水分条件变化对哀牢山亚热带常绿阔叶林林下幼苗死亡率的影响
2015-03-13杞金华章永江张一平刘玉洪鲁志云武传胜温韩东
杞金华, 章永江, 张一平, 刘玉洪, 鲁志云, 武传胜, 温韩东
1 中国科学院西双版纳热带植物园, 勐仑 666303 2 中国科学院哀牢山亚热带森林生态系统研究站, 景东 676200 3 哈佛大学有机和进化生物学系, 美国剑桥 02138
水分条件变化对哀牢山亚热带常绿阔叶林林下幼苗死亡率的影响
杞金华1,2, 章永江1,3,*, 张一平1,2, 刘玉洪1,2, 鲁志云1,2, 武传胜1,2, 温韩东1,2
1 中国科学院西双版纳热带植物园, 勐仑 666303 2 中国科学院哀牢山亚热带森林生态系统研究站, 景东 676200 3 哈佛大学有机和进化生物学系, 美国剑桥 02138
为探讨环境水分条件变化对亚热带森林林下乔木幼苗动态的影响,以及木材密度和幼苗在干旱中死亡率的关系,研究了哀牢山常绿阔叶林2005—2011年的总体和主要树种幼苗(黄心树、多果新木姜子、多花山矾、鸭公树和大花八角)的死亡率,并分析了幼苗死亡率和旱季幼苗根系所在土层土壤质量含水量、旱季降水量、旱季降水日数以及木材密度的关系。结果表明:1)幼苗死亡率与旱季降水日数、旱季浅层土壤平均质量含水量有显著的负相关关系,和旱季降水量无显著相关性;2)总幼苗及5种乔木幼苗的死亡率均在2010年(西南干旱)达到有观测以来最高,是2009年的2—10倍,其中多花山矾、黄心树幼苗的死亡率最高,大花八角幼苗的死亡率最低;3)在种间,幼苗2010年干旱中的死亡率和木材密度显著正相关,即木材密度较大的物种幼苗死亡率更高。研究表明林下幼苗由于根系较浅,对降雨变化较为敏感,因而受到了这次干旱的较大影响。由于木材密度较低的树种在干旱中有较低的死亡率,干旱频度和强度的增加可能使低木材密度幼苗的丰富度增加,森林的组成也将受到影响。
干旱; 幼苗动态; 土壤质量含水量; 降水; 木材密度
幼苗在森林中的定居和生长发育决定了种群的天然更新和群落动态的维持,也很大程度上决定了森林群落演替方向和植被的恢复过程[1]。幼苗是植物生活史中最脆弱的时期,对环境变化反应较为敏感[2]。降低幼苗死亡率也是植被恢复中的关键[3]。土壤水分含量是影响幼苗存活最重要的环境因子之一,旱季降水的减少会抑制幼苗的生长甚至造成幼苗死亡[4]。在气候变化的大背景下,了解水分条件变化和干旱事件对幼苗死亡率的影响,将有助于预测未来森林的更新和演替。之前对幼苗更新研究多为幼苗天然更新的影响因子分析[4-8],也有模拟降水变化对幼苗生长的影响[9],研究幼苗在自然干旱状态下的表现将有助于理解控制实验结果并为控制实验设计提供思路。
水分条件是影响生态系统初级生产力和碳汇功能的重要环境因子[10-12],也是影响树木幼苗成活与生长的重要限制因子[13-14]。而全球气候变化将可能改变很多地区的降水格局并导致更多的地区受到更为严重的干旱胁迫[15-16]。干旱是指在相对广阔的地区长期无降水或降水异常少的气候背景下,水分供求严重不足的现象[17]。在2009年底至2010年初,我国西南五省市遭受了百年一遇的干旱,其中云南是受干旱影响较为严重的地区之一[18-20]。如果这一地区在未来干旱频率增加,强度加大[21],植物的生长、幼苗的存活和森林更新也将受到显著影响[22]。而该地区森林生态系统中幼苗对水分条件变化的响应尚未得到很好研究。此外,树木木材密度和树木的抗旱性及其在干旱条件下的死亡率密切有关[23],因而也很可能可以用来指示幼苗的抗旱性。因此,了解水分条件变化对森林幼苗动态的影响,探讨木材密度与幼苗在干旱环境下死亡率的关系,对预测和解释森林对长期气候变化的响应和制定有效森林保护和管理策略具有重要意义。
常绿阔叶林是我国亚热带地区最主要的原生植被类型,而目前在世界范围内对亚热带常绿阔叶林生态系统过程的了解相对较少。位于中国西南以木果石栎为标志的云南哀牢山中山湿性常绿阔叶林(以下简称常绿阔叶林)连接成片,林相完整,结构复杂,生物资源丰富,且地势较为平坦,是极具代表性的西南亚热带常绿阔叶林[24]。本文以哀牢山中山湿性常绿阔叶林的林下乔木幼苗为研究对象,研究了其基本形态特征和幼苗死亡率年际动态,并分析了其和水分条件之间的相关关系,旨在:1)了解亚热带森林林下幼苗对水分条件变化的敏感性并探讨水分条件和极端干旱事件对幼苗死亡率的影响;2)建立木材密度和幼苗应对干旱能力、幼苗死亡率之间的关系;3)为探讨气候变化对幼苗更新和森林演替影响提供依据。
1 研究方法
1.1 研究地概况
本研究在哀牢山国家级自然保护区北段的徐家坝地区开展,所选样地是中国科学院哀牢山亚热带森林生态系统定位研究站的永久监测样地,地理位置为24.32N,101.01E,海拔2488m,本区旱季(11月—4月)、雨季(5月—10月)分明,属于西南季风气候。年平均气温11.0℃,最热月(7月)平均气温15.3℃,最冷月(1月)平均气温5.0℃,极端最高气温25.0℃,极端最低气温-8.3℃,≥10.0℃积温为3420℃,霜期达半年(190d);年平均相对湿度83%,年平均降水量1931.1mm,年日照时数1239h,其主要的气候特征是长冬(5个月)无夏,春秋(7个月)相连,全年温凉潮湿,水湿资源丰富。林地土壤为山地黄棕壤[25]。
原生的常绿阔叶林乔木树种主要由壳斗科(Fagaceae)、茶科(Theaceae)、樟科(Lauraceac)和木兰科(Magnoliaceae)组成。壳斗科的优势树种有木果柯(Lithocarpusxylocarpus)、硬斗柯(Lithocarpushancei)、变色锥(Castanopsiswattii)等组成;茶科的优势树种有南洋木荷(Schimanoronhae)、舟柄茶(Hartiasinensis)和小花山茶(Camelliaforrestii)等组成;樟科的优势树种有黄心树(Machilusbombycina)、多果新木姜子(Neolitseapolycarpa)、滇润楠(Machilusyunnanensis)和鸭公树(Neolitseachuii)等组成;木兰科的主要优势树种有红花木莲(Manglietiainsignis)、多花含笑(Micheliafloribunda)等组成;另外,乔木层还有大花八角(Illiciummacranthum)、多花山矾(Symplocosramosissima)等物种组成。该植被的垂直分布为乔木层、灌木层和草本层,乔木层树高25—30m,平均盖度达90%以上。灌木层以箭竹(Sinarundinarianitida)为优势种,盖度达75%左右;草本层优势种有条穗苔草(Carexnemostachys)、粗齿冷水花(Pileasinofasciata)、金凤仙(Impatienscyathiflora)、华中瘤足蕨(Plagiogyriaeuphlebia)、平卧蓼(Polygonumstrindbergii)、球序蓼(Polygonumwallichii)和紫花沿阶草(Ophiopogonwallichianus)等组成,盖度约40%左右[26]。
1.2 研究方法
1.2.1 降水量和土壤平均质量含水量测定
降水量和土壤质量含水量是中国科学院哀牢山亚热带森林生态系统研究站的常规监测数据。降水量的监测采用口径是20 cm的雨量筒在雨天的08:00、14:00和20:00各观测1次;旱季降水量是上一年的11月至本年4月的降水量之和,雨季降水量即本年5月至10月的降水量总和,日降水量大于0.5mm的记为降水日数。在样地内的3个固定剖面采样点上自2005年1月至2011年12月进行采样(每个采样剖面由地表至地下),每个月采样1次,采回来的土样立刻带回实验室用BL610电子天平(Sartorius Inc., Germany)称重,再在烘箱内以105℃的恒温烘烤24h后称其干重,土壤采样及质量含水量计算参照文献[27]的方法。本研究考虑幼苗根系较浅,所用的含水量数据是由地表至地下20cm的土壤含水量平均值。因为旱季初期(11、12月)土壤储存水较多,而雨季初期(5月)土壤水分补充需要一定时间,以1至5月的土壤平均含水量来表示缺水季节(旱季)的土壤含水量。
1.2.2 幼苗形态测定
本研究根据乔木小苗的地面高度来定义幼苗[28],即将高度≤50cm的小苗作为幼苗研究对象[28]。为测定个体数较多的主要树种幼苗的形态特征和木材密度,于2013年在样地附近林下随机选择样地中丰富度最高的黄心树、多果新木姜子、多花山矾、鸭公树和大花八角的幼苗各六株,将其全株挖出,测出地上部分的高度及地下部分自然状态下的最大垂直深度。幼苗主干的木材密度采用饱和排水法测定[29]:将以上5种幼苗的主干切取5cm长度的样本,去掉皮和髓心,放水中浸泡48h后测出其体积,之后在烘箱内以80℃恒温烘至绝干,再称取干重,用下式计算出幼苗主干的木材基本密度:
Pr=M0/Vmax
式中,Pr为木材密度;M0为木材的干重;Vmax为木材含水量饱和时的体积。
1.2.3 幼苗动态调查
本研究的幼苗数据(2004年12月至2011年12月)为哀牢山亚热带森林生态系统研究站的常规监测。样地是该研究站的永久监测样地,面积为1hm2,该样地设为10 m×10 m的Ⅱ级样方100个,在其中的25个10 m×10 m样方内各选定一个2m×2m的幼苗更新调查的小样方进行长期定点监测,定期观测幼苗数量、物种、坐标、基径和高度。幼苗测定均在每年的12月下旬进行。本研究以小样方为统计单位,其中当年新生的幼苗数量不用于计算当年的幼苗死亡率。年度幼苗死亡率按以下公式计算[28]:
M=(d0-d) ×100/d0
式中,M为幼苗死亡率;d0为前一年幼苗总数;d为当年的幼苗数(不包括当年新生的幼苗数)。本研究分析了总幼苗和丰富度较高的5种乔木幼苗的死亡率。总幼苗为研究样地所有乔木幼苗,包括5种常见乔木幼苗树种之外的其它树种。
1.3 数据处理
形态特征、相对丰度、木材密度的种间差异,总幼苗和各树种幼苗死亡率的年际差异,各年各物种间死亡率的差异分别用One-way ANOVA分析。用Persons相关分析幼苗死亡率(总幼苗、黄心树、多果新木姜子、多花山矾、鸭公树和大花八角)与旱季降水量、旱季降水日数及旱季幼苗根系所在土层(0—20cm)土壤质量含水量间的相关性以及物种间2010年幼苗死亡率和木材密度之前的相关性。所有统计分析均在SPSS 18.0软件中进行。
2 结果与分析
2.1 2005—2011年哀牢山常绿阔叶林降水量、降水日数及浅层土壤质量含水量月动态
哀牢山常绿阔叶林月降水量与降水日数趋势较为一致(图1),降水量和降水日数在12—3月最少,其中2010年旱季连续3个月(12—2月)降水日数均为1d。2010年旱季降水量少且延续时间长,从2009年9月至2010年5月,降水量一直很少。2010年旱季3—5月幼苗根系所在浅层土壤(0—20cm)质量含水量也为有观测以来最低(图1)。
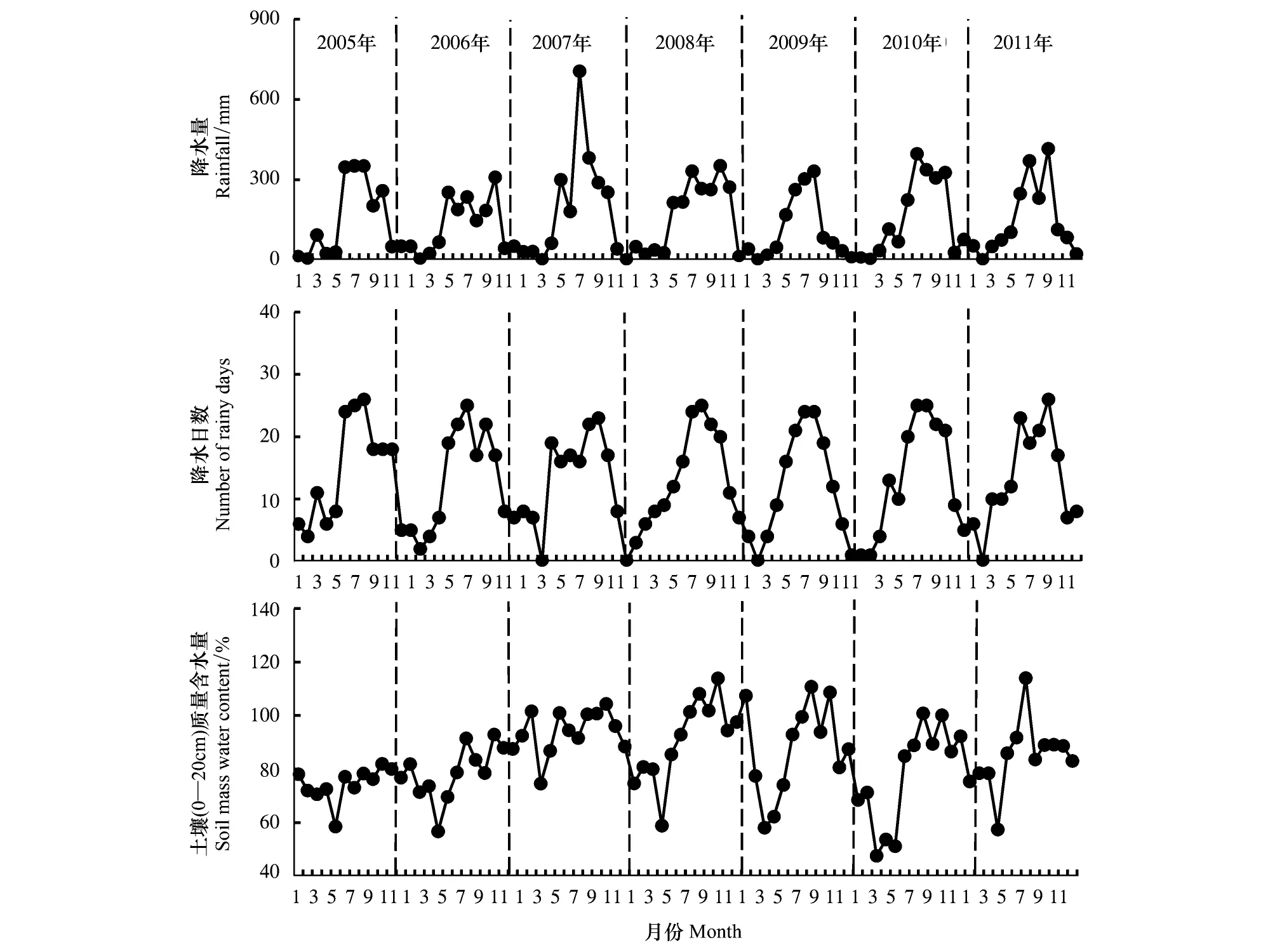
图1 2005—2011年降水量、降水日数及土壤(0—20 cm)质量含水量月动态Fig.1 Seasonal dynamics of rainfall, number of rainy days and soil (0—20 cm) mass water content from 2005 to 2011
2.2 5种乔木幼苗的形态特征
黄心树、多果新木姜子、多花山矾、鸭公树和大花八角的幼苗平均高度在27.85—37.08cm之间(表1),树种间无显著差异。根系深度在14.10—20.78cm之间,黄心树幼苗的根系最深,不过种间差异不显著。幼苗主干的木材密度在0.50—0.65g/cm3之间,其中多花山矾的木材密度最大,大花八角的木材密度最小并显著小于其它物种。5种乔木幼苗中,黄心树相对丰度(占总幼苗数的百分比)最高(64.35%),并显著高于其它4个树种(表1)。
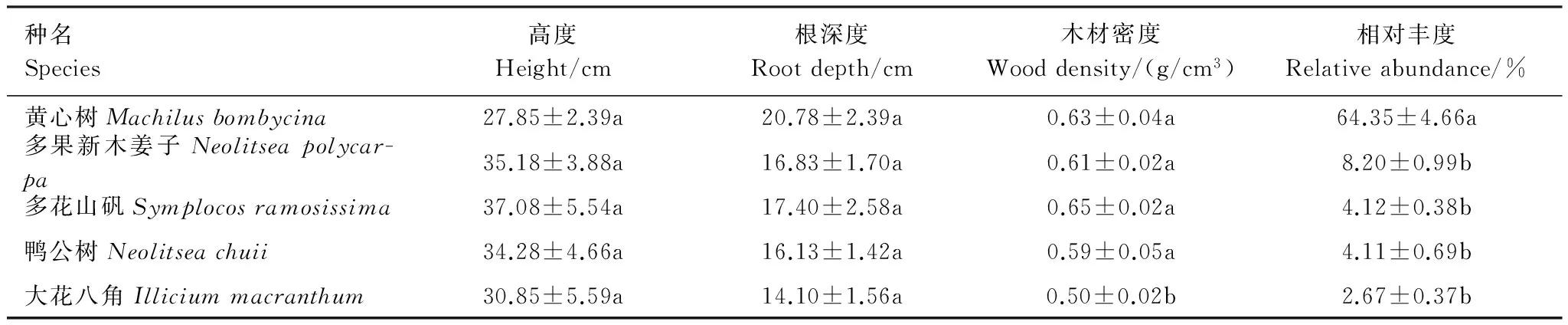
表1 5种常见树种幼苗的形态特征、木材密度和相对丰度Table 1 Seedling morphological characters, wood density and relative abundance of 5 common tree species
平均值±标准误;同一列间,相同小写字母表示种间差异不显著(P> 0.05)
2.3 幼苗死亡率年际变化
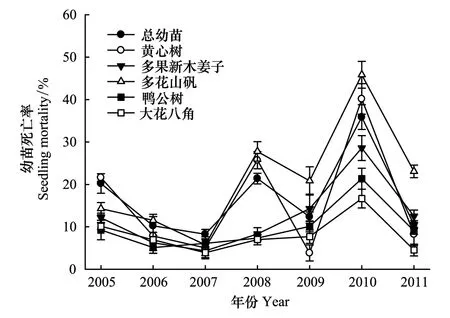
图2 2005—2011年幼苗死亡率的年际变化Fig.2 Dynamics of seeding mortality from 2005 to 2011
年际间,2010年总幼苗和丰富度较高的5种乔木幼苗死亡率均显著高于往年(P< 0.05),是2009年的2—10倍。2008年总幼苗、黄心树、多花山矾的幼苗死亡率显著低于2010年(P< 0.05)但显著高于其它各年(P< 0.05)。各树种间,多花山矾幼苗在2010年死亡率最高,黄心树次之,大花八角的死亡率最低,多花山矾和黄心树的死亡率显著高于其它3个物种(P< 0.05,图2)。
2.4 幼苗死亡率与气候因子的相关性
年际间,总幼苗死亡率与旱季降水日数有显著的负相关关系,即旱季降水日数越多幼苗总体死亡率越低(表2)。年际间,2010年旱季降水日数最低(26日),幼苗死亡率最高(35.86%),表明2010年干旱显著影响了常绿阔叶林幼苗的存活。黄心树、多果新木姜子、鸭公树和大花八角的幼苗死亡率与旱季降水日数有显著的负相关关系,多花山矾幼苗死亡率与旱季降水日数有极显著的负相关关系,即旱季降水日数越多,5种乔木幼苗死亡率越低。总幼苗死亡率、多果新木姜子、大花八角幼苗死亡率与旱季土壤平均质量含水量(0—20mm)也有显著的负相关关系(表2)。

表2 林下幼苗死亡率和旱季降雨、降雨日数及土壤含水量之间的Person相关系数Table 2 The Persons correlations between seedling mortality and rainfall, number of rainy days, and soil water content in the dry season
*P< 0.05; **P< 0.01
2.5 幼苗死亡率与主干木材密度的种间关系
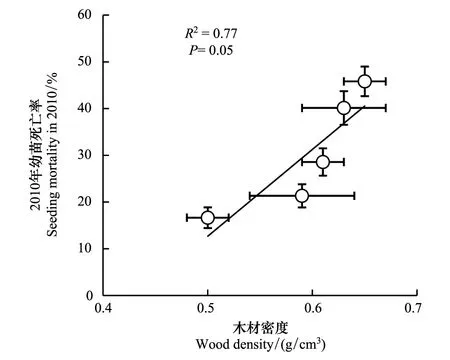
图3 2010年幼苗死亡率和木材密度之间的种间相关关系Fig.3 The interspecific relationship between seedling mortality in 2010 and wood density
在种间,幼苗在2010年死亡率与木材密度有线性正相关关系(图3),即木材密度高的树种幼苗在2010年死亡率更高。这表明幼苗主干的木材密度对幼苗在干旱中的死亡率有较强的指示作用。
3 讨论
影响林下幼苗死亡率和天然更新的因素众多,其中环境水分条件是最重要的因素之一,水分缺乏会直接导致幼苗的死亡[30]。研究表明,2010年西南干旱中哀牢山常绿阔叶林树木并未遭遇水分胁迫,但是草本层叶面积指数显著低于往年[31]。而同草本层一致,林下幼苗也受到了这次干旱事件的较大影响。哀牢山在2009年底至2010年初西南干旱中,半年多相对较少的降水使2010年旱季幼苗根系所在土层(0—20cm)的土壤质量含水量也是有观测以来最低,同时总体和各树种(黄心树、多果新木姜子、多花山矾、鸭公树和大花八角)幼苗死亡率也是有观测以来最高。此外,本研究还表明了木材密度大的树种幼苗在干旱中死亡率较高,不同树种幼苗在干旱中差异巨大的死亡率也表明未来降水格局的改变可能会通过影响幼苗动态影响森林的更新、演替和组成。
幼苗对降雨缺乏引起的水分亏缺比大树更为敏感,因而水分是影响森林更新的重要因子[32]。因为幼苗还未形成较深的根系,比如哀牢山常绿阔叶林5种幼苗根系深度在14.10—20.78cm之间,幼苗难以利用深层土壤水和地下水,所以其对降雨频度依赖较大,对环境水分条件变化极为敏感[6]。此外,哀牢山常绿阔叶林林下幼苗大部分时期处以阴凉潮湿的环境中[25],这可能也是其对环境水分条件变化很敏感的一个原因。哀牢山林下乔木幼苗死亡率与旱季降水日数、旱季幼苗所在土层含水量有显著的负相关关系也支持了幼苗死亡率对水分条件变化很敏感的结论。而乔木幼苗死亡率和旱季降水量之间却无显著相关性,可能是因为降水频度(降水日数)比绝对降雨量对维持浅层土壤较好水分状况更为重要。和本研究结果一致,鼎湖山亚热带常绿阔叶林对锥栗(Castanopsischinensia)和荷木(Schimasuperba)、黄果厚壳桂(Cryptocaryaconcinna)群落的幼苗死亡率也对环境水分条件变化极为敏感[4]。
木材密度和树木的抗旱性密切相关[23],研究表明其和幼苗在干旱中死亡率也有很好的相关性,因而木材密度也是一个可以指示幼苗抗旱能力、并用来预测幼苗死亡率的重要功能性状。这可能是因为木材密度和储水能力之间存在很强的负相关关系,即木材密度大的树种储水能力较弱[33]。木材密度及其枝条储水能力同时也和树木枝条在干旱条件下的死亡率存在很大的关系[23]。多花山矾和黄心树主干的木材密度较高,二者2010年幼苗死亡率均较高,这可能是其较弱的主干储水能力使其对干旱反应较为敏感。相反,大花八角幼苗主干的木材密度较小,其在2010年死亡率相比往年增加有限。黄心树虽然幼苗数量较多,但在干旱年份死亡率极高,导致幼苗数量急剧减少,在干旱频度增加的情况下将处于较以往不利的地位。此外黄心树和多花山矾幼苗在2008年也具有很高的死亡率,这是因为2008年旱季降水量也较少,而其较高的木材密度和对应较低的储水能力使其在2008年的降水状况下就表现出较高的死亡率。相比之下,低木材密度的大花八角在2008年死亡率和一般年份并无差异。如果未来干旱频度不断增加,木材密度较高的黄心树和多花山矾幼苗的相对丰富度可能会降低,而木材密度较低的大花八角的丰富度可能会呈现增加趋势。因而,极端气候事件的增多将可能通过影响幼苗动态来影响森林的树种组成。
综上所述,哀牢山亚热带常绿阔叶林林下幼苗死亡率和旱季降水日数及旱季幼苗根系所在土层相对含水量显著负相关。2010年的西南干旱使林下幼苗死亡率急剧上升,但是不同树种的幼苗表现存在很大差异,且幼苗死亡率和幼苗主干木材密度正相关:木材密度高的多花山矾和黄心树在2010年干旱中幼苗死亡率最高,而低木材密度的大花八角死亡率仍较低。如果未来该地区干旱频度和强度增加,大花八角的丰富度可能增加,而多花山矾、黄心树的丰富度可能减小。总之,哀牢山常绿阔叶林林下幼苗的死亡率对水分条件变化十分敏感,地区性干旱虽对哀牢山原生常绿阔叶林树木的影响较小[27],但可能会通过影响幼苗的死亡率和动态来影响整个森林的更新、演替和组成。
致谢:哀牢山亚热带森林生态系统研究站提供幼苗和气象数据,在幼苗的野外调查和气象数据的积累中得到了李达文、杨文争和罗鑫的支持,特此致谢。
[1] 潘春柳, 赖家业, 黎向东, 石海明. 单性木兰种子雨与天然更新的初步调查. 生态学杂志, 2008, 27(12): 2235-2239.
[2] 彭闪江, 黄忠良, 彭少麟, 欧阳学军, 徐国良. 植物天然更新过程中种子和幼苗死亡的影响因素. 广西植物2004, 24(2):113-121.
[3] Sánchez-Coronado M E, Coates R, Castro-Colina L, Castro-Colina L, de Buen AG, Paez-Valencia J, Barradas V L, Huante P, Orozco-Segovia A. Improving seed germination and seedling growth ofOmphaleaoleifera(Euphorbiaceae) for restoration projects in tropical rain forests. Forest Ecology and Management, 2007, 243(1): 144-155.
[4] 黄忠良, 彭少麟, 易俗. 影响季风常绿阔叶林幼苗定居的主要因素. 热带亚热带植物学报, 2001, 9(2): 123-128.
[5] 白志强, 刘华, 张新平, 刘端, 郭仲军. 新疆额尔齐斯河流域杨树幼苗天然更新影响因子分析. 西北林学院学报, 2011, 26(1): 98-102.
[6] 陈迪马, 潘存德, 刘翠玲, 常静, 尹锴. 影响天山云杉天然更新与幼苗存活的微生境变量分析. 新疆农业大学学报, 2005, 28(3): 35-39.
[7] Fahey T J, Battles J J, Wilson G F. Responses of early successional northern hardwood forests to changes in nutrient availability. Ecological Monographs, 1998, 68(2): 183-212.
[8] Thompson K, Grime J P. A comparative study of germination responses to diurnally-fluctuating temperatures. Journal of Applied Ecology, 1983, 20(1): 141-156.
[9] 肖春旺, 周广胜. 毛乌素沙地中间锦鸡儿幼苗生长、气体交换和叶绿素荧光对模拟降水量变化的响应. 应用生态学报, 2001, 12(5): 692-696.
[10] 郝彦宾, 王艳芬, 崔骁勇. 干旱胁迫降低了内蒙古羊草草原的碳累积. 植物生态学报, 2010, 34(8): 898-906.
[11] Tao F L, Hayashi Y, Zhang Z, Sakamoto T, Yokozawa M. Global warming, rice production, and water use in China: developing a probabilistic assessment. Agricultural and Forest Meteorology, 2008, 148(1): 94-110.
[12] 卢丽萍, 程丛兰, 刘伟东, 覃志豪. 30年来我国农业气象灾害对农业生产的影响及其空间分布特征. 生态环境学报, 2009, 18(4): 1573-1578.
[13] Morgan J M. Osmoregulation and water stress in higher plants. Annual Review of Plant Physiology, 1984, 35(1): 299-319.
[14] Hare P D, Cress W A, Van Staden J. Dissecting the roles of osmolyte accumulation during stress. Plant, Cell and Environment, 1998, 21(6): 535-553.
[15] IPCC.Climate Change: The Scientific Basis//Hougton JT, Dung Y, Griggs DJ, Noguer M, Vander Linden PJ, Dui X, Maskell K, Johson CA, eds. Contribution of Working Group I. Third Assessment Report of Intergovermental Panel on Climate Change. Cambridge: Cambridge University Press, 2007.
[16] Bouwman A F, Van Vuuren D P, Derwent R G, Posch M. A global analysis of acidification and eutrophication of terrestrial ecosystems. Water, Air, and Soil Pollution, 2002, 141(1/4): 349-382.
[17] 李新周, 刘晓东, 马柱国. 近百年来全球主要干旱区的干旱化特征分析. 干旱区研究, 2004, 21(2): 97-103.
[18] 王小军. 应对西南干旱的思考. 中国水利, 2010, (7): 11-13.
[19] Qiu J. China drought highlights future climate threats. Nature, 2010, 465(7295): 142-143.
[20] Stone R. Severe drought puts spotlight on Chinese dams. Science, 2010, 327(5971): 1311-1311.
[21] Wang W X, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta, 2003, 218(1): 1-14.
[22] HeisiEr-White J L, Knapp A K, Kelly E F. Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia, 2008, 158(1): 129-l40.
[23] Zhang Y J, Meinzer F C, Hao G Y, Scholz F G, Bucci S J, Takahashi F S, Villalobos-Vega R, Giraldo J P, Cao K F, Hoffmann W A, Goldstein G. Size-dependent mortality in a neotropical savanna tree: The role of height-related adjustments in hydraulic architecture and carbon allocation. Plant, Cell and Environment, 2009, 32(10): 1456-1466.
[24] 李宏文. 哀牢山徐家坝地区植被概况//吴征镒, 曲仲湘, 姜汉侨. 云南哀牢山森林生态系统研究. 昆明: 云南科技出版社, 1983: 63-73.
[25] 邱学忠, 谢寿昌. 哀牢山森林生态系统研究. 昆明: 云南科技出版社, 1997.
[26] 钱洪强. 哀牢山徐家坝地区常绿阔叶林结构分析//吴征镒, 曲仲湘, 姜汉侨. 云南哀牢山森林生态系统研究. 昆明: 云南科技出版社, 1983: 118-150.
[27] 杞金华, 章永江, 张一平, 刘玉洪, 杨秋云, 宋亮, 巩合德, 鲁志云. 哀牢山常绿阔叶林水源涵养功能及其在应对西南干旱中的作用. 生态学报, 2012, 32(6): 1692-1702.
[28] 巩合德, 杨国平, 鲁志云, 刘玉洪, 曹敏. 哀牢山常绿阔叶林乔木树种的幼苗组成及时空分布特征. 生物多样性, 2011, 19(2): 151-157.
[29] 徐清乾, 许忠坤, 周小玲, 陈善喜. 四川桤木木材密度和纤维特性研究. 湖北林业科技, 2008, 152(4): 12-15.
[30] 李芳兰, 包维楷, 庞学勇, 冷俐. 岷江干旱河谷5种乡土植物的出苗、存活和生长. 生态学报, 2009, 29(5): 2219-2230.
[31] 杞金华, 章永江, 张一平, 刘玉洪, 鲁志云, 武传胜, 温韩东. 西南干旱对哀牢山常绿阔叶林凋落物及叶面积指数的影响. 生态学报, 2013, 33(9): 2877-2885.
[32] Schupp E W, Milleron T, Russo S E. Dissemination limitation and the origin and maintenance of species-rich tropical forests//Levey D J, Silva W R, Galetti M, eds. Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. Wallingford: CABI Publishing Press, 2002: 19-33.
[33] Scholz F G, Bucci S J, Goldstein G, Meinzer F C, Franco A C, Miralles-Wilhelm F. Biophysical properties and functional significance of stem water storage tissues in neotropical savanna trees. Plant, Cell and Environment, 2007, 30(2): 236-248.
The influence of changes in water availability on seedling mortality of a subtropical evergreen broadleaf forest on Ailao Mountain
QI Jinhua1,2, ZHANG Yongjiang1,3,*, ZHANG Yiping1,2, LIU Yuhong1,2, LU Zhiyun1,2, WU Chuansheng1,2,WEN Handong1,2
1XishuangbannaTropicalBotanicalGarden,ChineseAcademyofSciences,Menglun666303,China2AilaoshanStationforSubtropicalForestEcosystemStudies,ChineseAcademyofSciences,Jingdong676200,China3DepartmentofOrganismicandEvolutionaryBiology,HarvardUniversity,Cambridge,MA02138,USA
In order to understand the influence of changes in water availability on the seedling dynamics of subtropical forests and the relationship between wood density and seedling mortality during a prolonged drought period, we studied seedling mortality patterns across all species and of 5 common tree species from an evergreen broadleaf forest on Ailao Mountain, southwest (SW) China from 2005 to 2011. The five tree species studied wereMachilusbombycina,Neolitseapolycarpa,Symplocosramosissima,Neolitseachuii, andIlliciummacranthum. The relationships between seedling mortality and rainy days, rainfall, soil water mass content (0—20 cm) during the dry season, and wood density were also analyzed. Our results showed that 1) Seedling mortality was negatively correlated with number of rainy days in the dry season and dry season soil water mass content, but not with total dry season rainfall; 2) The seedling mortalities of all species as a whole and of each of the 5 species studied reached a historical maximum in the 2010 SW China extreme drought, which were 2 to 10 times more intense than for 2009 (MachilusbombycinaandSymplocosramosissimahad the highest seedling mortalities whileIlliciummacranthumhad the lowest in 2010); 3) Across species, the seedling mortality in the 2010 drought was positively correlated with species-specific wood densities; species with higher wood densities had higher seedling mortalities. Previous studies have shown that trees in this forest did not experience severe drought stress during the 2010 SW China drought. However, the present study suggests that the seedlings were severely influenced by the rainfall anomaly during the 2010 dry season, probably owing to their shallow rooting depths and consequently high sensitivities to changes in rainfall. Additionally, because of the lower seedling mortalities in lower wood density species, the potential increase in drought frequency and severity in the future may increase the relative abundance of low wood density species, which may potentially lead to changes in forest species composition.
drought; seedling dynamics; soil mass water content; rainfall; wood density
中国科学院哀牢山亚热带森林生态系统研究站研究基金
2013-06-10;
日期:2014-05-16
10.5846/stxb201306101572
*通讯作者Corresponding author.E-mail: zhangyj@xtbg.ac.cn
杞金华, 章永江, 张一平, 刘玉洪, 鲁志云, 武传胜, 温韩东.水分条件变化对哀牢山亚热带常绿阔叶林林下幼苗死亡率的影响.生态学报,2015,35(8):2521-2528.
Qi J H, Zhang Y J, Zhang Y P, Liu Y H, Lu Z Y, Wu C S,Wen H D.The influence of changes in water availability on seedling mortality of a subtropical evergreen broadleaf forest on Ailao Mountain.Acta Ecologica Sinica,2015,35(8):2521-2528.
