长白山森林不同演替阶段比叶面积及其影响因子
2015-03-11胡耀升么旭阳刘艳红
胡耀升, 么旭阳, 刘艳红
北京林业大学森林培育与保护省部共建教育部重点实验室, 北京 100083
长白山森林不同演替阶段比叶面积及其影响因子
胡耀升, 么旭阳, 刘艳红*
北京林业大学森林培育与保护省部共建教育部重点实验室, 北京 100083
比叶面积(SLA)是植物功能性状的重要指标,反映了植物生长过程中资源收获策略。以长白山森林演替过程中4个阶段的典型群落为研究对象,分析了不同演替阶段比叶面积的差异及影响因素。结果表明:比叶面积随演替的发生进行呈现明显增加趋势,且差异显著(P<0.05)。相关分析结果表明,比叶面积与海拔、坡位和土壤氮含量均呈显著正相关关系,与其他影响因子无显著相关关系。进一步的通径分析结果表明,演替前期坡位是主要的影响因素,演替中期模型解释度不高,各通径系数较小,演替后期土壤氮含量对比叶面积的作用凸显。
长白山森林类型; 演替阶段; 比叶面积; 通径分析
近年来,植物功能性状及其与环境因子间的关系受到越来越多的关注[1- 3]。在各项植物功能性状中,叶作为植物与环境接触面积最大的器官,其功能性状的变化被认为是对特定环境的适应性表现,并且这些性状在植物对资源的获得、水分关系和能量平衡方面有重要作用[4- 5]。比叶面积定义为单位重量叶片面积 (SLA),是表征植物生长过程中资源收获策略的关键叶性状指标,能够反映植物在不同生境下资源获取能力,所以被认为是研究特定环境条件下植物生理生态学策略的首选指标[6- 7]。目前,国内相关研究多集中在物种水平[8- 10],或者是某一特定时间的研究[11- 12],而鲜见从群落演替角度对叶功能性状的研究。群落演替的动态变化格局反映了群落环境的变化和不同生态适应型植物构成对这种变化的响应过程[13],同时,某一(些)功能性状很有可能决定着植物种群在竞争中的结果,最后导致群落结构和性质产生改变,进而推动演替[14]。McGill[15]曾指出,群落生态学应该以植物功能性状为基础,通过研究性状、环境梯度、相互作用、行为趋势这4个方面得到群落演替的普遍规律。以往的相关研究多利用多元回归进行统计分析[16- 18],在此过程中由于回归系数带有单位,使环境因子对功能性状的影响效应不能直接进行比较,从而不能比较各自环境因子的相对重要性,而运用通径分析则可以很好得解决这一问题,可对环境因子的相对重要性进行定量评价[19]。
阔叶红松林作为长白山地区的地带性顶极群落,其演替规律的研究一直是生态学关注的重点,同时也是红松阔叶林合理经营的基础[20]。本文以阔叶红松林不同演替阶段的植物群落为对象,将乔木层优势树种的比叶面积作为指标,旨在分析:(1)红松林演替过程中植物群落比叶面积的差异;(2)应用通径分析探讨各项环境因子对比叶面积的直接和间接影响,进一步认识长白山森林在不同演替过程中比叶面积的适应特征。本研究可为长白山森林群落动态演替过程提供理论基础和数据支持。
1 研究地区与研究方法
1.1 研究地区概况
研究区域设置在长白山自然保护区(N42°24′,E128°06′)的红松阔叶林带,该林带分布在海拔720—1100 m范围内,属于受季风影响的温带大陆性山地气候,年平均温度3.3 ℃,最高温出现在8月,平均20.5 ℃,最低温出现在1月,平均16. 5 ℃。年降雨量600—900 mm,土壤为山地暗棕色红壤,土层厚度20—100 cm。植被丰富,乔木主要有:红松(Pniuskoraiensis)、色木槭(Acermono)、蒙古栎(Quercusmongolica)、紫椴(Tiliaamurensis)、水曲柳(Fraxinumandshurica)等;灌木主要有:假色槭(A.pseudo-sieboldiarum)、东北溲疏(Deutziaamurensis)、青楷槭(A.tegmentosum)和毛榛(Corylusmandshurica)等;草本包括:山茄子(Brachybotrysparidiformis)、薹草(Carexspp.)及一些蕨类[21]。
1.2 野外调查和样品采集
本研究于2012年7月—8月进行。采用样方调查法,对30年生次生杨桦林、70年生次生阔叶林、130年生针叶林以及200年生阔叶红松林等4种处于不同演替阶段的主要林型进行样方调查。每种林型随机选择4个20 m×20 m样地,在样地中进行群落学调查。对乔木进行每木检尺,记录树高、胸径、冠幅,再将样地分成16个5 m×5 m的单元格在坐标纸上标记每棵树的位置。记录每个样地的海拔高度、坡度、坡位和坡向。
计算每个样地乔木层各物种的重要值,选取重要值占优势(大于0.1)的物种进行叶片采集。确定优势种后,每个样地中,每一优势种选择5株生长成熟、长势良好的个体,采集完全展开、没有病虫害且未被遮光的叶片。尽量选取在树冠外层枝条上的叶片,将叶片置于两片湿润的滤纸之间,放入自封袋内,带回实验室,放入冰箱保鲜层中储存。
在每个样地随机挖4个土壤剖面,采集0—20、20—40 cm土层的土壤样品,装入土袋,带回实验室,将土样放至通风干燥处,剔除石粒和草根等杂物,自然风干备用。
1.3 指标测定方法
用Epson PerfectionV700 Photo扫描仪进行叶面积扫描,用Winfolia 2007进行叶面积的计算,得到叶面积即叶大小(cm2);SLA=叶片面积(m2) /叶片干重(kg)[22];土壤样品分别过2目和100目筛。全氮的测定采用凯氏定氮法;全磷的测定采用钼锑抗比色法[22]。
1.4 数据处理与分析
1.4.1 重要值的计算
采集样品之前进行乔木层各物种重要值的计算,选取重要值占优势(大于0.1)的物种进行叶片采集和测定。重要值的计算公式如下:乔木重要值=(相对密度+相对优势度+相对频度)/300;各演替阶段测定的优势种如表1所示。
1.4.2 环境因子的测定
数据输入过程中,海拔、坡度数据均以实际观测记录值表示(表1)。坡位数据1表示上坡位,2表示中坡位,3表示下坡位。坡向数据是以朝东为起点(即为0°)顺时针旋转的角度表示,采取每45°为一个区间的划分等级制的方法,以数字表示各等级:1表示北坡(247.5—207.5°),2表示东北坡(292.5—337.5°),3表示西北坡(202.5—247.5°),4表示东坡(337.5—22.5°),5表示西坡(167.5—202.5°),6表示东南坡(22.5—67.5°),7表示西南坡(112.5—167.5°),8表示南坡(67.5—112.5°),即数字越大,表示越向阳,越干热[23]。土壤养分数据见表2。

表1 不同演替阶段群落基本特征与优势种组成Table 1 Characteristics and Dominant species of forests at different succession stages
Ⅰ:30a次生杨桦林30a Poplar-Birch Forest;Ⅱ:70a次生阔叶林70a broad-leaved forest;Ⅲ:130a针叶林130a aciculiailvae;Ⅳ:200a阔叶红松林200a broad- leaved Korean pine forest;坡位数据1表示上坡位,2表示中坡位,3表示下坡位; 坡向数据是以朝东为起点(即为0°)顺时针旋转的角度表示,采取每45°为一个区间的划分等级制的方法,以数字表示各等级:1表示北坡(247.5—207.5°),2表示东北坡(292.5—337.5°),3表示西北坡(202.5—247.5°),4表示东坡(337.5—22.5°),5表示西坡(167.5—202.5°),6表示东南坡(22.5—67.5°),7表示西南坡(112.5—167.5°),8表示南坡(67.5—112.5°),即数字越大,表示越向阳,越干热[23]
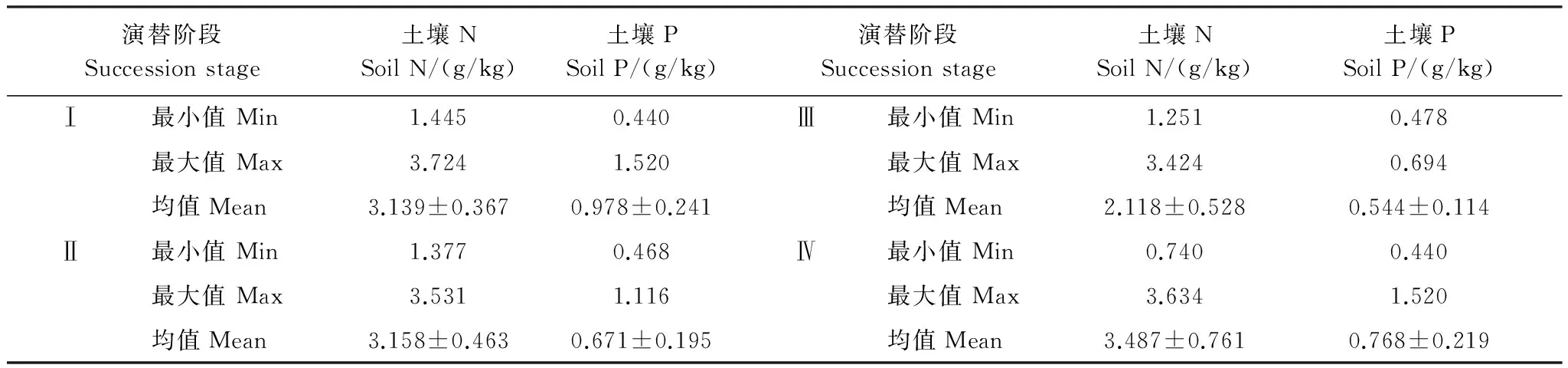
表2 不同演替阶段群落土壤养分特性Table 2 Soil characteristics of forests at different succession stages
1.4.3 数据处理
对各优势种的重要值及其比叶面积值进行加权,获得各样地优势树种的加权值,同一群落间进行平均,从而对不同演替阶段进行比较。不同演替间比叶面积的差异评价采用单因素方差分析(One-way ANOVA),各指标间的相关性分析由 SPSS18. 0 完成。文中图表用 Origin 7.5 和 Excel 2007 完成。
通径分析是多元统计分析方法的一种,根据用户的研究需要构建模型,定义外生变量和内生变量以及它们之间的因果关系(以单向箭头表示的通径系数和双向箭头表示的相关系数来描述),通过联立方程组求解通径系数,从而检验各变量之间的作用方向、作用强度和解释能力。通径系数实际上是标准化的偏回归系数,变量之间的作用强度可通过线性可加的通径系数的代数和来描述[19], 通径分析采用美国北卡罗来纳SAS institute 公司研发的SAS 9.2完成。
2 研究结果
2.1 不同演替阶段森林群落间比叶面积的比较

图1 不同演替阶段比叶面积变化趋势 Fig.1 The variation of SLA between different succession stages
演替群落的各个成员由于其优势度不同,在决定群落的结构和生态功能上作用不同,演替群落与其所在环境也有着不可分割的联系,生境特征决定着森林群落的种类;同时森林群落也不断影响着生境的许多特征[24]。因此,本文试图依据重要值的概念,对不同演替阶段中优势物种的比叶面积进行加权,从而得到整个演替群落的功能性状总体特征,进而比较不同群落的演替特征。
由图1可看出比叶面积在不同演替阶段表现为:演替前期<演替中期Ⅰ阶段<演替后期<演替中期Ⅱ阶段。其中演替中期Ⅱ阶段的SLA与演替前期的SLA有显著差异(P<0.05);其他各演替阶段的SLA均无显著差异(P>0.05)
2.2 比叶面积与各环境因子间的相关分析
相关分析结果如表3所示,结果表明:比叶面积与海拔、坡位和土壤N均呈显著正相关关系,其他指标间的相关性不显著。

表3 比叶面积与环境因子相关系数Table 3 Pearson correlation coefficients among SLA and environmental factors
*P<0.05;SLA:specific leaf area

图2 演替前期通径路径图Fig.2 Path analysis model of early succession
2.3 通径分析模型建立及各演替阶段比较分析
在相关性分析的基础上,通过通径分析建立合理的路径图可以更加明确比叶面积与各环境因子的关系。相关分析中,土壤P、坡度、坡向与比叶面积之间无显著的相关关系,因此在建立通径分析路径之前,首先将这些指标剔除,不纳入考虑[25]。将各个演替阶段的数据进行计算,得出各影响因子对比叶面积的通径系数。各演替阶段的通径分析路径图均通过了模型检验。
2.3.1 演替前期通径分析路径图及通径系数表
如图2所示,显著的路径(P<0.05)共有3条:海拔对比叶面积、坡位对比叶面积、海拔对土壤N,其他路径均不显著(P>0.05)。
如表4所示,各影响因子对于比叶面积的解释度为0.913。海拔、坡位和土壤N对于比叶面积的总影响的通径系数分别为-0.212、-0.906、-0.336。

表4 演替前期通径系数表Table 4 Path coefficient of early succession
2.3.2 演替中期Ⅰ阶段通径分析路径图及通径系数表
如图3所示,显著的路径(P<0.05)共有2条:土壤N对比叶面积、坡位对比叶面积,其他路径均不显著(P>0.05)。
如表5所示,各影响因子对比叶面积的解释度为0.686。各因子中,坡位对比叶面积的总影响通径系数最大,为0.759;其次为海拔,通径系数为0.548;土壤N最小,通径系数为0.400。

表5 演替中期Ⅰ阶段通径系数表Table 5 Path coefficient of middle successionⅠ

图3 演替中期Ⅰ阶段通径路径图Fig.3 Path analysis model of middle successionⅠ
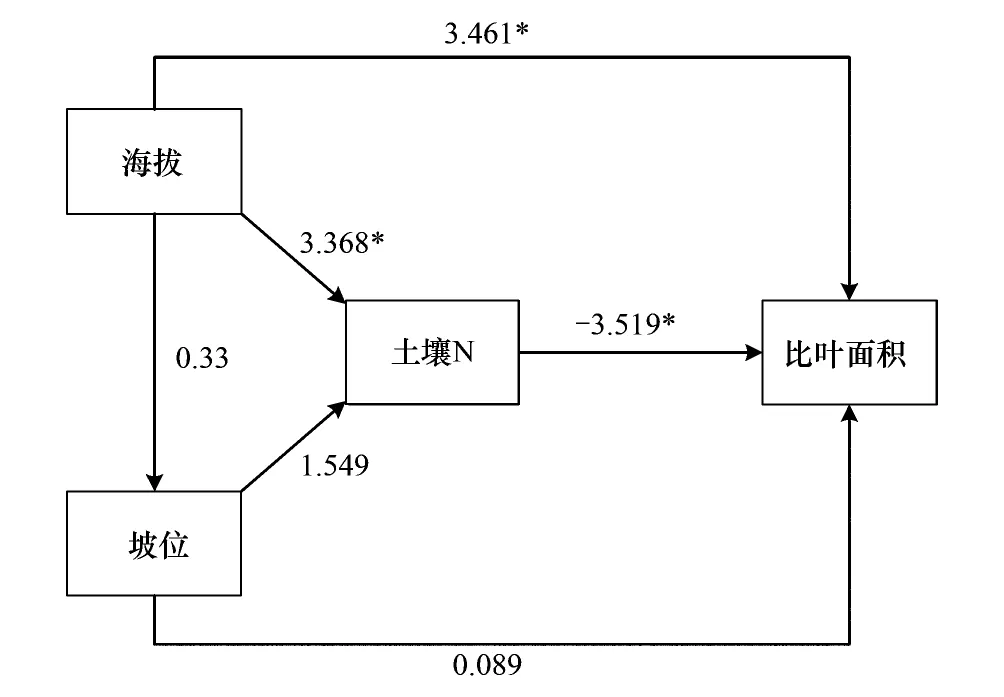
图4 演替中期Ⅱ阶段通径路径图Fig.4 Path analysis model of middle successionⅡ
2.3.3 演替中期Ⅱ阶段通径分析路径图及通径系数表
如图4所示,显著的路径(P<0.05)共有3条:海拔对比叶面积、土壤N对比叶面积、海拔对土壤N,其他路径均不显著(P>0.05)。
如表6所示,各影响因子对于比叶面积的解释度为0.719。坡位对比叶面积的总影响通径系数达到2.734;海拔的通径系数为0.125;土壤N对于比叶面积产生负作用,为-0.168。

表6 演替中期Ⅱ阶段通径系数表Table 6 Path coefficient of middle successionⅡ
2.3.4 演替后期通径分析路径图及通径系数表
如图5所示,显著的路径(P<0.05)共有2条:坡位对比叶面积、土壤N对比叶面积,其他路径均不显著(P>0.05)
如表7所示,各影响因子对于比叶面积的解释度为0.853。坡位对比叶面积的总影响通径系数达到0.88;土壤N的通径系数为0.125;海拔对于比叶面积产生负作用,为-0.385。

表7 演替后期通径系数表Table 7 Path coefficient of late succession
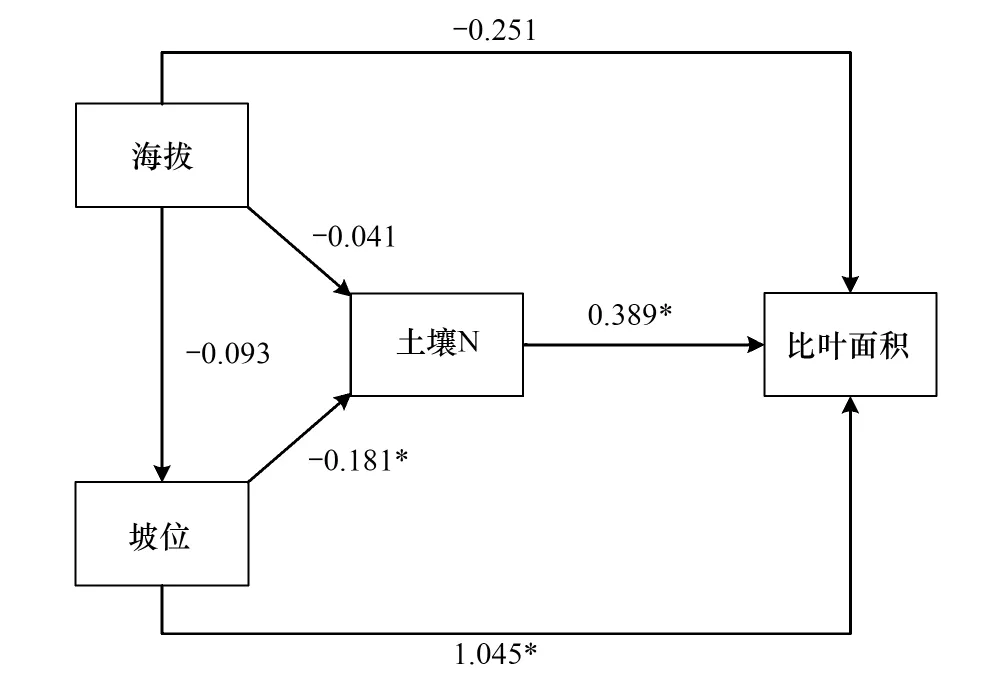
图5 演替后期通径路径图Fig.5 Path analysis model of late succession
3 结论与讨论
3.1 不同演替阶段群落比叶面积的差异比较
本研究表明,随着演替的进行,比叶面积呈现明显增加趋势。比叶面积能够反映植物对营养的获取与利用的平衡关系,与植物的相对生长速率和资源利用有紧密的关系[26]。Wilson等[7]发现在英国植物区系里,比叶面积可以很好地预测植物在资源利用轴上的位置。演替前期植物处于开放的环境,有着较小的比叶面积,叶片可能将大部分干物质用于构建保卫细胞,增加叶厚或叶肉细胞密度,同时使叶片内部水分向叶片表面扩散的距离或阻力增大,降低植物内部水分散失[27]。有相关研究表明,在郁闭林内进行的更新,常绿植物与落叶植物相比有着较低的比叶面积,这降低了单位叶面积的呼吸碳损失,而且通过延长叶寿命增加了碳收获,同时较低的生长速率得以维持正碳平衡[28]。演替中后期,植物所处的环境较为复杂,比叶面积大的植物能很好地保持体内营养,从而适应资源丰富的环境。
3.2 不同演替阶段群落比叶面积差异影响原因分析
植物功能性状受自身生理特性和环境条件影响。本研究通过相关性分析表明比叶面积与海拔、坡位、土壤N呈正相关关系。地形是各环境因子的综合,温度、水分、土壤养分的变化会随地形梯度的变化而变化,进而影响植物功能性状。尤其在小尺度研究领域,这些非生物因子的影响尤其明显[29]。海拔对植物功能性状的影响机理主要是随着海拔的升高,植物生长发育逐渐受到温度和水分的限制,因而适应性会发生显著变化,尤其叶片对环境变化反应更为敏感,通常来讲,生活在相对贫瘠的环境中的植物具有较小的SLA,而将更多的干物质投入用于抵御不良环境[30]。在山地环境中,不同坡位形成的小生境差异较大,上坡位太阳辐射和紫外线照射强、土层薄、土壤贫瘠、土壤含水量低、温度较高等特点[27],与中下坡位形成差异,因而表现出坡位对比叶面积的影响较大。坡向的变化亦会影响到温度、湿度等条件,各种不同坡向间所受得光照时间和强度不同,其所在区域的环境条件必然也会有一定的差异。这种环境条件的差异必然会影响植物的叶片形态[26]。植物的资源利用策略受土壤资源影响,主要体现在两方面:一方面,不同群落功能群的结构来决定不同优势植物的资源策略;另一方面,通过影响物种内的性状变异以影响植物策略的改变[31]。有研究表明,土壤全氮、土壤全磷等对植物地上部分的生长产生重要影响[32]。演替中后期随着土壤全氮的增加而比叶面积增加,这很有可能是因为此阶段针叶树为优势树种,一定程度比叶面积增加可降低植物内部水分散失,而氮素的增加可提高气孔导度,加强光合,在二者共同作用下,一定范围内仍就能够提高植物获取资源能力[33]。
在相关性分析的基础上,本研究运用了通径分析的方法,试图研究各个演替阶段,土壤和地形因子是如何影响比叶面积最终导致其在演替阶段的差异。演替前期,由于环境较为开放,使得各因子间的相互制约变强,均成为比叶面积的显著影响因子。通径分析的结果证实,比叶面积主要受坡位和海拔的影响,总通径系数很大。演替中期,模型的解释度较小,各影响因子的总通径系数也不如其他阶段大,这可能是与演替进入该阶段后,植物群落结构趋于复杂,种间竞争剧烈有关。演替后期,通径分析表明土壤N成为比叶面积的显著影响因子,这可能是因为当演替进入后期阔叶红松林,红松成为优势种类,此类植物具有较高的凋落物氮含量,返还土壤的养分数量有所增加[34]。
综合以上研究结果表明,不同演替阶段上影响比叶面积的因素和作用程度均有不同,且作用的关系有正有负。此外,也不能忽视物种因素的作用,不同演替阶段物种组成不一样,由于其重要值不同,所赋予比叶面积值也有所差异。所以,比叶面积在各演替阶段的差异原因是不同因素综合作用的结果。
[1] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001- 11006.
[2] 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150- 165.
[3] 杨冬梅, 章佳佳, 周丹, 钱敏杰, 郑瑶, 金灵妙. 木本植物茎叶功能性状及其关系随环境变化的研究进展. 生态学杂志, 2012, 31(3): 702- 713.
[4] Vendramini F, Díaz S, Gurvich D E, Wilson P J, Thompson K, Hodgson J G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytologist, 2002, 154(1): 147- 157.
[5] Roche P, Daz-Burlinson N, Gachet S. Congruency analysis of species ranking based on leaf traits: which traits are the more reliable. Plant Ecology, 2004, 174(1): 37- 48.
[6] 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844-852.
[7] Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155- 162.
[8] 刘贤娴, 李俊清. 北京野鸭湖湿地植物叶功能性状研究. 安徽农业科学, 2008, 36(20): 8406-8409.
[9] 闫道良, 任燕燕, 连俊方, 盛琳杰. 3个优势科树种叶功能性状及其关联特性. 林业技术开发, 2012, 26(3): 34- 37.
[10] 黎莉, 冯树丹, 王建波, 倪红伟, 付晓玲, 杨丽影, 徐明华. 三江平原12种湿地植物光合特性与叶功能性状的关系研究. 湿地科学, 2010, 9(3): 225- 235
[11] 施宇, 温仲明, 龚时慧. 黄土丘陵区植物叶片与细根功能性状关系及其变化. 生态学报, 2011, 31(22): 6805- 6814.
[12] 徐庆华, 臧润国, 谢怀慈, 白志强, 郭仲军, 赵玲. 天山北坡4种栽培杨树的功能性状分析. 干旱区研究, 2012, 29(3): 425- 431.
[13] 张继义, 赵哈林, 张铜会, 赵学勇. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态. 植物生态学报, 2004, 28(1): 86- 92.
[14] 付登高, 段昌群. 植物功能性状在植物生态学中的研究进展 // 生态科学进展. 北京: 高等教育出版社, 2007: 97- 121
[15] Mcgill B J, EnquistB J, Weiher E, Westoby M. Rebuilding community ecology from functional traits. Trends in Ecology and Evolution, 2006, 21(4): 178- 185.
[16] 吕佳, 李俊清. 北京百花山植物叶片性状相互关系研究. 科学技术与工程, 2008, 8(12): 3287- 3289.
[17] 龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011, 31(20): 6088- 6097.
[18] 胡梦瑶, 张林, 罗天祥, 沈维. 西藏紫花针茅叶功能性状沿降水梯度的变化. 植物生态学报, 2012, 36(2): 136- 143.
[19] 任红松, 吕新, 曹连莆, 袁继勇. 通径分析的SAS 实现方法. 计算机与农业, 2003, (4): 17- 19.
[20] 于振良 赵士洞 董立荣, 张元林. 阔叶红松林的研究综述. 吉林林学院学报. 1996, 12(4): 235- 238.
[21] 姬兰柱, 董百丽, 魏春艳, 王淼. 长白山阔叶红松林昆虫多样性研究. 应用生态学报, 2004, 15(9): 1527- 1530.
[22] 鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 2000: 135- 155.
[23] 张峰, 张金屯, 张峰. 历山自然保护区猪尾沟森林群落植被格局及环境解释. 生态学报, 2003, 23(3): 421- 427.
[24] 戈峰. 现代生态学. 北京: 科学出版社, 2002: 210- 211.
[25] Grace J B, Kelley J E. A structural equation model analysis of post fire plant diversity in California shrub lands. Ecological Applications, 2006, 16(2): 503- 514.
[26] Ackerly D D, Knight C A, Weiss S B, Barton K, Starmer K. Leaf size, specific leaf area and microhabitat distribution of chaparral plants: contrasting patterns in species level and community level analyses. Oecologla, 2002, 130(3): 449- 457.
[27] 祁建, 马克明, 张育新. 北京东灵山不同坡位辽东栎(Quercusliaotungensis)叶属性的比较. 生态学报, 2008, 28(1): 122- 128.
[28] Sterck F J, Poorter L, Schieving F. Leaf traits determine the growth-survival trade-off across rain forest tree species. The American Naturalist, 2006, 167(5): 758- 765.
[29] Loreau M, Naeem P, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804-808.
[30] Westoby M, Falster D S, Moles A T, Vesk P A, Wright I J. Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 2002, 33: 125- 159.
[31] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas SC, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827.
[32] 刘旻霞, 马建组. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295- 3300.
[33] 李永华, 罗天祥, 卢琦, 田晓娅, 吴波, 杨恒华. 青海省沙珠玉治沙站17种主要植物叶性因子的比较. 生态学报, 2005, 25(5): 994- 999.
[34] 刘颖, 武耀祥, 韩士杰, 林鹿. 长白山四种森林类型凋落物分解动态. 生态学杂志, 2009, 28(3): 400- 404.
Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains
HU Yaosheng, YAO Xuyang, LIU Yanhong*
TheKeyLaboratoryforSilvicultureandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China
Specific leaf area(SLA) is an important indicator of plant function trait, reflecting resources utilization strategy during the plant growth process. The typical forest communities under four different succession stages in Changbai Mountains were selected,the SLA and its influencing factors in different stages of successions were compared. The results showed that SLA was obviously different across forest successions,and significantly increased with forest succession stages. Correlation analysis showed that SLA had significant positive correlations with elevation、slope position、soil nitrogen (withP<0.05). Further path analysis indicated that on early stage of succession, slope position was an important factor, while Path analysis model was with low path coefficient on middle stage of succession, and the negative effect of soil N was significant on later stage of succession.
Changbai Mountains; community succession; specific leaf area;path analysis
国家“十二五”林业科技支撑项目(2012BAC01B03- 3)
2013- 10- 13;
日期:2014- 07- 14
10.5846/stxb201310132459
*通讯作者Corresponding author.E-mail: liuyh@bjfu.edu.cn
胡耀升, 么旭阳, 刘艳红.长白山森林不同演替阶段比叶面积及其影响因子.生态学报,2015,35(5):1480- 1487.
Hu Y S, Yao X Y, Liu Y H.Specific leaf area and its influencing factors of forests at different succession stages in Changbai Mountains.Acta Ecologica Sinica,2015,35(5):1480- 1487.
