高山森林不同类型粗木质残体腐殖化特征
2015-03-12刘辉杨万勤倪祥银肖洒吴福忠
刘辉,杨万勤,倪祥银,肖洒,吴福忠
四川农业大学生态林业研究所//林业生态工程重点实验室//高山森林生态系统定位研究站,四川 成都 611130
高山森林不同类型粗木质残体腐殖化特征
刘辉,杨万勤,倪祥银,肖洒,吴福忠*
四川农业大学生态林业研究所//林业生态工程重点实验室//高山森林生态系统定位研究站,四川 成都 611130
摘要:高山森林粗木质残体腐殖化是促进生态系统土壤发育和碳吸存的主要途径之一,并可能受到腐烂等级和木质残体类型的影响,但一直缺乏必要的关注。因此,在2013年8月,以青藏高原东缘海拔3 600 m左右的高山森林中具有代表性的岷江冷杉(Abies faxoniana)原始林为研究对象,根据区域内坡度和物种组成设置3个100 m×100 m的典型样地,在研究样地内随机选取5个腐烂等级的岷江冷杉粗木质残体,每个腐烂等级3株,4种粗木质残体,共计60株,调查和研究区域内不同腐烂等级倒木、枯立木、根桩和大枯枝等主要类型粗木质残体的腐殖化特征。结果表明:枯立木、大枯枝和倒木腐殖质碳、胡敏酸碳和富里酸碳含量随腐烂等级增加而增大,而根桩腐殖质碳、胡敏酸碳和富里酸碳含量随腐烂等级增加而减小,并且枯立木在各个分解阶段腐殖质碳、胡敏酸碳和富里酸碳含量均保持较高水平。同时,枯立木、大枯枝和倒木的腐殖化度随腐烂等级增加呈增大趋势,根桩腐殖化度却随着腐烂等级增加而减小,其腐殖化度分别为52.93%~85.88%、49.2%~73.68%、54.94%~67.21%和53.41%~68.68%。枯立木腐殖化度明显高于其他3种类型粗木质残体,且在腐殖化后期(腐烂Ⅳ级或Ⅴ级)腐殖化度最大。这对进一步认识高山森林生态系统植物-土壤互作过程具有重要意义。
关键词:高山森林;粗木质残体;腐烂等级;腐殖化
引用格式:刘辉,杨万勤,倪祥银,肖洒,吴福忠. 高山森林不同类型粗木质残体腐殖化特征[J]. 生态环境学报, 2015, 24(7): 1143-1149.
LIU Hui, YANG Wanqin, NI Xiangyin, XIAO Sa, WU Fuzhong. Characters of Different Type of Coarse Woody Debris Humification in An Alpine Forest [J]. Ecology and Environmental Sciences, 2015, 24(7): 1143-1149.
粗木质残体(coarse woody debris)广泛存在于高山森林地表,主要包括倒木、枯立木、大枯枝等,是森林生态系统结构和功能的重要组成,在森林群落更新、维持森林系统完整性和稳定性以及能量流动等方面具有十分重要的作用(侯平等,2001;何东进等,2009;Harmon et al.,1986)。受低温、频繁地质灾害以及极端气候影响,使得高山土壤发育受阻,土壤土层结构不完善(Yang et al.,2005)。粗木质残体腐殖化对增加土壤有机质,促进土壤发育,和维持生态系统物质循环和平衡具有重要意义(Lal,2005;Ponge,2013)。然而由于立地环境、分解者和自身质量的影响,不同类型粗木质残体腐殖化在不同腐烂等级下可能存在较大差异(侯平等,2001)。已有研究发现,相对于立于地上的枯立木和悬于树干的大枯枝,倒木和根桩可以更好地受到土壤生物类群的作用,且土壤表面相对稳定的温湿度可以一定程度上维持较好的生物生存环境,使得倒木和根桩腐殖化程度更高、腐殖化进程更快(Yatskov et al.,2007)。同时,由于生态系统中粗木质残体类型多样,腐殖化过程差异较大,其对高山森林土壤的形成具有重要意义,但缺乏必要关注。
位于青藏高原东缘和长江上游的川西高山森林,对区域气候调节、涵养水源和维持生物多样性等方面发挥特殊效应(Yang et al.,2005)。现有研究更加注重的是高山森林生态系统粗木质残体生物量以及分解过程中微生物群落的变化特征(肖洒等,2014;常春晖等,2014),但对其腐殖化过程以及与土壤形成之间的关系研究非常的缺乏。因此,我们基于本团队前期的研究结果基础上,以高山岷江冷杉(Abies faxoniana)原始林为研究对象,进一步研究其地表普遍存在的倒木、大枯枝、根桩和枯立木等不同类型粗木质残体在不同腐烂等级的腐殖化特征,为进一步研究高山森林生态系统土壤发育以及水土保持提供基础资料,为深入了解川西高山森林生态系统粗木质残体腐殖化过程以及对森林生态系统的可持续管理提供一定的理论依据。
1 材料和方法
1.1研究区域概况
研究区域位于四川省阿坝藏族羌族自治州理县毕棚沟风景区(31°14′~31°19′N,102°53′~102°57′E,海拔2458~4619 m),地处青藏高原东缘与四川盆地过渡的高山峡谷地带。该区域年平均气温2~4 ℃,1月和7月平均温度分别为-8.67和9.53 ℃,年降水量850 mm左右,绝大数的降水集中在5─8月。每年11月至次年4月发生土壤季节性冻融,每年11月下旬开始降雪,12月下旬至次年3月初完全雪被覆盖,直至4月开始融化(谭波等,2011)。区域典型的优势乔木有红桦(Betula albo-sinensis)、岷江冷杉(Abies faxoniana)、川西云杉(Picea balfouriana)等,灌木有康定柳(Salix paraplesia)、高山杜鹃(Salix paraplesia)、华西箭竹(Fargesia nitida)等(杨万勤等,2007)。土壤有机层有机碳、全磷、全氮含量分别为(150.3±15.9)、(1.2±0.2)、(9.7±0.9) g·kg-1(肖洒等,2014),腐殖质碳和腐殖化度分别为(97.25±0.88) g·kg-1和(61.10%±6.21%)(倪祥银等,2014)。该区域土壤浅薄、发育缓慢,为发育于坡积物上的暗棕壤(Wu et al.,2010)。
1.2研究方法
本次所采集的样品均是直径≥10 cm的粗木质残体(CWD)。根据粗木质残体形态和尺度大小进行分类,据其在森林生态系统中的位置和状态,粗木质残体又被进一步分为倒木(Fallen log)、枯立木(Snag)、根桩(Stump)和大枯枝(Large branch)。为了区分倒木和枯立木,枯立木指的是倾斜度(偏垂直方向)不超过45°,粗头直径≥10 cm的死木质物,长度>1 m的;与枯立木其他特征相似,将长度<1 m的木质残体定义为根桩(闫恩荣等,2005;Harmon et al.,1986)。
通过对研究区域内粗木质残体的物种组成与储量以及其分布特征,土壤类型、坡度、坡向以及郁闭度等因素的调查研究后,在2013年8月以高山森林海拔3600 m左右具有代表性的岷江冷杉原始林为研究对象,设置3个100 m×100 m的典型样地,按照闫恩荣等(2005)提出的粗木质残体5级腐烂系统的划分标准(I级:调查的死木质物新鲜,死不足1年;II级:开始腐解,刀片可刺进几毫米;III级:刀片可刺进约2 cm;IV级:刀片可刺进2~5 cm;V级:刀片可随意刺穿粗死木的木质体),在样地内随机选取5个腐烂等级的岷江冷杉粗木质残体,每个腐烂等级3株,4种粗木质残体,共计60株,去除粗木质残体表面泥土、苔藓、石块等附生物质做好标记后装入无菌的封口袋低温保存,带回实验室进行实验分析。
1.2.1胡敏酸和富里酸
胡敏酸、富里酸的提取和分离参考《中华人民共和国林业行业标准LY/T 1238─1999》。称取风干样品0.500 g于150 mL锥形瓶,加100 mL 0.1 mol·L-1NaOH和0.1 mol·L-1Na4P2O7·10 H2O混合提取液,加塞振荡10 min,沸水浴1 h,待冷却后过滤,将过滤液进行两部分处理:一方面,过0.45 μm滤膜,滤液为浸提液。取浸提液1 mL于PE管,稀释10倍,测定腐殖质碳(humus carbon,HC);另一方面,取浸提液20 mL于试管,加热至沸,取出,逐滴加0.5 mol·L-1H2SO4至pH=2(絮状沉淀),于80 ℃水浴30 min。用0.05 mol·L-1H2SO4洗涤,过滤,沉淀即为胡敏酸。用热的0.05 mol·L-1NaOH少量多次洗涤沉淀,过滤至100 mL容量瓶,定容,取溶解的胡敏酸溶液4 mL于离心管中,稀释10倍,测定胡敏酸碳(humic acid carbon,HAC)(Ni et al.,2015;Wang et al.,2010)。使用TOC(multi N/C 2100,analytic jena,Germany)测定腐殖质碳、胡敏酸碳,由此计算富里酸碳(fulvic acid carbon,FAC)以及胡敏酸碳和富里酸碳比值。
富里酸碳(fulvic acid carbon,FAC)=腐殖质碳-胡敏酸碳
1.2.2△logK和E4/E6
根据碱提取液在465和665 nm吸光度比值以及在400和600 nm时吸光度对数值表征粗木质残体的腐殖化度,即E4/E6和△logK。E4/E6和△logK反映的是腐殖酸分子组成和结构的变化,即粗木质残体腐殖化度变化。E4/E6和△logK值越大,其腐殖酸分子芳构化度和分子量越小,导致腐殖化度减小(窦森等,2007)。
称取风干样品0.100 g于150 mL锥形瓶中,加100 mL 0.1 mol·L-1NaOH和0.1 mol·L-1Na4P2O7·10 H2O混合提取液,加塞震荡10 min,100 ℃下水浴30 min,待冷却后,将提取液过0.45 μm滤膜,所得滤液即为待测液。使用紫外可见分光光度仪(TU-1901,Puxi,Beijing,China)测定。E4/E6和△logK值的计算方法如下(Ni et al.,2014):
△logK=log(A400/A600)
E4/E6=A465/A665
A400、A465、A600、A665分别是波长为400、465、600、665 nm测定的吸光度。
1.3数据处理和统计分析
腐殖化度(humIfication degree,HD)的计算公式(Gigliotti et al.,1999):
HD(%)=CHC/COC×100%
式中,CHC为腐殖质碳含量,COC为有机碳含量。
数据运用Excel 2007进行数据整理与绘图、SPSS 20.0(IBM SPSS Statistics Inc.,Chicago,IL,USA)进行方差分析。用单因素方差分析(one-way ANOVA)和最小显著差异法(Least significant difference,LSD)检验不同腐烂等级下的不同类型粗木质残体腐殖化过程中胡敏酸碳、富里酸碳、胡敏酸碳/富里酸碳、E4/E6和△logK值以及腐殖化度的差异显著性。用双因素方差分析(two-way ANOVA)检测腐烂等级、粗木质残体类型对腐殖质碳、胡敏酸碳、富里酸碳、胡敏酸碳/富里酸碳、E4/E6、△logK以及腐殖化度影响。显著性水平设为P=0.05。数值以平均值±标准误差(mean±SE)表示。
2 结果
2.1腐殖质碳、胡敏酸碳和富里酸碳含量
高山森林不同腐烂等级显著影响粗木质残体在腐殖化过程中腐殖质碳、胡敏酸碳、富里酸碳含量(P<0.05),且4种粗木质残体之间腐殖质碳表现出极显著类型差异(P<0.01),胡敏酸碳和富里酸碳含量表现出显著(P<0.05)类型差异(表1)。不同类型粗木质残体腐殖化过程中腐殖质碳含量、胡敏酸碳含量以及富里酸碳含量随腐烂程度变化基本一致。具体表现为:倒木、枯立木腐殖质碳含量、胡敏酸碳和富里酸碳含量在腐殖化前期(腐烂Ⅰ级或Ⅱ级)最低,整体呈现随腐烂等级增加大体呈增加趋势;根桩腐殖质碳含量、胡敏酸碳和富里酸碳含量整体出现降低;大枯枝在腐殖化过程各物质碳含量出现先降后升的变化并在腐烂Ⅲ级有明显增大。腐烂Ⅰ级时4种类型粗木质残体腐殖质碳含量根桩>枯立木>大枯枝>倒木,腐烂Ⅴ级时不同类型粗木质残体腐殖质碳含量枯立木最大,大枯枝和倒木次之,倒木的含量最小。与其他类型的粗木质残体相比,枯立木腐殖质碳含量在各个分解阶段均表现出较高水平的(图1)。
2.2胡敏酸碳和富里酸碳比值
高山森林不同腐烂等级和类型对粗木质残体腐殖化过程中胡敏酸碳/富里酸碳无显著影响(P>0.05)(表1)。倒木和枯立木腐殖化过程胡敏酸碳/富里酸碳表现出随腐烂程度加深而降低的趋势;根桩腐殖化过程中胡敏酸碳/富里酸碳呈现先增后减的变化,在腐烂Ⅳ级呈显著降低;大枯枝胡敏酸碳/富里酸碳呈现先降低后增加的趋势,并且在腐烂Ⅲ级时出现明显升高,但粗木质残体在各个分解阶段胡敏酸碳/富里酸碳总体上均大于1。在腐烂Ⅰ级时,不同类型粗木质残体胡敏酸碳/富里酸碳值枯立木>倒木>大枯枝>根桩,腐烂Ⅴ级时,根桩胡敏酸碳/富里酸碳增加程度最大。与其他类型粗木质残体相比,大枯枝在各个分解阶段均表现出较高的胡敏酸碳和富里酸碳比值(图2)。

表1 不同腐烂等级(decay class),不同类型粗木质残体(coarse woody debris)对腐殖质碳(HC)、胡敏酸碳(HAC)、富里酸碳(FAC)、胡敏酸碳/富里酸碳、△logK、E4/E6和腐殖化度(HD)F值双因素方差分析Table 1 F values for Two-way ANOVA results for the effect of decay class, coarse woody debris and their interactions on humus carbon, humic acid carbon, fulvic acid carbon, humic acid carbon/fulvic acid carbon, △logK, E4/E6and humification degree
2.3△logK
高山森林不同类型粗木质残体对腐殖化过程中△logK值无显著影响(P>0.05),而不同腐烂等级极显著影响粗木质残体在腐殖化过程中△logK值(P<0.01)(表1)。不同类型粗木质残体腐殖化过程中△logK值随腐烂程度加深,倒木和根桩整体呈下降趋势,在腐烂Ⅱ级或腐烂Ⅳ级△logK值最低;而枯立木和大枯枝表现出先降后升的趋势变化,△logK值在腐烂Ⅳ级下的明显增加(图3)。
2.4E4/E6
高山森林不同腐烂等级极显著影响粗木质残体在腐殖化过程中E4/E6值(P<0.01),且4种粗木质残体之间表现出极显著(P<0.01)的类型差异(表1)。在腐殖化过程中,E4/E6值的变化趋势与△logK值的一致。根桩和倒木腐殖化过程中E4/E6随腐烂程度加深而减小,在腐烂Ⅳ级时E4/E6值最小;枯立木和大枯枝E4/E6却随腐烂等级增加基本呈先降后升的趋势,在腐烂Ⅳ级时E4/E6值显著升高。整体看,根桩在腐烂Ⅳ级下的E4/E6值最小,而最大的E4/E6值出现在枯立木腐烂时。枯立木较其他3种粗木质残体,在不同分解阶段时的E4/E6值高于倒木、根桩和大枯枝(图4)。
2.5腐殖化度
高山森林不同腐烂等级极显著影响粗木质残体在腐殖化过程中腐殖化度(P<0.01),而4种粗木质残体之间表现出显著的(P<0.05)类型差异(表1)。倒木和枯立木在腐殖化过程中腐殖化度随腐烂等级变化呈现增大趋势;大枯枝腐殖化度的变化表现出先降后升的变化,在腐烂Ⅲ级增加最明显;而根桩腐殖化度随腐烂等级呈总体降低的变化,并在腐烂前期(腐烂Ⅱ级)最小。同时发现,除倒木外,其他类型粗木质残体腐殖化度在腐殖化后期(腐烂Ⅳ级或V级)达到最大。总体看,4种粗木质残体在各个分解阶段均保持较高的腐殖化度,枯立木的腐殖化度最高(52.93%~85.88%),根桩(49.20%~73.68%)次之,大枯枝(54.94%~67.21%)和倒木(53.41%~68.68%)的腐殖化度最低(图5)。
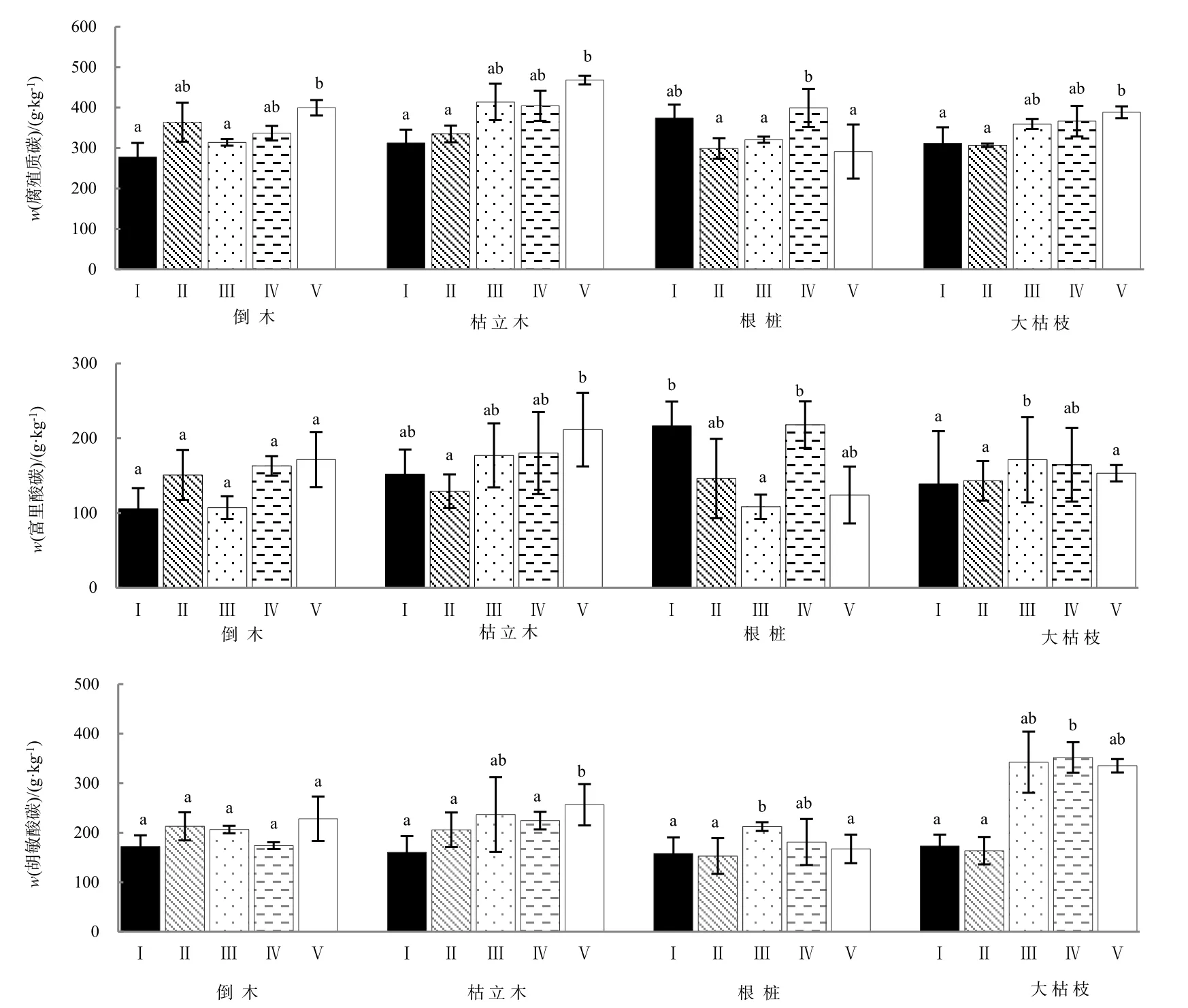
图1 不同腐烂等级粗木质残体腐殖质碳、胡敏酸碳和富里酸碳(平均值±标准误差,n=3)Fig. 1 Different decay class coarse woody debris humus carbon, humic acid carbon and fulvic acid carbon (mean±SE, n=3)
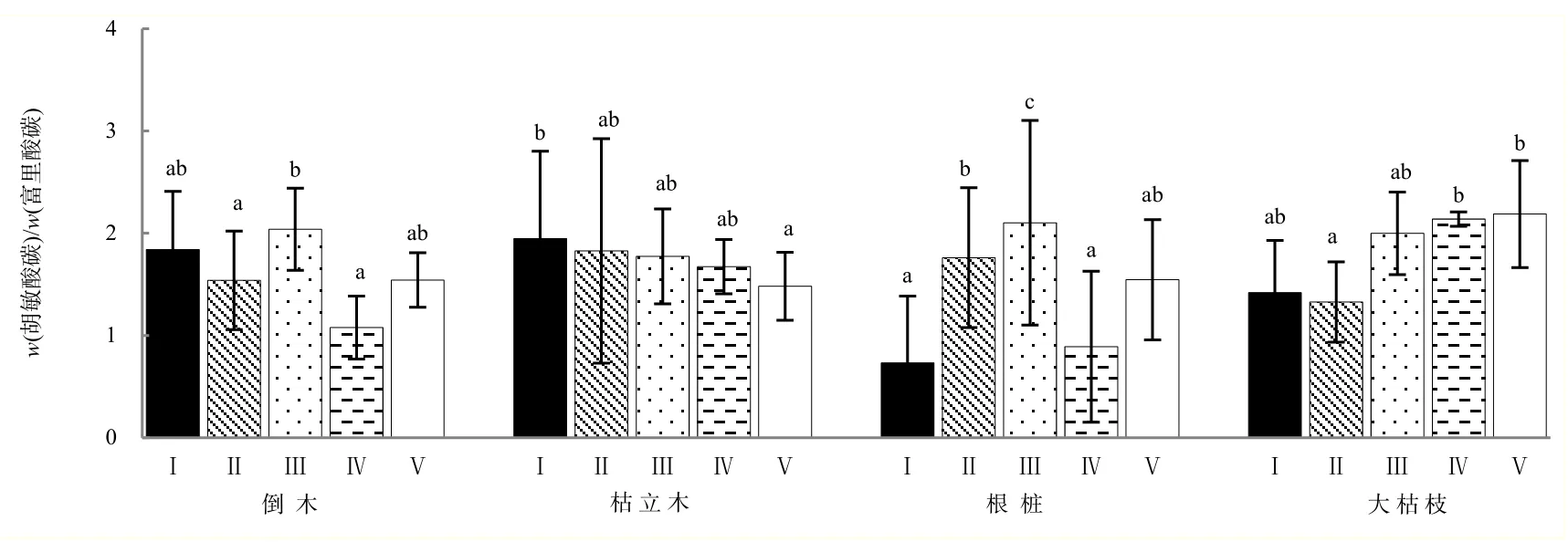
图2 不同腐烂等级粗木质残体胡敏酸碳/富里酸碳(平均值±标准误差,n=3)Fig. 2 Different decay class coarse woody debris humic acid carbon/fulvic acid carbon (mean±SE, n=3)
3 讨论
川西高山或亚高山森林生态系统常年遭受低温以及频繁地质灾害的影响,直接导致土壤形成和发育过程缓慢(杨万勤等,2007)。因此,地上部分粗木质残体腐殖化对于地表以及地下部分碳库输入和维持森林生态系统养分循环起着重要的作用。高山森林粗木质残体在低温的环境仍保持较高程度的腐殖化,但腐烂程度受粗木质残体类型影响(表1),这可能是不同类型粗木质残体受基质质量和立地环境的相互作用导致其腐烂程度不同,金光泽等(2009)也得出类似的结论。研究发现,粗木质残体腐殖质在各个分解阶段的腐殖化度存在一定差异,且随腐烂等级增加而增大。枯立木、根桩、大枯枝和倒木腐殖化度分别为52.93%~85.88%、49.2%~73.68%、54.94%~67.21%和53.41%~68.68%(图5),远远高于亚高山/高山森林凋落叶腐殖化度(倪祥银等,2014),说明高山森林广泛存在的粗木质残体在一定程度上可以平衡森林生态系统地上-地下部分碳库和养分库,这对增加土体肥力、促进高山土壤发育和加快土壤发生与演替过程具有十分重要的意义。
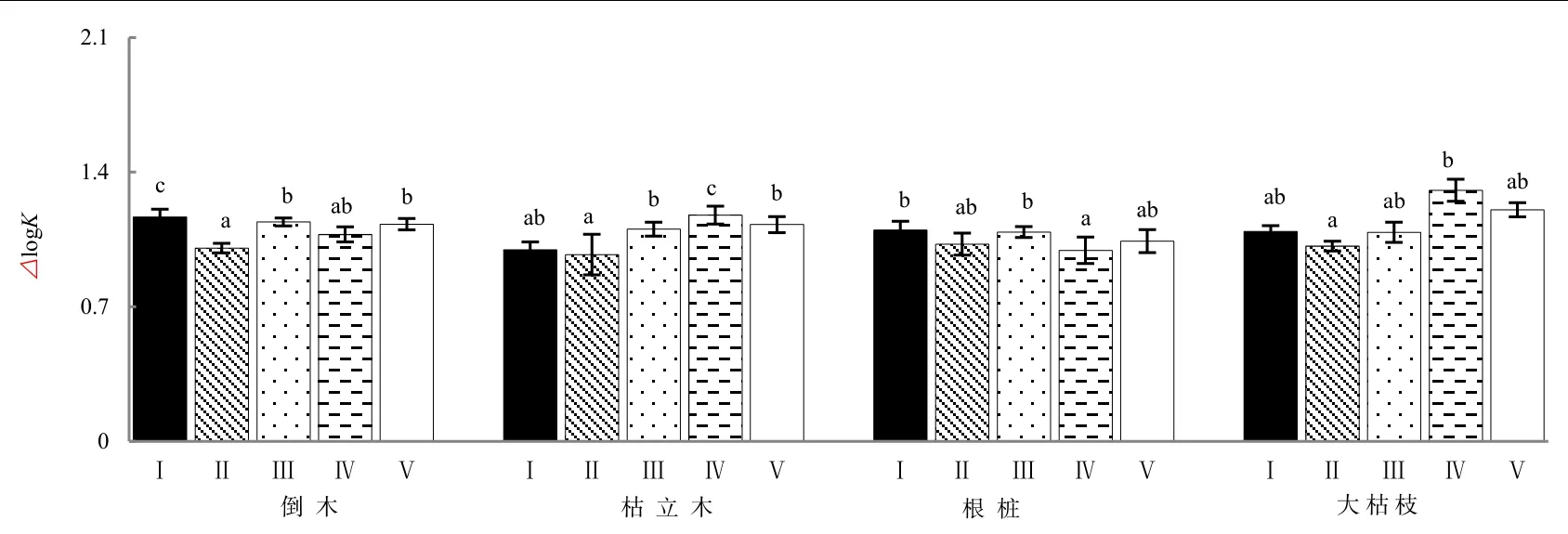
图3 不同腐烂等级粗木质残体△logK(平均值±标准误差,n=3)Fig. 3 Different decay class coarse woody debris △logK (mean±SE, n=3)
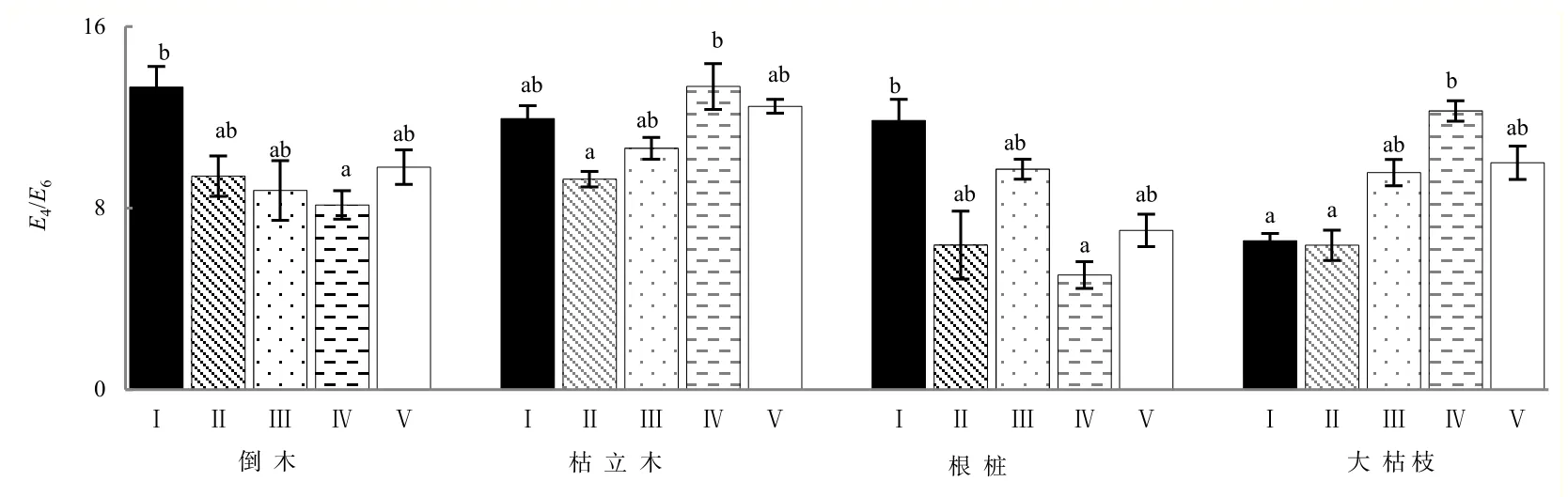
图4 不同腐烂等级粗木质残体E4/E6(平均值±标准误差,n=3)Fig. 4 Different decay class coarse woody debris E4/E6(mean±SE, n=3)

图5 不同腐烂等级粗木质残体腐殖化度(平均值±标准误差,n=3)Fig. 5 Different decay class coarse woody debris humification degree (mean±SE, n=3)
粗木质残体作为高山森林生态系统重要组成,需漫长时间才能完全分解,同时受频繁的地质灾害和极端天气影响,造成大量的粗木质残体残存于地表,其中充足的易分解成分为微生物提供了良好的底物有效性。本研究中,粗木质残体腐殖质碳、胡敏酸碳和富里酸碳,三者变化一致(图1)。总体看,4种粗木质残体在腐殖化过程,其腐殖质碳、胡敏酸碳、富里酸碳含量在各个分解阶段下存在差异,且到腐殖化后期变化缓慢,可能后期腐殖化受粗木质残体质量影响更显著,粗木质残体中大多数易分解组分在腐殖化后期均分解缓慢并累积更多难降解物质(Kreyling,2013),最后络合成腐殖质高聚物(Ponge,2013),这对微生物分解者生存、森林更新以及维持动植物和微生物的多样性具有特殊作用(唐旭利等,2005)。另外,在本研究中发现所有类型粗木质残体在各个分解阶段胡敏酸碳/富里酸碳均大于1(图2),出现这样的结果一方面是粗木质残体腐殖化过程中,低温和强烈的冻融循环抑制微生物参与形成富里酸的代谢途径,使得胡敏酸的形成速率要大于富里酸,这与窦森等(2007)长期的堆肥实验不一致;另外一方面粗木质残体在腐殖化过程中不同的分解阶段胡敏酸可能在粗木质残体产生之前已经形成。

表2 不同森林类型粗木质残体储存形式Table 2 Storage of coarse woody debris in different types of forest ecological system
森林粗木质残体腐殖化是物理、化学和生物因子相互作用的共同结果,为动、植物以及微生物提供良好生境。△logK和E4/E6这两个指标在一定程度上反映了粗木质残体在各个分解阶段的腐殖化的强弱程度(张晋京等,2004)。研究表明,根桩和倒木腐殖化过程E4/E6和△logK值随腐烂程度变化而减小,在腐烂Ⅳ级其腐殖化程度最大,根桩和倒木在腐殖化后期通透性增高(Mummey et al.,2002),空气的流通性加强,这种条件下利于好氧微生物的生长,因此腐殖化后期(腐烂Ⅳ级)腐殖化度要比腐殖化前期强,与常春晖等(2014)研究结果类似;大枯枝和枯立木腐殖化中E4/E6和△logK值变化与之相反,这与Yatskov et al.(2007)研究结论相符。大枯枝和枯立木存在的方式与根桩和倒木不同,立于空中,而大枯枝和枯立木相对根桩、倒木木质结构紧实,又不能充分与土壤接触,这在一定程度上阻碍了土壤微生物对大枯枝和根桩的腐解,减缓了大枯枝和枯立木的腐殖化进程,从而导致大枯枝和枯立木的腐殖化程度明显弱于根桩和倒木的。
综上所述,高山森林粗木质残体具有较高的腐殖化度,但腐殖化度受木质残体类型影响(表2),枯立木的腐殖化度最高,而倒木和大枯枝腐殖化度最低,且不同腐烂等级下粗木质残体腐殖化程度存在差异,在腐殖化后期(腐烂IV级或V级)其腐殖化度最高。同时,粗木质残体腐殖化过程中腐殖质碳、胡敏酸碳和富里酸碳含量表现出随腐烂程度加深而增大的趋势,并受腐烂等级和木质残体类型影响。总之,土壤发育受阻情境下,少有的土壤大多来自于地上部分粗木质残体腐殖化形成的腐殖物质,因此粗木质残体腐殖化过程对于调控和驱动植物-土壤互作过程以及碳吸存过程非常重要。本研究结果对认识高山森林生态系统地上部分-地下部分之间的互作过程以及高山森林可持续管理提供一定的理论基础。
4 结论
以高山冷杉原始林为研究对象,通过调查林内各类型粗木质残体在腐殖化过程中腐殖质碳、胡敏酸碳、富里酸碳、胡敏酸碳/富里酸碳、E4/E6、△logK、腐殖化度以及腐烂等级与他们之间的关系进行研究,得出以下重要结论:
(1)研究区域内枯立木、大枯枝和倒木腐殖质碳、胡敏酸碳和富里酸碳含量随腐烂等级增加而增大,而根桩腐殖质碳、胡敏酸碳和富里酸碳含量随腐烂等级增加而减小,并且枯立木在各个分解阶段腐殖质碳、胡敏酸碳和富里酸碳含量均保持较高水平。
(2)枯立木、大枯枝和倒木的腐殖化度随腐烂等级增加呈增大趋势,根桩腐殖化度却随着腐烂等级增加而减小,其腐殖化度分别为52.93%~85.88%、49.2%~73.68%、54.94%~67.21%和53.41%~68.68%。
(3)相对其他3种类型粗木质残体,枯立木腐殖化度最高,并在腐殖化后期(腐烂Ⅳ级或Ⅴ级)腐殖化度最大。
参考文献:
NI X Y, WU F Z, YANG W Q, et al. 2015. Accelerated foliar litter humification in forest gaps: Dual feedbacks of carbon sequestration during winter and the growing season in an alpine forest [J]. Geoderma, 241-242: 136-144.
YANG W Q, WANG K Y, KELLMAKI S. 2005. Litter dynamics of three subalpine forests in western Sichuan [J]. Pedosphere, 15(5): 653-659.
YATSKOV M, HARMON M E, KRANKINA O N. 2007. A chronosequence of wood decomposition in the boreal forest of Russia [J]. Canadian Journal of Forest Research, 33(17): 1211-1226.
NI X Y, WU F Z, YANG W Q, et al. 2014. The response of early foliar litter humification to reduced snow cover during winter in an alpine forest [J]. Canadian Journal of Soil Science, 94: 1-9.
LAL R. 2005. Forest soils and carbon sequestration [J]. Forest Ecology andManagement, 220(1-3): 242-258.
HARMON M E, FRANKIN J F, SWANSON F J, et al. 1986. Ecology of coarse woody debris in temperate ecosystems [J]. Advances in Ecological Research, 15: 133-302.
GIGLIOTTI G, BUSENLLI D, GIUSQUIANI P L. 1999. Composition changes of soil humus after massive application of urban waste compost: a comparison between FT-IR spectroscopy and humification parameters [J]. Nutrient Cycling in Agroe-cosystems, 55(1): 23-28.
PONGE J F. 2013. Plant-soil feedbacks mediated by humus forms: A review [J]. Soil Biology and Biochemistry, 57: 1048-1060.
KREYLING J. 2013. Snow removal reduces annual cellulese decomposition in a riparian boreal forest [J]. Canadian Journal of Soil Science, 93: 427-433.
WANG H, HONG Y T, LIN Q H, et al. 2010. Response of humification degree to monsoon climate during the Holocene from the Hong yuan peat bog, eastern [J]. Palaeogeography, 286(3-4): 171-177.
MUMMEY D L, PETER D, STAHL. 2002. Microbial biomarkers as an indicator of ecosystem recovery following surface mine reclamation [J]. Applied Soil Ecology, 21 (3): 251-259.
WU F Z, YANG W Q, ZHANG J, et al. 2010. Litter decomposition in two subalpine forests during the freeze-thaw season [J]. Acta Oecologica, 23(1): 135-140.
GODDERT VON OHEIMB, CHRISTINA WESTPHAL, WERNER HÄRDTLE. 2007. Diversity and spatio-temporal dynamics of dead wood in a temperate near-natural beech forest (Fagus Sylvatica) [J]. European Journal Forest Research, 126(3): 359-370.
GROVE S J. 2001. Extent and composition of dead wood in Australian lowland tropical rainforest with different management histories [J]. Forest Ecology and Management, 154: 35-53, 19.
杨万勤, 邓仁菊, 张健, 等. 2007. 森林凋落物分解及其对全球气候变化的响应[J]. 应用生态学报, 18(12): 2889-2895.
闫恩荣, 王希华, 黄建军, 等. 2005. 森林粗木质残体概念和及其分类[J].生态学报, 25(1):158-167.
侯平, 潘存德. 2001. 森林生态系统中粗木质残体及其功能[J]. 应用生态学报, 12(2): 309-314.
何东进, 何小娟, 洪伟, 等. 2009. 森林生态系统粗木质残体研究进展[J].林业科学研究, 22(5): 715-721.
常春晖, 吴福忠, 杨万勤, 等. 2014. 川西高山森林倒木的不同分解阶段的微生物群落特征变化[J]. 应用与环境生物学报, 20(6): 978-985.
倪祥银, 杨万勤, 吴福忠, 等. 2014. 雪被板块对高山森林凋落物腐殖化过程中胡敏酸和富里酸累积的影响[J]. 土壤学报, 51(5): 225-238.
肖洒, 杨万勤, 吴福忠, 等. 2014. 川西高山森林生态系统林下生物量及其随林窗的变化特征[J]. 生态环境学报, 23(9): 1515-1519.
窦森, 丁永强, 张晋京, 等. 2007. 不同CO2浓度对玉米秸秆分解期间土壤腐殖质形成的影响[J]. 土壤学报, 44(3): 458-466.
金光泽, 刘志理, 刘妍妍, 等. 2009. 小兴安岭谷地云冷杉林粗木质残体的研究[J]. 自然科学学报, 24(7): 1256-1266.
张晋京, 窦森. 2004. 土壤腐殖质分组研究[J]. 土壤通报, 35(6): 707-709.
唐旭利, 周国逸. 2005. 南亚热带典型森林演替类型粗死木质残体贮量及其对碳循环的潜在影响[J]. 植物生态学报, 29(4): 559-568.
谭波, 吴福忠, 杨万勤, 等. 2011. 雪被去除对川西高山森林冬季土壤温度及碳、氮、磷动态的影响[J]. 应用生态学报, 22(10): 2553-2559.
方江平, 项文化. 2008. 西藏色季拉山原始冷杉林生物量及分布规律[J].林业科学, 44(5):17-23.
刘妍妍, 金光泽. 2010. 小兴安岭阔叶红松林粗木质残体基础特征[J].林业科学, 44(4): 8-14.
王飞, 张秋良, 王冰, 等. 2013. 不同林龄草类-兴安落叶松林粗木质残体特征[J]. 东北林业大学学报, 41(5): 61-65.
Characters of Different Type of Coarse Woody Debris Humification in An Alpine Forest
LIU Hui, YANG Wanqin, NI Xiangyin, XIAO Sa, WU Fuzhong*
Long-term Research Station of Alpine Forest Ecosystem//Key Laboratory of Ecological Forestry Engineering//Institute of Ecology and Forestry, Sichuan Agricultural University, Chengdu 611130, China
Abstract:Humification of coarse woody debris was one of the main pathways of soil formation and carbon sequestration in alpine forest ecosystems, where type of coarse woody debris and decay class could play an important role in coarse woody debris humification thanks to its insulation effect. However, the results were still not clear which limit the understanding and response of coarse woody debris humification. Therefore, three 100 m×100 m plots were sampled in a Minjiang fir (Abies faxoniana) primary forest ecosystem around 3 600 m altitude in western Sichuan in August, 2013. In order to explore effects of type of coarse woody debris and different decay class, we investigated different decay class fallen logs, snags, stumps and large branches, each decay class of coarse woody debris and collected altogether 60 trees. Results clearly showed that: snag, fallen log and large branch of humus carbon, humic acid and fulvic acid carbon content increased with the change of decay levels, whereas snag humus carbon, humic acid and fulvic acid carbon content was maintaining decline. At the same time, snag, stump, large branch, fallen log, which humification degree respectively 52.93%~85.88%, 49.2%~73.68%, 54.94%~67.21% and 53.41%~68.68%. Snag, its humification degree was significantly higher than the other three types of coarse woody debris, and humification degree was the highest in the late humification (IV decay class or V decay class). For further understanding about the alpine forest ecosystem of plant and soil interaction process which is of great significance.
Key words:alpine forest; coarse woody debris; decay class; humification
收稿日期:2015-03-31
*通信作者:吴福忠(1981年生),男,博士,博士生导师,主要从事森林生态等研究。E-mail: wufzchina@163.com
作者简介:刘辉(1992年生),男,硕士研究生,主要从事森林生态学研究。E-mail: 1350737643@qq.com
基金项目:国家自然科学基金项目(31170423;31270498);国家“十二五”科技支撑计划(2011BAC09B05);四川省杰出青年学术与技术带头人培育项目(2012JQ0008;2012JQ0059);中国博士后科学基金项目(2012T50782)
中图分类号:Q148;S718.5
文献标志码:A
文章编号:1674-5906(2015)06-1143-07
DOI:10.16258/j.cnki.1674-5906.2015.07.011
