水体无机碳升高对蓝绿藻生长和种群竞争的影响研究进展
2015-03-12张君枝王齐马文林陈汉赵远玲张艳娜
张君枝,王齐,马文林*,陈汉,赵远玲,张艳娜
1. 北京应对气候变化研究和人才培养基地//城市雨水系统与水环境教育部重点实验室//北京建筑大学,北京 100044;2. 中国石油勘探开发研究院,北京 100083
水体无机碳升高对蓝绿藻生长和种群竞争的影响研究进展
张君枝1,王齐1,马文林1*,陈汉1,赵远玲1,张艳娜2
1. 北京应对气候变化研究和人才培养基地//城市雨水系统与水环境教育部重点实验室//北京建筑大学,北京 100044;2. 中国石油勘探开发研究院,北京 100083
摘要:蓝绿藻是水体中浮游植物的重要组成部分,也是淡水与海洋生态系统中主要的初级生产者,近些年来,随着CO2排放的日益增加,全球气候发生着巨大的变化,极端气候事件频发,对水体中的无机碳含量也带来了影响,甚至直接影响了蓝绿藻的CCM机制。该机制能使细胞在核酮糖磷酸羧化氧化酶(RubisCO)固碳位点提高CO2浓度,以增加光合作用和减少光呼吸。文章以大气CO2浓度升高带来的气候变化为大背景,首先叙述了不同形态无机碳在开放水体中的平衡,阐述了大气、水体、底泥等水环境中碳循环过程及其对藻类产生影响的各种作用,总结了近年来模拟环境无机碳浓度增加的各种实验模拟技术及大气CO2浓度升高对蓝绿藻生长及种群竞争的研究方法的进展。通过对蓝绿藻利用无机碳的碳浓缩机制的机理的研究,无机碳进入蓝绿藻主要通过卡尔文循环,在碳酸酐酶的作用下,HCO3-、CO32-形态的无机碳可以转变为CO2,提高了RubisCO周围的CO2浓度,提高了藻利用无机碳的效率,但同时蓝绿藻的上述CCM机制也受到pH、光照、N、P等因素的影响。在此理论基础上讨论了无机碳对蓝绿藻细胞结构、细胞大小和个体增殖等方面的影响。研究发现,高浓度CO2促进细胞结构中蛋白核的增大,而细胞体积较小的藻类对CO2浓度响应更明显。由于CO2能被蓝绿藻细胞直接利用,因此CO2更能促进藻细胞个体的增殖。地表天然水体的pH在中性-弱碱性范围,因而更多的研究集中于蓝绿藻细胞对CO2、HCO3-的利用,继而直接影响了蓝绿藻的CCM机制。综上,虽然气候变化的影响已经有不少的研究,但是气候变化对于蓝绿藻的碳利用机制及细胞的微观变化的影响以及蓝绿藻在生长以及水华形成过程中对气候变化CO2浓度升高的响应机制仍需要进一步深入的研究。
关键词:气候变化;无机碳;蓝绿藻;CO2浓缩机制
引用格式:张君枝,王齐,马文林,陈汉,赵远玲,张艳娜. 水体无机碳升高对蓝绿藻生长和种群竞争的影响研究进展[J]. 生态环境学报, 2015, 24(7): 1245-1252.
ZHANG Junzhi, WANG Qi, MA Wenlin, CHEN Han, ZHAO Yuanling, ZHANG Yanna. A Review on the Effect of Inorganic Carbon on Blue-green Algae Growth [J]. Ecology and Environmental Sciences, 2015, 24(7): 1245-1252.
人类活动在不断的改变地球的生物化学循环,其中最主要的大尺度全球变化是近年来大气中CO2浓度的不断攀升(Low-Décarie et al.,2014)。近几年大气中CO2浓度以每年0.5%的速度递增,根据IPCC第五次报告,目前CO2体积分数已达到391×10-6,比工业革命前高约30%。据预测,若CO2排放不加控制,21世纪下半叶CO2浓度将加倍,大气温度将上升3 ℃左右(程丽华等,2005)。
气温的升高和水体有机物、营养元素输入的增多,水体中更多的有机碳会加速降解为无机碳(Rosemond等,2015),有机碳年输入被迅速耗尽,同时伴随着淡水湖泊pH的升高,一些湖泊逐渐由碳源转为碳汇(Finlay等,2015),所以水体中无机碳的转化潜力将大幅增加。大气升温会导致水体升温,水体表层温度升高对藻类的光合作用的影响已有不少研究,并且伴随着CO2的溶解增加,水体酸化与温度的耦合将会协同影响蓝绿藻在水生态系统中的分布及生长(Reichwaldt et al.,2012)。目前对于气候变化,更多的研究关注高温、风速、光照强度、水力条件、降水规律、干旱天气、盐度以及高有机碳含量对藻类生长以及群落演替的影响(Emelko et al.,2011;Paerl et al.,2012;Sinha et al.,2012;Tang et al.,2013;Zhang et al.,2012),很少关注到碳平衡导致的无机碳升高对淡水藻类带来的影响(O’Neil et al.,2012)。虽然淡水生态系统对全球碳平衡以及整个生物地球化学循环的影响要远小于海洋生态系统,但淡水生态系统与人类生产生活联系更加紧密,而目前还很少有关全球无机碳水平升高对淡水生态系统影响的研究(赵旭辉等,2012)。藻类在进行自身光合作用和代谢的过程中使得pH和碱度升高,水体内各种溶解态无机碳形式会产生平衡转化(支彦丽等,2008a)。
研究发现,在一定的CO2浓度水平和pH范围内,蓝藻和绿藻都有可能成为优势种群并引发水华(Su et al.,2015;支彦丽等,2008a)。在弱碱性介质中,藻类的净光合速率将主要取决于于对HCO3-的主动转运过程,所以在正常淡水中,藻类的对于无机碳的利用近80%为HCO3-(戴树桂,2006)。近些年来,许多学者在研究水体蓝藻暴发的模型,其中包含温度,分层,氮磷营养物水平等变量(Elliott,2012),但均未考量水体不同无机碳水平的影响。其原因在于无机碳对于淡水藻类生长的影响尚未被充分关注并进行实验研究。因此,研究无机碳在不同浓度水平下的不同组成形式以及对藻类的生长和种群关系的影响有着重要意义(支彦丽等,2008b),尤其是其中藻类对于各种形态无机碳转运的机理和影响因素。
本文概述了无机碳对于藻类影响的机理以及目前的研究进展,包括无机碳在开放水体中的平衡以及全球CO2水平升高的条件模拟实验的进展、蓝绿藻在水体中对于无机碳的浓缩机制以及无机碳对蓝绿藻细胞结构、大小及不同形态无机碳对蓝绿藻在种群竞争中的影响,通过对实验室或原位条件下无机碳水平升高对于蓝绿藻的影响分析,以期进一步认识气候变化对于水体无机碳水平的影响及蓝绿藻对气候变化的响应。
1 无机碳在开放水体中的平衡以及模拟环境无机碳浓度增长研究方法进展
1.1气候变化情境下无机碳在开放水体中的平衡
19、20世纪以来,大规模化石燃料的燃烧使得大气中CO2浓度升高,并且增长速率由20世纪的1%增长为现在的3%(O’Neil et al.,2012)。大气CO2体积分数从350×10-6增加到800×10-6时,会引起水体中pH和水体中溶解性无机碳(DIC)的变化。更细致的讲,大气中的CO2首先通过大气体系内传输导致水体表面CO2的浓度提高,导致水体溶解更多的CO2,碳酸盐组成发生变化,碳酸根离子浓度(cCO32-)降低,碳酸氢根离子浓度(cHCO3-)略有增加(Raven et al.,2014)。由于水体中无机碳主要以CO32-、HCO3-和CO2的形式存在,并存在溶解和解离平衡,所以水体中溶解性无机碳对大气CO2浓度升高的响应较为复杂(夏建荣等,2002a)。天然水体是开放体系,CO2与大气不断交换对于蓝绿藻的影响过程是长期的,如海水或者淡水长期的碳酸平衡对于蓝绿藻生长的影响,此时则认为CO2与水是处于平衡状态,可以更近似于真实情况。淡水和海水的碳酸平衡图非常相似。但由于离子强度的影响,海水的酸度常数比淡水的要大些,因为海水除含有溶解CO2外,还含有对海水有缓冲作用的硼酸(J.J.摩尔根,1987)。
水气界面碳交换理论是实验室模拟水体无机碳升高的重要基础。鉴于实验室进行藻类培养通常是基于开放体系的,而开放体系中的无机碳对于大气中CO2浓度升高的响应复杂,比较理想的模拟方法是配置高CO2浓度的模拟空气,对藻类生长的实验装置上方进行连续通气,可获得对于大气CO2浓度升高的直接模拟(Low-Décarie et al.,2014),但现阶段实验室实验较少采用。
在大气CO2浓度升高对于陆生植物的影响方面,已经有比较详细的研究(赵天宏等,2006)。在进行水华和赤潮的研究时一般认为,藻类不受水中无机碳限制,而是受以氮磷为主的营养盐、光照和浮游动物的捕食压力限制。然而,近年来的一些研究,包括室内培养的细胞和自然环境的浮游植物群落,均表明它们的生长和光合固碳受到水中CO2浓度的限制(陈雄文等,2003)。大气CO2浓度的升高可以通过开放水体的碳酸平衡进而影响到水体中藻类的生长,虽然这种影响会被碳酸平衡有所削弱,但是大幅度CO2的增长也会对藻类生长和固碳有所影响(姚臣谌等,2012;喻元秀等,2009)。水环境中纵向碳循环的过程如图1所示。
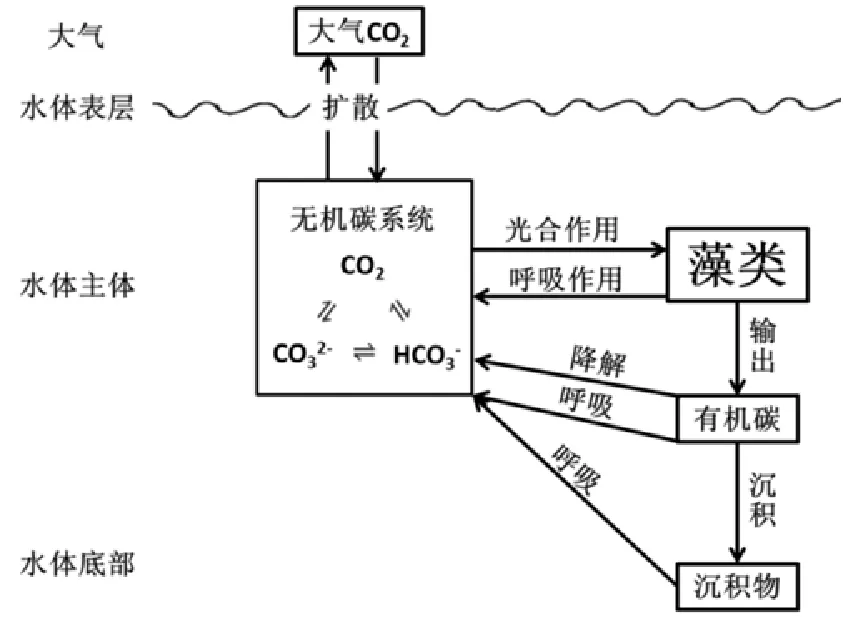
图1 水环境中碳循环的过程Fig. 1 The carbonate cycle in the water environment
1.2模拟环境无机碳浓度增长研究方法进展
目前实验室中的控制水体中CO2浓度的方法及其演替如表1,主要有:采用CO2加富器,使得当地空气CO2浓度上升到目标水平,对实验水体用模拟空气进行鼓气培养,在诸多实验室实验和太湖的原位试验中有所应用。其中,CO2浓度实验的时间控制也存在较大的差异,最短的培养时间仅为几小时,一般为3~6 d,而其他需要较长时间培养的实验,如在高CO2浓度环境中藻类水华研究和群落结构变化的观察,一般均采用半连续培养的方式进行(夏建荣,2006)。该法在实验室较容易实现,缺陷是对于实验装置的气水平衡有所影响,加快了水与气体的传质速率,进而影响到氧气等其他气体在水中的交换,对实验结果影响不易估计。
而另一种方法,通过添加碳酸氢盐来调节水中的碱度,再辅以pH调节,是实验室目前模拟无机碳升高情景常用的手段(夏建荣,2006),在HCO加富实验中,碱度的添加,因其简便易用、HCO指标易于控制而得到广泛应用(李鑫等,2011;孙凌等,2007),药剂的选择一般为NaHCO3和Na2CO3,添加量范围通常在100 μmol·L-1以内。实验中通过对于碱度和pH的测定,可以通过无机碳转化平衡计算出但问题是碱度的添加通常是以添加碳酸氢盐的形式存在,绝大多数实验又选用的NaHCO3。在提升HCO的同时升高了Na+的浓度。因部分藻类无机碳转运模型中谈及Na+浓度对于HCO转运速率的影响,所以选择模拟添加无机碳时添加碳酸氢钠时要慎重(Ghoshal et al.,2002)。由于目前对于无机碳对于藻类生长影响的研究多在实验室阶段,最大程度模拟真实水体环境下的无机碳水平以及平衡是最重要的。
上述几种方法都可以做到提高水体的无机碳水平,但问题是真实水体无机碳升高是建立在大气中CO2浓度大幅度上升的假设下的,如果只是单次的添加碳酸氢盐,可能在一定pH范围内造成水体无机碳以CO2的形式耗散出去,而并非被藻类所吸收,造成实验的极大误差。所以,在不同pH范围内,水体无机碳向外界耗散情况需要通过进一步的条件试验证明。目前可以通过调节实验水体pH或向实验装置内通入与之平衡的模拟空气,避免此类系统误差的出现。
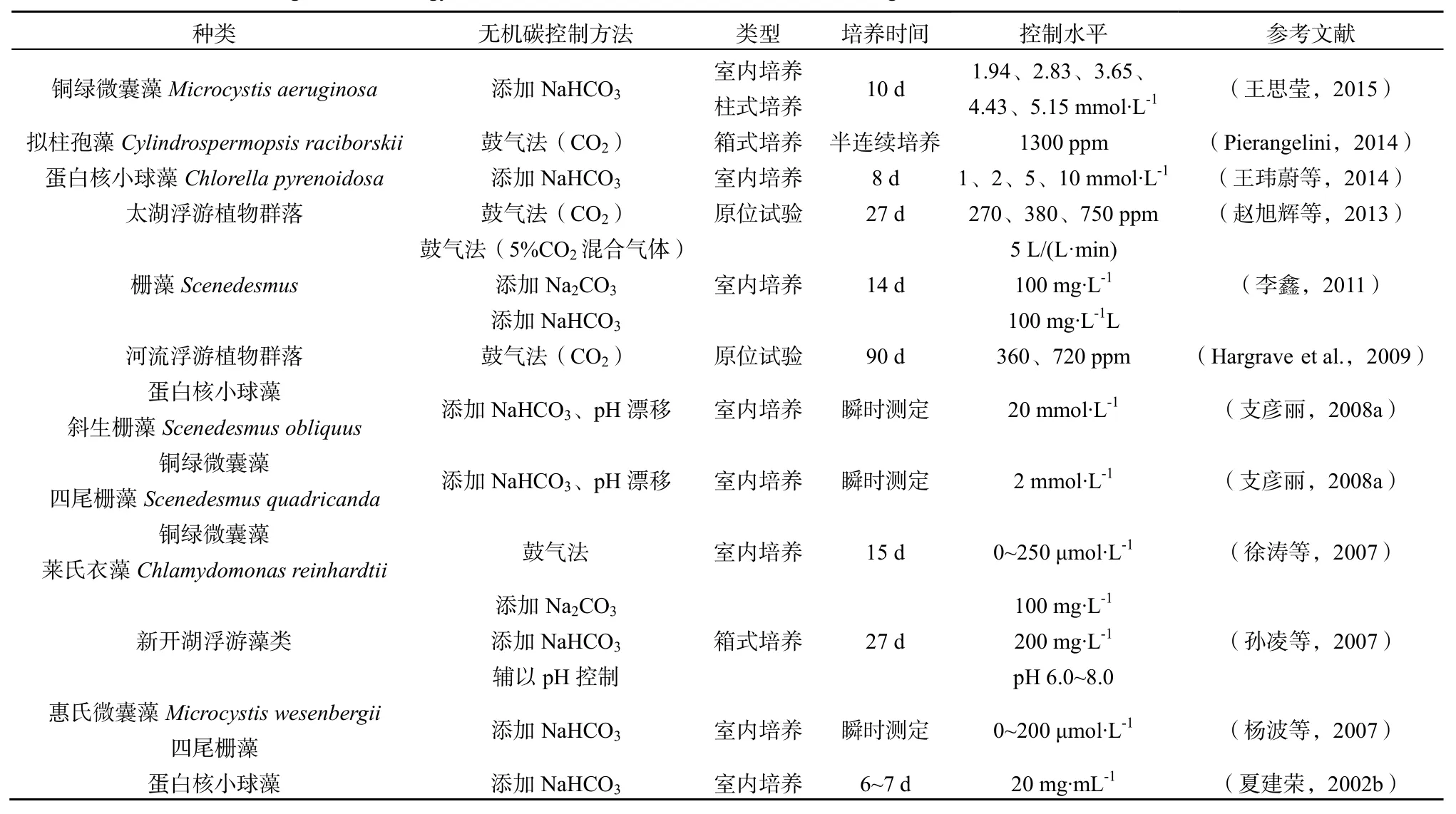
表1 模拟环境无机碳浓度增长技术发展概述Fig. 1 Technology Evolution of the Simulation of the Elevated Inorganic Carbon in the Environment
2 蓝绿藻利用无机碳的碳浓缩机制
藻类是地球上最古老的生物之一,在漫长的自然进化过程中,大气中的CO2浓度下降O2浓度上升,蓝绿藻的C3固碳作用中核酮糖-1, -5二磷酸羧化酶(RubisCO)对CO2的亲和力较低,但是蓝绿藻细胞光合作用对CO2的亲和力很高。这主要是因为蓝绿藻拥有几乎在所有光合生物中最有效的CO2浓缩机制(CCM,CO2-concentrating Mechanism)来提高羧体内CO2浓度以补偿其RubisCO对CO2亲和力的不足,进而在巨大自然选择的压力下获得优势(Bharti et al.,2014)。人们对于藻类CCM机制的认知从研究衣藻体内CO2的浓度高于体外40倍的机理开始(Winck et al.,2013),并且蓝藻的CCM机制较进化更加完全的绿藻更加有效,效率更高(O’Neil et al.,2012)。
无机碳进入蓝绿藻主要通过卡尔文循环(Calvin-Benson-Bassham cycle)(夏建荣等,2002a)。该循环中,1, 5-二磷酸核酮糖羧化酶/加氧酶(RubisCO)作为限速酶,将CO2固定在核酮糖-1, 5-二磷酸(RuBP)上,生成3-磷酸甘油酸(3-PGA)。因为CO2和O2具有化学相似性,所以通过RubisCO催化的固碳反应对O2高度敏感。然而,大气中O2含量远远超过CO2。为适应大气环境,自然界进化出CO2浓缩机制图2,即通过在细胞内高浓度积累CO2来提高RubisCO的固碳效率。在CCM中,藻细胞通过质膜主动转运HCO进入胞液,在细胞内通过碳酸酐酶的作用HCO转变为CO2(李小梅,2012),这样CCM可以将RubisCO周围CO2浓度提高到外界的500~1000倍以上。另外,羧酶体将RubisCO和碳酸酐酶(Carbonic anhydrase,CA)包裹在半透性外壳中,阻止了O2进入酶的活性位点,有利于RubisCO起羧化酶作用,抑制其氧化酶活性(成慧敏,2006)。同时提高了RubisCO周围CO2的浓度,而余下的CO2在向外扩散过程中被依赖于光能的CO2主动转运系统部分回收(成慧敏,2006;张宝燕等,2014)。同时大量无机碳跨膜流动可能起到耗散过剩光能,并维持胞内pH的作用。已有文献证明CCM在促进HCO3-吸收利用的同时,并不影响对CO2的正常利用(赵旭辉等,2012)。目前有文献发现CO2在真核藻类叶绿体中也有富集,但具体转运机理尚不明确(Winck等,2013)。
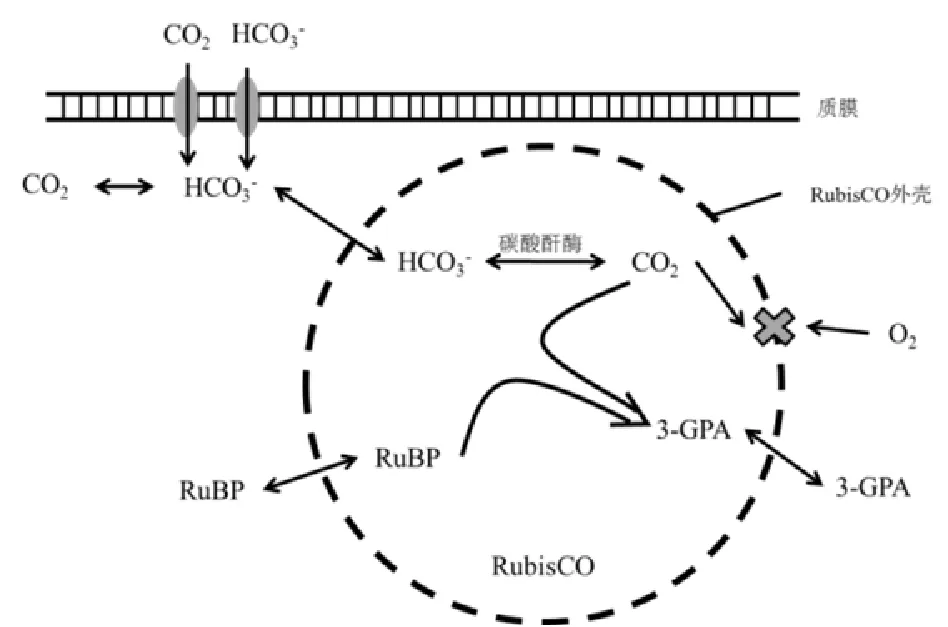
图2 蓝绿藻CO2浓缩机制Fig. 2 CO2-concentrating mechanism in blue and green algae,CCM
CCM机制同时受许多因素共同影响,包括无机碳、pH、光合成有效辐射(PAR)、紫外线(UV-B)、氮元素的形态和浓度、磷元素和铁元素的浓度,同时一些藻类的CCM还会受到温度和O2浓度的影响(Raven et al.,2014),藻类在夏季高日照情况下,CCM机制更加有效(Björn Rost,2006)。但温度与其他因素耦合对CCM机制的影响需要进一步研究。
当环境pH接近中性(6.0~7.5)时,很难观测到细胞CO2的直接转运利用(Moazami-Goudarzi et al.,2012)。即CO2浓度不足时,对于部分HCO3-在胞外CA的作用下解离成CO2扩散进入细胞,部分HCO3-则依赖离子泵主动运输进入细胞,同时也存在CO2的直接转运(Moazami-Goudarzi et al.,2012)。而胞内CA通过调节HCO3-和CO2之间的平衡,维持基质适宜pH。在碳限制条件下某些蓝藻可诱导产生多种转运子,形成互补的无机碳转运系统,从接近碳枯竭的基质中摄取无机碳进行光合作用(成慧敏,2006)当水中溶解的CO2过饱和时,水中pH也会相应下降,使得水中总溶解无机碳中的CO2/HCO3-比例增大(邹定辉等,2001),减弱浮游植物碳浓缩机制(CCM)的必要性(Winck等,2013;赵旭辉等,2013)。
营养物质的代谢与光合作用是两个紧密相连的过程,其中的氮限制将明显导致光合速率下降(夏建荣等,2002a)。无机碳的转运是一个需要光合作用提供能量的过程,而氮元素是制约蓝绿藻生长的主要营养元素,磷元素促进或抑制CCM的进程已有报道(Raven et al.,2014),但目前尚无明确结论。
3 无机碳对蓝绿藻生长及竞争关系的影响
3.1无机碳对蓝绿藻生长的影响
无机碳对于蓝绿藻的影响基本分为三个层次,分别为细胞结构、细胞大小和个体增殖方面。对于现有的研究在这几个方面加以梳理,见表2.
早期研究一般认为蓝绿藻必须通过水气界面交换和水体中的无机碳平衡才能接触CO2,并且CO2在水中的扩散速度远小于其在空气中的扩散速度。由于水体存在缓冲作用,传统观点认为蓝绿藻与典型陆生绿色植物相比,其增殖对大气CO2浓度升高的响应可能不明显。而这些已有的研究忽视了蓝绿藻在碳固定期间对于水体无机碳平衡的影响,并且此时水中溶解的CO2并未像之前设置的实验一样达到饱和。因此,理论上讲,大气CO2升高对于水体无机碳的影响必将显著提高蓝绿藻在淡水中的生物量。综合目前已有的研究,其对于大多数蓝绿藻包括小球藻属、双星藻属、栅藻属、葡萄藻属、绿球藻属、微囊藻属和螺旋藻属都有显著的影响(Singh,2014)。
从细胞结构看,研究发现高浓度的CO2会促进叶绿体增多,但同时促进蛋白核的形成和长大。随着CO2浓度的升高,藻细胞内叶绿体的数量也有逐渐上升的趋势(Ghoshal等,2002)。关于蛋白核的形成以及大小,在低浓度CO2培养下,蛋白核小球藻细胞内可见有明显的淀粉盘包围的蛋白核,当转移到高浓度CO2培养,细胞内蛋白核明显增大,直径增大1.5倍(Izumo等,2007;Singh,2014)。
细胞大小方面,根据藻类代谢规律,蓝绿藻的增殖速度与细胞的体积呈负幂函数关系。藻细胞体积越小,表面积越大,营养物质的在细胞中的扩散越充分,生物量和相对丰度也就越大。无机碳的补给情况与其它营养物质相似,同样受扩散速度的限制,因此体积较小的蓝绿藻可以将从水体无机碳水平升高的趋势中受益更多,例如隐球藻(Aphanocapsa)、隐杆藻(Aphanothece)、色球藻(Chroococcus)、腔球藻(Coelosphaerium)等(Singh,2014)。另外,有研究表明,气温在基础温度下每升高1 ℃,藻类直径下降2.5%,这也算是藻类对大气中CO2浓度升高的间接响应。
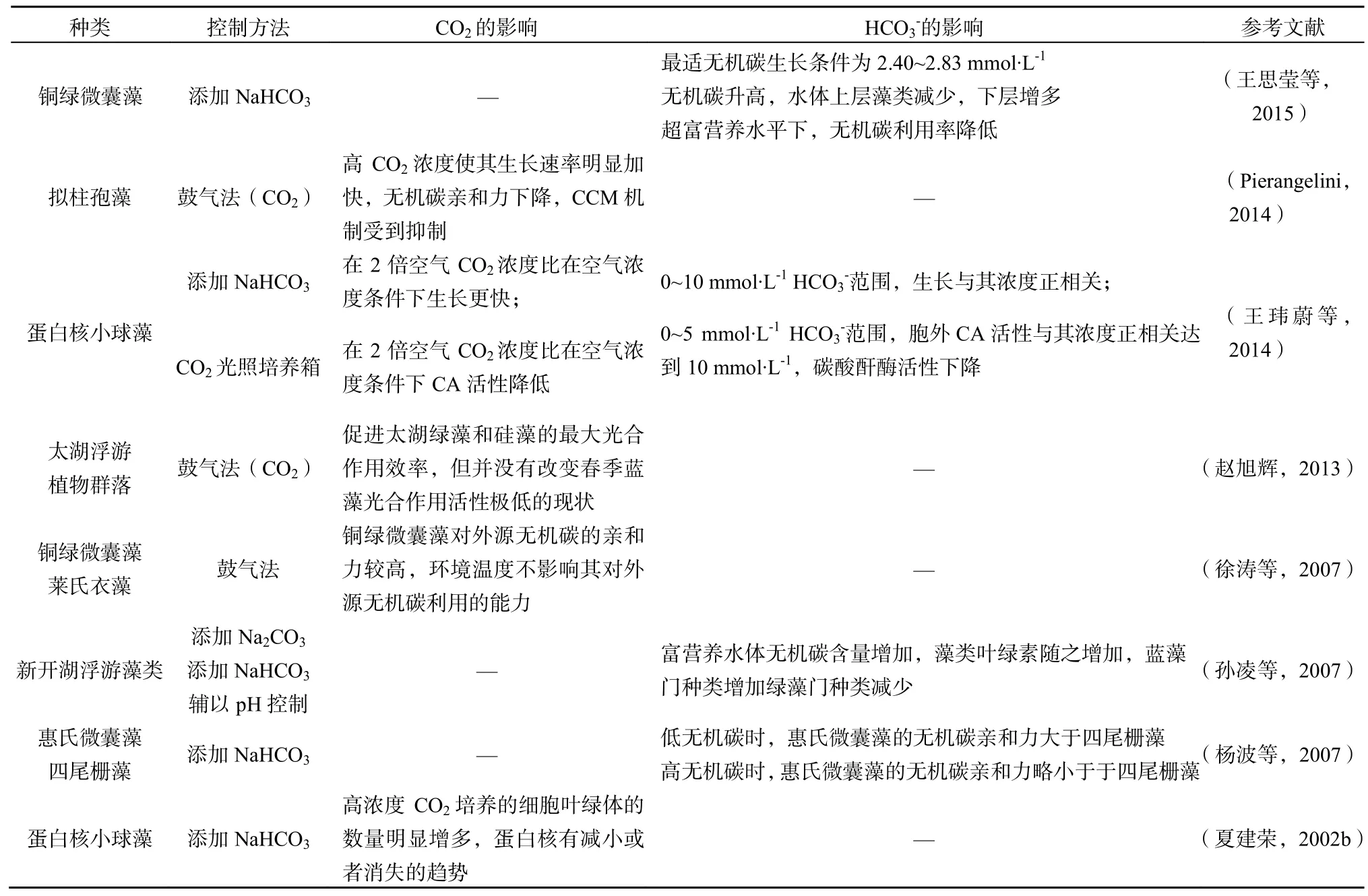
表2 无机碳对蓝绿藻生长及种群竞争的影响Fig. 2 The effects of inorganic carbon on the growth and species competition of blue-green algae
从个体增殖方面看,CO2浓度升高相比于HCO浓度升高更容易反映在藻类生长上面。理论上,根据藻类的CO2浓缩机制,在多种碳源中藻类更倾向于吸收水中的溶解CO2(赵旭辉等,2012),因这一过程的转运不消耗能量,CO2浓度增加后将强化这一过程。由于在实际条件下的原位水环境中,光和营养物质常常被认为是蓝绿藻的生长限制因子,藻细胞通过转运更多的水中溶解性CO2,减少了对HCO的摄取,因此这一部分主动运输的能量被节省下来,CO2浓度的升高对藻细胞的光能捕获能力也没有影响,所以藻类总体光合效率大幅提升。同时也表现出蓝绿藻生长对于大气中CO2浓度变化具有敏感性(夏建荣,2006),表现为藻类CCM机制必要性的降低,使得相关酶的活性有所降低。其中,随着CO2浓度的升高,胞外CA活性降低(详见2.2.1),说明该小球藻中存在胞外碳酸酐酶,并且其活性受高浓度CO2抑制(王玮蔚等,2014)。当CO2体积分数增加到更高的1%~5%时,部分绿藻的碳酸酐酶活性被明显抑制甚至完全丧失活性(李小梅,2012)。说明在高CO2浓度下,pH有所降低,藻类对于溶解CO2的利用增加,减少了对HCO的摄取,反映为胞外CA活性降低。从这方面看来,对于CCM机制较强的蓝藻,这一方面可能存在劣势,由于CCM机制受到抑制,导致蓝藻对HCO利用能力下降。另外,已有研究表明,高浓度CO2可以促进蛋白核小球藻的生长,因此目前微藻养殖多选择小球藻等绿藻(Brilman等,2013)。同时,若是在氮磷条件限制情况下,高CO2浓度的影响会被削弱,但仍会表现出生长促进作用。在贫营养湖泊内进行高CO2浓度培养的丝状绿藻,其生长速率在高CO2浓度下显著加快(Andersen et al.,2006)。
对于碳酸氢盐,其浓度的升高会使得绿藻的在水体中优势扩大并引发水华,从而导致浮游藻类多样性降低(孙凌等,2007)。实验证明,在低HCO水平时,惠氏微囊藻对外源CO2的代谢能力略大于四尾栅藻和菱形藻(杨波等,2007)。在0~10 mmol·L-1HCO范围内,HCO的浓度与藻类细胞密度成正相关(支彦丽等,2008b),蓝藻的最适碳酸氢盐浓度约为75 mmol·L-1,且最高耐受值可达250 mmol·L-1(Jaiswal et al.,2002)。当水华过程结束,水体无机碳会随着藻类种群的缩小,有机碳的不断分解而升高,甚至高过水华发生前的浓度,由于藻类的碱性代谢物减少,pH也随之降低(Hu等,2015)。
综上,高浓度CO2对于蓝绿藻主要起促进作用。具体表现为促进叶绿体的增多并部分抑制绿藻中蛋白核的生长,降低了CCM机制中的碳酸酐酶的活性,从而减少对于HCO的主动转运,间接提高了藻类的总体光合效率。在无机碳限制的情况下更易于水体中蓝藻的生长,因其CCM机制较绿藻更为有效;而当无机碳水平被整体提升时,绿藻进行繁殖的能力就更强一些。
3.2无机碳对蓝绿藻碳利用机制及种群竞争的影响
在水体无机碳浓度升高的背景下,藻类的竞争并不局限于传统的氮磷资源型竞争和干扰型竞争,无机碳作为一个传统藻类生长限制性因素,会极大影响蓝绿藻的CCM机制,而反映在不同种类的藻类上面效应并不相同。这种差异性改变了浮游藻类的竞争优势,进而影响浮游藻类的群落结构。同时,无机碳水平升高还会影响藻类的化学元素组成,影响藻类的营养结构。
从水华的产生机理看,当水体处于无机碳限制的情况下,蓝藻更易于生长,成为优势种,并进一步演替为水华。水体pH较高的碱性系统易于捕获大气中的CO2,有利于藻类进行光合作用,因而较高的生产力往往出现在偏碱性水体中。尤其是在水华暴发阶段,处于碱性环境的铜绿微囊藻是生长最旺盛的,它可以保持旺盛的光合作用,而且诸多有关蓝绿藻生长代谢的论文表明蓝绿藻在对数生长期时会不断代谢出碱性物质,使水体pH持续升高并稳定在较高水平,因为此时它的无机碳亲和力也较高(支彦丽等,2008b;徐涛等,2007;杨波等,2007)。此时,蓝藻等因其CCM机制较绿藻更高效,能够直接利用碳酸氢盐的藻类在水体中低浓度CO2的生长会明显增快,引起淡水中藻类群落的迅速演替,一跃成为优势种属,引发水华。这是因为蓝藻对于无机碳亲和力K0.5很高,在局部形成细胞高密度造成环境无机碳含量骤减情况下,也可以保证微囊藻能够正常生长以及保持其优势种的地位(徐涛等,2007)。反之,当无机碳水平被整体提升时,尤其是以CO2形式加入系统时,绿藻进行繁殖的能力就更强一些,并且水华的危害会被降低。当蓝绿藻CCM机制减弱时,原本CCM机制的藻类和其他不具备CCM机制的浮游植物可能在生态系统中获得新的增长优势,蓝绿藻之间以及与其他浮游生物的竞争关系将会大幅改变,由此水华的出现频率和强度可能减少。当水体中溶解CO2浓度升高,随之碳酸氢盐碱度升高。这时绿藻则会快速繁殖并成为优势藻种,同时在蓝藻内部,不产藻毒素的种类生长情况更优于产生藻毒素的那些物种(Yu等,2015)。因此,只有在适宜的碱度条件下,水体中藻类多样性才能达到最大。已进行的实验说明无机碳浓度的升高对于不同藻类组合有不同的结果,目前总体来说是不定向的。这一方面仍需要更多的原位实验来进一步证明(O’Neil et al.,2012)。
关于藻类的化学元素组成带给竞争的影响,传统理论认为,藻类的化学组成相对固定,由Redfield确定,藻类的C∶N∶P比值在106∶16∶1附近。但近些年的研究却表明浮游植物的化学元素组成受水体DIC含量影响较大,大气CO2水平的上升将会显著改变浮游植物的化学元素组成,这就打破了Redfield比值不受外界影响的传统观点。在大气CO2体积分数从380×10-6升至750×10-6后,在淡水湖泊中绿藻细胞的碳、氮元素组成在CO2水平升高后也发生了显著地提高,同时磷元素的构成并无明显变化(Sterner等,2002)。现阶段淡水湖泊富营养化大多处于磷元素限制的情况,而无机碳的升高却一定程度上使得水华磷限制的作用有所削弱,整体系统的正反馈作用加强。在众多藻类中,个别种属的硅藻成为例外,其C∶N比随着无机碳水平升高呈现逐渐降低的现象,从侧面解释了蓝绿藻在富营养化湖泊中优势扩大的现象。
4 研究展望
综上,全球大气中CO2浓度升高势必通过水气平衡影响水体中无机碳的形态转化和浓度水平,而无机碳对淡水环境中蓝绿藻的生长及竞争研究相对薄弱,目前,有限的淡水资源水质恶化严重,在全球气候变化的大背景下,更加迫切需要开展以下研究。
(1)在无机碳水平升高的情况下,蓝绿藻CCM机制发生怎样的变化,从细胞水平上无机碳的利用机制需进一步研究。
(2)在C、N、P元素都很丰富的条件下,蓝绿藻的种群竞争及其代谢机制会如何变化是很好的研究方向。
(3)全球CO2水平升高带来不同形态无机碳的变化,相关联的水文水质特征发生的变化对蓝绿藻的生长及种群竞争需要更多的研究来验证。
参考文献:
ANDERSEN T, F Ø ANDERSEN. 2006. Effects of CO2concentration on growth of filamentous algae and Littorella uniflora in a Danish softwater lake [J]. Aquatic Botany, 84(3): 267-271.
BHARTI R K, S SRIVASTAVA, I S THAKUR. 2014. Isolation, purification, characterization and mass spectroscopic analysis of carbonic anhydrase from Serratia sp. for sequestration of carbon dioxide and formation of calcite [J]. Journal of Environmental Chemical Engineering, 2(1): 31-39.
BJÖRN ROST, ULF RIEBESELL, DIETER SÜLTEMEYER. 2006. Carbon acquisition of marine phytoplankton: Effect of photoperiod length [J]. Limnology and Oceanography, 51(1): 12-20.
BRILMAN W, L GARCIA ALBA, R VENEMAN. 2013. Capturing atmospheric CO2using supported amine sorbents for microalgae cultivation [J]. Biomass and Bioenergy, 53: 39-47.
ELLIOTT J A. 2012. Is the future blue-green? A review of the current model predictions of how climate change could affect pelagic freshwater cyanobacteria [J]. Water Research, 46(5): 1364-1371.
EMELKO M B, U SILINS, K D BLADON, et al. 2011. Implications of land disturbance on drinking water treatability in a changing climate: Demonstrating the need for “source water supply and protection”strategies [J]. Water Research, 45(2): 461-472.
FINLAY K, R J VOGT, M J BOGARDET, et al. 2015. Decrease in CO2efflux from northern hardwater lakes with increasing atmospheric warming [J]. Nature, 519(7542): 215-218.
GHOSHAL D, H DAVID HUSIC, A GOYAL. 2002. Dissolved inorganic carbon concentration mechanism in Chlamydomonas moewusii [J]. Plant Physiology and Biochemistry, 40(4): 299-305.
HARGRAVE C W, GARY K P, ROSADO S K. 2009. Potential effects of elevated atmospheric carbon dioxide on benthic autotrophs and consumers in stream ecosystems: a test using experimental stream mesocosms [J]. Global Change Biology, 15(11): 2779-2790.
HU Y B, LIU C Y, YANG G P, et al. 2015. The response of the carbonate system to a green algal bloom during the post-bloom period in the southern Yellow Sea [J]. Continental Shelf Research, 94: 1-7.
IZUMO A, S FUJIWARA, Y OYAMA, et al. 2007. hysicochemical properties of starch in Chlorella change depending on the CO2concentration during growth: Comparison of structure and properties of pyrenoid and stroma starch [J]. Plant Science, 172(6): 1138-1147.
JAISWAL P, A K KASHYAP. 2002. Isolation and characterization of mutants of two diazotrophic cyanobacteria tolerant to high concentrations of inorganic carbon [J]. Microbiological Research, 157(2): 83-92.
LOW-DÉCARIE E, G F FUSSMANN, G BELL. 2014. Aquatic primary production in a high- CO2world [J]. Trends in Ecology & Evolution, 29(4): 223-232.
MOAZAMI-GOUDARZI M, B COLMAN. 2012. Changes in carbon uptake mechanisms in two green marine algae by reduced seawater pH [J]. Journal of Experimental Marine Biology and Ecology, 413: 94-99.
O’NEIL J M, T W DAVIS, M A BURFORD, et al. 2012.The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change [J]. Harmful Algae, 14: 313-334.
PAERL H W, V J PAUL. 2012. Climate change: Links to global expansion of harmful cyanobacteria [J]. Water Research, 46(5): 1349-1363.
PIERANGELINI, M., S. STOJKOVIC, P.T. ORR et al., Elevated CO2 causes changes in the photosynthetic apparatus of a toxic cyanobacterium, Cylindrospermopsis raciborskii[J]. Journal of Plant Physiology, 2014. 171(12): 1091-1098.
RAVEN J A, J BEARDALL. 2014. CO2concentrating mechanisms and environmental change [J]. Aquatic Botany, 118(0): 24-37.
REICHWALDT E S, A GHADOUANI. 2012. Effects of rainfall patterns on toxic cyanobacterial blooms in a changing climate: Between simplistic scenarios and complex dynamics [J]. Water Research, 46(5): 1372-1393.
ROSEMOND A D, J P BENSTEAD, P M BUMPERSET. et al. 2015. Freshwater ecology. Experimental nutrient additions accelerate terrestrial carbon loss from stream ecosystems [J]. Science (New York, N.Y.), 347(6226): 1142-5.
SINGH S P, P SINGH. 2014. Effect of CO2concentration on algal growth: A review [J]. Renewable and Sustainable Energy Reviews, 38: 172-179.
SINHA R, L A PEARSON, T W DAVIS, et al. 2012. Increased incidence of Cylindrospermopsis raciborskii in temperate zones – Is climate change responsible? [J]. Water Research, 46(5): 1408-1419.
STERNER R W, ELSER J J, VITOUSEK P. 2002. Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere [M]. Princeton: Princeton University Press: 24-78.
SU M, YU J, ZHANG J. et al. 2015. MIB-producing cyanobacteria (Planktothrix sp.) in a drinking water reservoir: Distribution and odor producing potential [J]. Water Research, 68: 444-453.
TANG R, J M CLARK, T BOND, et al. 2013. Assessment of potential climate change impacts on peatland dissolved organic carbon release and drinking water treatment from laboratory experiments [J]. Environmental Pollution, 173: 270-277.
WINCK F V, D O PÁEZ MELO, A F GONZÁLEZ BARRIOS. 2013. Carbon acquisition and accumulation in microalgae Chlamydomonas: Insights from “omics” approaches [J]. Journal of Proteomics, 94: 207-218.
YU L, KONG F, SHI X, et al. 2015. Effects of elevated CO2on dynamics of microcystin-producing and non-microcystin-producing strains during Microcystis blooms [J]. Journal of Environmental Sciences, 27: 251-258.
ZHANG M, DUAN H, SHI X, et al. 2012. Contributions of meteorology to the phenology of cyanobacterial blooms: Implications for future climate change[J]. Water Research, 46(2): 442-452.
陈雄文, 高坤山. 2003. CO2浓度对中肋骨条藻的光合无机碳吸收和胞外碳酸酐酶活性的影响[J]. 科学通报, 48(21): 2275-2279.
成慧敏. 2006. 两种模式蓝藻CCM的运转及其对过剩光能的耗散作用[D]. 华中师范大学.
程丽华, 张林, 陈欢林, 等. 2005. 微藻固定CO2研究进展[J]. 生物工程学报, 21(2): 177-181.
戴树桂. 2006. 环境化学[M]. 北京: 高等教育出版社.
李小梅. 2012. 氮磷比与光、二氧化碳对三角褐指藻光合无机碳利用的交互作用[D]. 广州大学.
李鑫, 胡洪营, 张玉平. 2011. 无机碳源对栅藻LX1生长及油脂积累特性的影响[J]. 环境科学, 32(8): 2260-2266.
(美)J.J.摩尔根, 瑞.W.斯. 1987. 水化学-天然水体化学平衡导论[M]. 科学出版社.
孙凌, 阚元卿, 张冬梅, 等. 2007. 无机碳对浮游藻类生长和群落结构的影响[J]. 环境污染与防治, 29(5): 352-356.
王思莹, 张君枝, 马文林. 2015. 超富营养水平下初始碱度对铜绿微囊藻生长影响的研究[J]. 绿色科技, (2): 203-206.
王玮蔚, 孙雪, 王冬梅, 等. 2014. 盐度和无机碳对蛋白核小球藻生长、胞外碳酸酐酶活性及其基因表达的影响[J]. 水产学报, 38(7):920-928.
夏建荣, 高坤山. 2002a. CO2浓度升高对斜生栅藻生长和光合作用的影响[J]. 植物生理学通讯, 38(5): 431-433.
夏建荣, 高坤山. 2002b. 绿藻CO2浓缩机制的研究进展[J]. 应用生态学报, 13(11): 1507-1510.
夏建荣. 2006. 大气CO2浓度升高对海洋浮游植物影响的研究进展[J].湛江海洋大学学报, 26(3): 106-110.
徐涛, 宋立荣. 2007. 三株铜绿微囊藻对外源无机碳利用的研究[J]. 水生生物学报, 31(2): 245-250.
杨波, 储昭升, 金相灿, 等. 2007. CO2/pH对三种藻生长及光合作用的影响[J]. 中国环境科学, 27(1): 54-57.
姚臣谌, 张靖, 吴以赢, 等. 2012. 三峡坝区干流及香溪河库湾水体秋季二氧化碳分压[J]. 生态学杂志, 31(1): 152-157.
喻元秀, 汪福顺, 王宝利, 等. 2009. 溶解无机碳及其同位素组成特征对初期水库过程的响应——以新建水库(洪家渡)为例[J]. 矿物学报, 29(2): 268-274.
张宝燕, 田平芳. 2014. 羧酶体结构及其CO2浓缩机制研究进展[J]. 生物工程学报, 30(8): 1164-1171.
赵天宏, 王美玉, 张巍巍, 等. 2006. 大气CO2浓度升高对植物光合作用的影响[J]. 生态环境, 15(5): 1096-1100.
赵旭辉, 孔繁翔, 谢薇薇, 等. 2012. 全球CO2水平升高对浮游植物生理和生态影响的研究进展[J]. 生态学报, 32(21): 6880-6891.
赵旭辉, 汤龙升, 史小丽, 等. 2013. 模拟大气CO2水平升高对春季太湖浮游植物生理特性的影响[J]. 环境科学, 34(6): 2126-2133.
支彦丽, 储昭升, 钟远, 等. 2008b. 铜绿微囊藻和四尾栅藻光合利用碳酸氢盐探讨[J]. 湖泊科学, 20(4): 443-449.
支彦丽, 金相灿, 钟远, 等. 2008a. 淡水绿藻的光合碳酸氢盐利用探讨[J]. 环境科学学报, 28(8): 1519-1525.
邹定辉, 高坤山, 阮祚禧. 2001. 高CO2浓度对石莼光合作用及营养盐吸收的影响[J]. 青岛海洋大学学报(自然科学版), 31(6): 877-882.
A Review on the Effect of Inorganic Carbon on Blue-green Algae Growth
ZHANG Junzhi1, WANG Qi1, MA Wenlin1*, CHEN Han1, ZHAO Yuanling1, ZHANG Yanna2
1. Beijing Climate Change Response Research and Education Center//Beijing University of Civil Engineering and Architecture, Beijing 100044, China; 2. Research Institute of Petroleum Exploration and Development, Beijing 100083, China
Abstract:The blue-green algae is one kind of the important components of phytoplankton in waterbodies. It is also the main primary producer of freshwater and marine ecosystems. In recent years, with the increase of CO2emission to atmosphere, the global climate is changing significantly. The frequent extreme weather events exert great impact on the concentration of dissolved organic carbon in waterbodies, as a consequence, CO2-concentrating mechanism (CCM) of algae is affected. Through CCM, the concentration of CO2could be increased located at RubisCO of the algae cells, in order to promote the photosynthesis and diminish light respiration of algae. Based on the climate change due to the increase of CO2concentration, the equilibrium among inorganic carbon in different forms in open waterbodies was first described in this review. The carbon recycling processes in waterbodies and their effect on the algae was then analyzed. Various experimental simulation technology of the elevated inorganic carbon concentration in the environment as well as the research method of the algae growth and species competition with the elevated CO2concentration were also summarized.According to the researches on the CCM of blue-green algae, inoraganic carbon enter the blue-green algae via the Calvin Cycle. Under the catalysis of carbonic anhydrase, HCOand COas the form of inorganic carbon could be transformed into CO2, which increases the CO2concentration around RubisCO and the inorganic carbon utilization efficiency of algae. However, CCM of algae is also affected by pH, illumination, nitrogen and phosphorus, etc. Based on this theory, the effect of inorganic carbon on the cell structure, size and the individual cell proliferation of blue-green algae was summarized.Some researchers indicated that CO2with high concentration could promote the growth of nuclear protein of algae cells, and this phenomenon is much significant for the algae cells with small size. Since the CO2could be directly utilazed by blue-green algae, CO2play a more important role on promoting the reproduction of individual algal cell proliferation. The pH of natural surface water is in the range of neutral to weak alkaline. Therefore, there are more researchers focus on the CO2, HCOutilization by blue-green algae and their CCM. In summary, although there are a great number of studies on the effect of climate change, the factors such as the mechanisms of carbon utilization, the micro-changes in the cell and the CCM of blue-green algae during the algal growth and bloom still need to be further studied.
Key words:climate change, inorganic carbon, blue-green algae, CO2-concentrating mechanism
收稿日期:2015-03-23
*通信作者:马文林,E-mail: mawenlin@bucea.edu.c
作者简介:张君枝(1979年生),女,讲师,博士,主要从事气候变化和水质安全保障相关研究。E-mail: zhangjunzhi@bucea.edu.cn
基金项目:国家自然科学基金青年科学基金项目(51408022);北京市优秀人才培养资助项目(2013D005017000009)
中图分类号:X17
文献标志码:A
文章编号:1674-5906(2015)07-1245-08
DOI:10.16258/j.cnki.1674-5906.2015.07.025
