杉木-厚朴混交林林下物种多样性研究
2015-03-18潘世龙
潘世龙
(福建省建瓯市顺阳林业工作站,福建建瓯353100)
杉木是中国南方重要的速生造林树种之一,由于其生长快、产量高、材质好、易繁殖等特点,深受广大农民的喜爱[1]。长期以来,由于集约经营、短轮伐期、炼山等原因,杉木人工林生态系统受到了巨大的干扰,人工林连栽生产力下降日趋严重[2]。如何采用有效措施尽快恢复杉木人工林地力水平,是当前杉木人工林持续经营中亟需解决的重大问题之一。
林下植被是森林生态系统的重要组成部分,在维护地力、促进养分循环、维持森林物种多样性、保持水土等方面具有不可忽视的作用[1]。近年来由于杉木人工林生产力下降明显,有关杉木林下植被的研究逐渐得到重视[3-7]。恢复林下植被物种多样性对杉木林生态系统的重建具有重要的理论和实践意义。目前研究者主要针对杉木纯林林下植被展开研究,而关于杉木与阔叶树种混交林林下植被发育的研究较少。鉴于此,笔者以福建省建瓯市顺阳乡溪东村杉木与厚朴混交林及杉木纯林为研究对象,开展林下植被组成、物种多样性的调查研究,为提高杉木林地生态系统的生物多样性和生态稳定性提供理论依据。
1 试验地概况
试验地点位于福建省建瓯市顺阳乡溪东村(118°35′54″E,27°4′53″N),属于亚热带季风气候,山地气候明显,春季多雨,夏长较热,秋高气爽,冬短多雾,年平均气温18.7℃;冬季平均气温8.0℃,夏季月平均气温28.5℃,年平均降水量1 663.0 mm,年均蒸发量1 458.4mm,年均相对湿度是81%,无霜期286 d。地势属丘陵地带,平均海拔110 m,土壤为酸性红壤,土层平均厚度50 cm。研究对象为杉木-厚朴混交林和杉木纯林,混交林采用行状混交方式,3行杉木混交2行厚朴,株行距2 m×2 m。2种林分前茬均为杉木纯林采伐迹地,采用人工直播方式,于2001年造林,林龄均为15年。样地概况见表1。
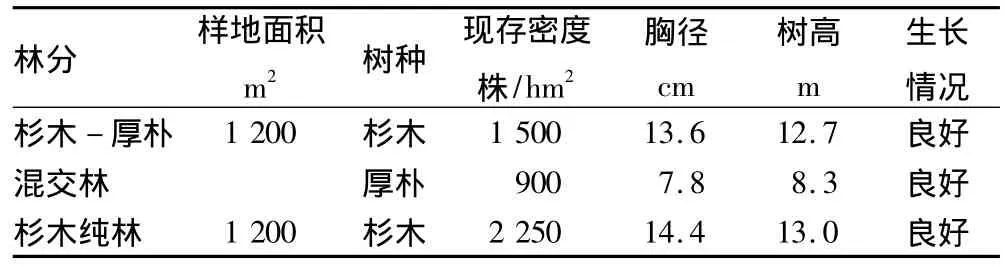
表1 样地概况
2 研究方法
2.1 样地调查 2015年5月中旬设置样地,调查取样,每种林分分别设置20 m×20 m的标准地3个,采用梅花型布点方法设置5个1 m×1 m小样方,分别记录灌木种类、株数、高度和冠幅,草本和层间植物的种类、株丛数和盖度。
2.2 重要值(IVI)及多样性指数
2.2.1 重要值[8]。
灌木重要值=相对显著度(%)+相对密度(%)+相对频度(%)
草本层和层间植物重要值=相对盖度(%)+相对密度(%)+相对频度(%)
2.2.2 物种多样性。采用物种丰富度指数(S)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(P)进行评价。计算公式如下:
物种丰富度指数R0=S Pielou均匀度指数J=H/ln S
式中,ni为第i种的个体数,N为所有种的个体数;Pi为第i种的个体数ni占所有种个体总数的比例,即Pi=ni/n;S为种i所在样地物种种类总数,即物种丰富度指数。
3 结果与分析
3.1 林下植被物种组成 由表2可知,在杉木-厚朴混交林3个大样方(1 200 m2)内共发现维管束植物35种,其中灌木14种,占40.00%,分属9科;草本16种,占45.71%,分属14科;层间植物5种,占14.29%,分属5科。杉木纯林共发现维管束植物21种,其中灌木8种,占38.10%,分属7科;草本10种,占47.62%,分属9科;层间植物3种,占14.29%,分属3科。
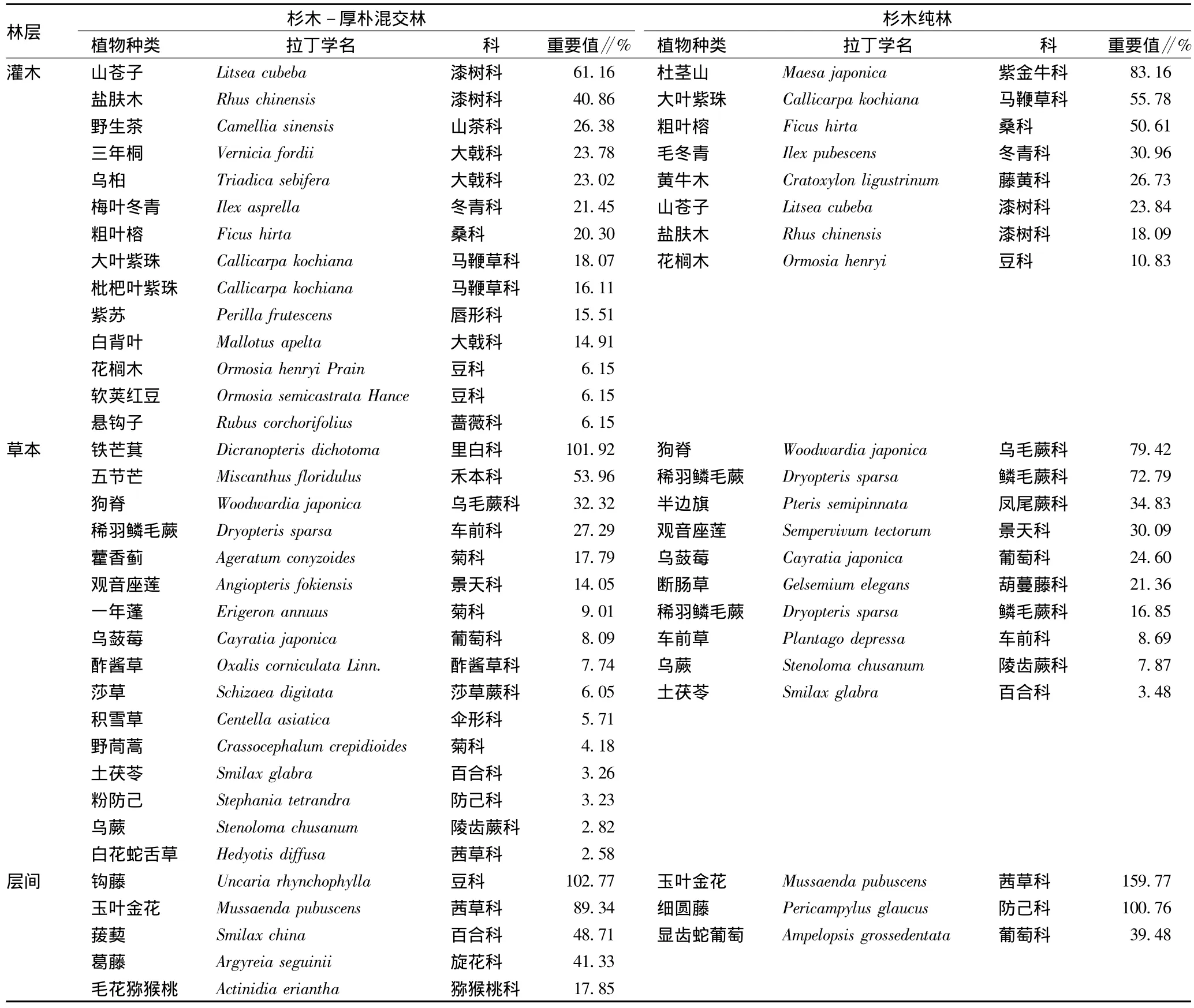
表2 杉木-厚朴混交林与杉木纯林林下物种组成及重要值比较
3.2 林下植被物种重要值
3.2.1 灌木层。重要值(IVI)是以综合值来表示群落中不同种群的相对重要性和对群落的适应程度,是研究群落的重要指标之一。由表2可知,杉木纯林中耐阴的优势树种较多,如灌木层中喜阴植物杜茎山重要值排在第1位,其次为大叶紫珠、粗叶榕和毛冬青等,这些物种在荫蔽环境分布广泛。而喜光植物山苍子的重要值在纯林中排在第6位,但在杉木-厚朴混交林中重要值居第1位,阳生物种盐肤木的重要值也从杉木纯林中第7位上升到第2位。盐肤木属于中生性偏阳的物种,可以在郁闭度不高或冠层较稀疏、透光度改善的林分中生长,却不能在郁闭度较高的林分中生长。混交林中,阴生物种大叶紫珠和粗叶榕重要值分别从杉木纯林中第2位、第3位降到第8位和第7位。混交林比纯林新增加了野生茶、三年桐、乌桕、梅叶冬青等9种灌木。
3.2.2 草本层。杉木纯林草本层中的优势物种主要有狗脊、稀羽鳞毛蕨、半边旗、观音座莲、乌蔹莓等喜阴植物,而在杉木-厚朴混交林草本层中排在前2位的为铁芒萁和五节芒。铁芒萁是一个喜阳性的蕨类植物,在荫蔽的林下较少见到。而对荫蔽环境有着较好适应特性的狗脊、稀羽鳞毛蕨、乌蔹莓和乌蕨等草本植物在杉木-厚朴混交林中的重要地位明显下降,其中狗脊降到第3位、稀羽鳞毛蕨降到第4位、乌蔹莓降到第8位、观音座莲降到第6位、乌蕨降到第15位。在混交林中又出现了铁芒萁、五节芒、藿香蓟、一年蓬和乌蔹莓等12种草本植物。因此,与杉木-厚朴混交林相比,杉木人工纯林中林分郁闭度较高,透光率较低,其中草本优势种多为一些耐阴植物,并且几乎都是蕨类植物和禾本科植物,双子叶植物几乎没有进入优势种的范围。
3.2.3 层间。2种林分中层间植物种类较少,杉木纯林中仅发现玉叶金花、细圆藤和显齿蛇葡萄,杉木-厚朴混交林中又出现了钩藤、菝葜、葛藤和毛花猕猴桃植物,其中葛藤属于阳生物种,随郁闭度的增加其数量急速下降,直到乔木冠层逐渐稀疏、透光度改善的林分中才会爆发。喜阴的玉叶金花重要值在混交林中也由杉木纯林中首位降到第2位。
3.3 林下植被物种多样性 由表3可知,林下灌木、草本和层间植物的丰富度指数(S)、多样性指数(D和H)均表现为杉木-厚朴混交林明显高于杉木纯林。灌木层与层间植物的均匀度指数也是混交林高于纯林,但草本层的均匀度指数却表现为纯林高于混交林。林下植被不同层次间,草本层丰富度指数(S)>灌木层丰富度指数(S)>层间植物丰富度指数(S),除杉木-厚朴混交林灌木层均匀度指数(J)>层间植物均匀度指数(J)>草本层均匀度指数(J)外,2种林分多样性指数(D和H)和均匀度指数(J)在各林层从大到小依次表现为灌木层、草本层、层间植物。其中灌木层D指数变化范围为0.903 8 ~0.942 5,草本层 D 为 0.797 2 ~0.822 8,层间植物D为0.492 6~0.693 1;灌木层 H指数变化范围为2.900 5 ~3.776 4,草本层H 为2.581 6 ~2.802 0,层间植物H为1.141 2~1.936 0;灌木层均匀度指数 J变化范围为0.890 7 ~0.993 3,草本层 J为0.701 9 ~0.774 2,层间植物 J为0.717 2 ~0.820 3。
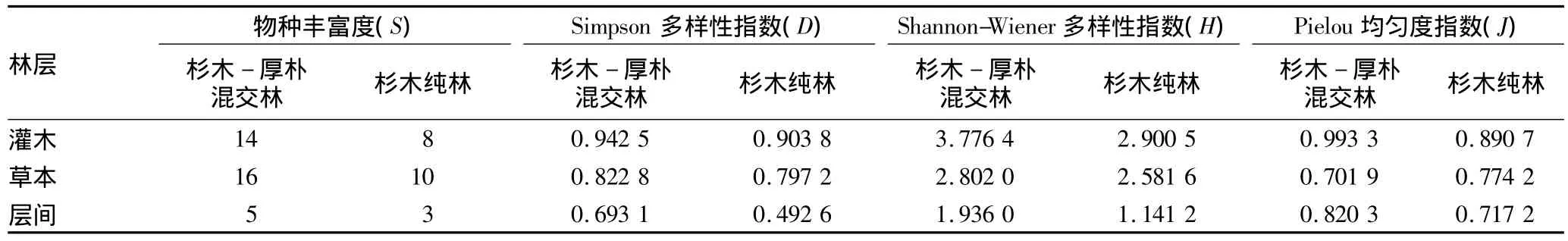
表3 杉木-厚朴混交林与杉木纯林物种多样性比较
4 讨论
根据群落演替理论,不同物种对光照、土壤等资源环境的竞争利用导致其在变化环境中处于不同的地位[9]。杉木自身的生物学特性导致杉木枝叶会宿存于树干长达4~5年之久,林分郁闭度增加,林下植物多以耐阴性物种为主,直到杉木15年生后才大量脱落,林分开始逐渐疏开[10]。研究发现,杉木-厚朴混交林与杉木纯林物种丰富度差异较大,分别为35种和21种,其优势物种的生态学特性差异明显。杉木人工纯林(15年)林下植被以阴生物种杜茎山、大叶紫珠、粗叶榕、狗脊、稀羽鳞毛蕨、玉叶金花和细圆藤等占绝对优势,杉木-厚朴混交林林下阳生物种山苍子、盐肤木、铁芒萁、五节芒和钩藤优势地位明显上升。说明杉木与厚朴混交林林分改善了内部的垂直结构,林冠层逐渐疏开,促进了阳生物种的定居。
物种的多样性水平反映了生物群落在组成、结构、功能方面的异质性,体现了群落结构在发展过程中的稳定程度和生境的差异[11]。研究发现,杉木-厚朴混交林的多样性指数均高于杉木纯林,灌木层与层间植物的均匀度指数也表现为混交林的较高。林下物种多样性水平与林冠层的郁闭度关系密切,杉木-厚朴混交林林分垂直的立体结构,有利于林下植被灌、草和层间植物的形成,促进林下植被物种多样性的提高,该研究结果与其他人的结论相似[3,12]。曾斌[12]研究发现,杉木人工林的多样性指数与均匀度指数均表现为中龄林最高,而过熟林与幼龄林较低,说明杉木幼龄林较高的郁闭度导致林下物种多样性较低,伴随杉木林分的自然稀疏和人工整枝,林分郁闭度下降,透光率增加,物种多样性也开始丰富起来。
5 结论
(1)在每种调查林分内(1 200m2),杉木-厚朴混交林林下物种丰富度为35种,明显高于杉木人工纯林21种,林下植被组成中草本层物种所占比例最大(45.71% ~47.62%)。杉木纯林郁度高,林分透光率差,林下植被物种单一,阴生物种较多,如杜茎山、大叶紫珠、粗叶榕、毛冬青、狗脊、稀羽鳞毛蕨、玉叶金花和细圆藤等。与杉木纯林相比,杉木-厚朴混交林改变了林分的空间结构,随着林冠层疏开,林分郁闭度下降,林内透光率增加,使林下植被出现了较明显的3层结构,主要灌木物种有山苍子、盐肤木、野生茶和三年桐等,草本物种有铁芒萁、五节芒、狗脊和稀羽鳞毛蕨等,层间植物有钩藤、玉叶金花和菝葜等。其中山苍子、盐肤木、铁芒萁和钩藤等阳生物种在混交林分中出现并占有重要地位。
(2)2种林分Simpson多样性指数(D)和Shannon-Wiener多样性指数(H)在各林层中从大到小均依次表现为灌木层、草本层、层间植物,杉木-厚朴混交林林下植被各层次的多样性指数均大于杉木纯林,混交林的灌木和层间植物Pielou均匀度指数(J)大于杉木纯林,但草本层却表现为杉木纯林较大。因此,杉木-厚朴混交林构建了复合层次的林分结构,使林分内透光率增加,使林下物种多样性得以提高,对于恢复杉木退化地物种多样性和生态功能具有重要促进作用。
[1]杨承栋,屠南星,姚茂和,等.杉木林下植被对改良土壤理化、生物特性的效用[M]//盛炜彤.人工林地力衰退研究.北京:中国科技出版社,1992:276-283.
[2]郭琦,王新杰.不同混交模式杉木人工林林下植被生物量与土壤物理性质研究[J].中南林业科技大学学报,2014(5):70-74.
[3]周德明,王宗永.南方不同林龄杉木人工林林下物种多样性比较[J].林业资源管理,2010(6):65-70.
[4]范少辉,马祥庆,傅瑞树,等.不同栽植代数杉木林林下植被发育的比较研究[J].林业科学研究,2001,14(1):8-16.
[5]林开敏,黄宝龙.杉木人工林林下植物物种β多样性的研究[J].生物多样性,2001,9(2):157-161.
[6]林开敏,俞新妥,黄宝龙,等.杉木人工林林下植物物种多样性的动态特征[J].应用与环境生物学报,2001,7(1):13-19.
[7]杨承栋.发育林下植被是恢复杉木人工林地力的重要途径[J].林业科学,1995,31(3):275-283.
[8]游水生,张志翔,李如泽,等.福建武平帽布米槠林火烧后植物种类变化的研究Ⅱ.火烧前后重要值和物种多样性变化[J].福建林学院学报,1998,18(1):20-23.
[9]杨小波.植物群落演替的生理生态机理研究的现状和展望[J].海南大学学报(自然科学版),1997,15(2):147-152.
[10]张家城,盛伟彤.杉木人工林树上宿存枯死枝、叶在冠层与枯枝叶层分解的比较研究[J].林业科学,2001,37(6):7-11.
[11]杨立学,孙跃志.不同林龄胡桃楸林下植物多样性的差异[J].生态学杂志,2013,32(4):807-812.
[12]曾斌,刘瑞敏,翟学昌,等.不同林龄杉木人工林物种多样性研究[J].安徽农业科学,2010,38(9):4877-4879.
