塑性的影响[J]. 生态学杂志,2014(10):2625-2632.
[23] 谢晓玲,周蓉,谢焕松,沈振西,邓自发. 藏北高原不同海拔四川嵩草克隆种群表型和繁殖对策[J]. 草业科学,2014,31(12):2270-2276.
[24] Li D Z,Ito M,Okajima T.Effects of soil nutrient heterogeneity on the growth of plants under the various distributions and levels of nutrients in Zoysia japonica Steud[J].Grassland Science,2005,51(1):41-44.
[25] Eriksson O,Jerling L.Hierarchical selection and risk spreading in clonal plants[A]. van Groenendael J,de Kroon H. Clonal Growth in Plans:Regulation and Function[M].The Hague:SPB Academic Publishing,1990:79-94.
[26] Caraco T,Kelly C K.On the adaptive value of physiological integration in clonal plants[J].Ecology,1991,72:81-93.
[27] 李立科,李德志,赵鲁青,潘宇,吕媛媛,赵美霞,高锦瑾,王浩,程立丽,何莹莹.克隆植物结缕草主匍匐茎中氮素的传输特征[J].东北林业大学学报,2011(11):40-45.
[28] 何莹莹,李德志,徐洁,王浩,程立丽,刘微.不同光照条件下结缕草全株碳素生理整合格局及其生态效应[J].植物资源与环境学报,2013(4):1-10.
[29] 程立丽,李德志,朱志玲,何莹莹,王浩,纪倩倩,刘微,陈慧娟,赖苏雯.克隆植物结缕草的水分生理整合格局特征及其生态效应分析[J].西北植物学报,2013,33(7):1442-1451.
[30] de Kroon H,Schieving F.Resource allocation patterns as a function of clonal morphology:A general model applied to a foraging clonal plant[J].Journal of Ecology,1991,79:519-530.
[31] 张淑敏,于飞海,董鸣.土壤养分水平影响绢毛匍匐委陵菜匍匐茎生物量投资[J]. 植物生态学报,2007,31(4):652-657.
[32] Prefitzer K S,Deforest J L,Burton A J,Allen M F,Ruess R W,Hendrick R L.Fine root architecture of nine north American trees[J].Ecological Monographs,2002(2):293-309.
[33] Hutchings M J,De Kroon H.Foraging in plants:The role of morphological plasticity in resource acquisition[J].Advances in Ecological Research,1994,25:159-238.
[34] Williamson L C,Ribrioux S P C P,Fitter A H,Leyser H M O.Phosphate availability regulates root system architecture in Arabidopsis[J].Plant Physiology,2001,126(2):875-882.
[35] Hui D F,Jackson R B.Geographical and interannual variability in biomass partitioning in grassland ecosystems:A synthesis of a field data[J].New Phytologist,2006,169(1):85-93.
[36] Enquist B J,Niklas K J.Global allocation rules for patterns of biomass partitioning in seed plants[J].Science,2002,295:1517-1520.
[37] 辛小娟,王刚,杨莹博,任正炜. 氮、磷添加对亚高山草甸地上/地下生物量分配的影响[J]. 生态科学,2014,33(3):452-458.
[38] de Kroon H,Huber H,Stuefer J F,van Groenendael J M.A modular concept of phenotypic plasticity in plants[J].New Phytologist,2005,166:73-82.
[39] 李元恒,王正文,马晖玲. 生境斑块对比度对鹅绒委陵菜克隆内分工的影响[J]. 植物生态学报,2008,32(5):1166-1174.
