海南海岸青皮林繁殖物候特征
2015-03-10刘淑菊郝清玉
吕 冰,王 娜,刘淑菊,郝清玉
热带动植物生态学省部共建教育部重点实验室, 海南师范大学生命科学学院, 海口 571158
海南海岸青皮林繁殖物候特征
吕 冰,王 娜,刘淑菊,郝清玉*
热带动植物生态学省部共建教育部重点实验室, 海南师范大学生命科学学院, 海口 571158
石梅湾海岸青皮林是海南岛重要的天然海防林及旅游区风景林,对其繁殖物候的研究有助于揭示青皮在贫瘠海岸沙滩成功定居及繁殖的生物学特性。采用种群、个体定位和小枝标记相结合的方法,连续3a对其开花物候进行观测,结果表明:青皮个体繁殖物候期约为101d,小枝平均花蕾数为133.3个,幼果坐果率为42.4%,结实率为4.4%;青皮种群为1a连续开花型,但集中开花模式明显,有2个高峰期且花量多,非集中开花期花量很少;个体间及个体中小枝间开花存在一定的不同步性;开花及种子丰年现象明显,歉年花量及种子雨量很少;开花物候期前1个月的降雨量对开花物候期无显著影响,但3、4月份平均气温分别高于24℃和26.4℃,开花期提前明显,低于此温度开花期推迟相对缓慢,即物候期的提前与推迟对温度的上升与下降的响应是非线性的。
海岸青皮林; 繁殖物候; 海南岛
植物物候是指植物受生物因子和非生物因子影响而出现的以年为周期的自然现象,包括各种植物的发芽、展叶、开花、叶变色、落叶等[1]。物候对环境变化起到越来越重要的指示作用,它是监测生态系统响应气候变化的一个直观的指示器[2],对植物多样性保护、林业管理、病虫害控制、以及人类健康等具有重要的作用[3]。此外,植物物候对全球气候变化响应的研究可以揭示全球变化的生物学响应机制,是植被-气候关系研究中的重要环节[4]。根据开花时间和频度,可将植物分成4种类型:统一开花型,以3—10a的不规则周期大量开花,占35%;多年开花型,占19%;1a开花型,占13%;1a多次开花型,占5%[5]。植物物候及其变化受气温、光照、水分等气候因子的综合影响,其中温度是影响物候变化最重要的因子,水分成为胁迫因子时对物候的影响也十分重要[4]。植物种类不同,其开花物候的影响因素也会不同,如:冬季和早春植物开花的主要影响因素是温度,晚春和早夏植物开花受温度和日照同时影响,而晚夏植物花粉的开始传播主要受日照的影响[6]。研究结果表明,全球气温升高对植物物候期有明显的影响,年平均温度上升1 ℃,各种木本植物物候期春季一般提前3—4 d,秋季一般推迟6—8 d,大部分植物始花期提前3—6 d[7];德国对20种植物的研究发现春季温度每上升1 ℃,物候提前2.5—6.7 d,生长季延长2.4—3.5 d[8]。降雨模式也对植物开花物候也有显著影响,如:地中海地区雨季延迟导致树木和灌木开花日期延期且开花数量减少[9]。
青皮(Vaticamangachapoi)为龙脑香科(Dipterocarpaceae)青梅属(VaticaLinn.), 濒危物种,是国家Ⅱ级重点保护植物。青皮是青梅属在海南岛的仅存的一个种[10- 11],且仅石梅湾海滨才有大片的单优青皮林。相对人工海防林,青皮林物种丰富,群落层次结构复杂,天然更新良好,植株高大,树冠茂密,具有较强的群体抗风能力和耐瘠薄特点,是海南岛最为理想的海岸沙滩海防林,也是人工海防林向天然林恢复的示范林[12]。此外,青皮犹如珍珠玉滴般的花蕾、淡香圣洁的白花及钟状鲜红的种子萼片,与树木一起构成了白、绿、红三色美景,为石梅湾旅游区增色不少。目前对青皮林的研究主要集中在群落结构、群落多样性、青皮遗传多样性、天然更新等方面的研究,有关青皮林繁殖物候方面的研究还未见报道。本文主要研究(1)青皮繁殖物候的生物学特性;(2)青皮繁殖物候对气候变化的响应,以期为青皮植物的保护与管理,青皮花期、种子萼片变红期及种子雨期的预测等提供理论依据。
1 研究地概况
海岸青皮林位于海南岛东部万宁市沿海,18°39′ N, 110°14′ E。林带长约22 km,宽约400 m,离潮汐线约50—100 m,沿海岸呈带状分布,是世界上仅有的两片海滩青皮林之一,是国家重点珍稀植物保护区[13]。保护区属热带海洋性季风气候,终年高温,干湿季明显。年均气温24.5 ℃,光照率50%,年均降水2032 mm,雨量集中在5—10月,约占全年总降水量的80.2%。沿海滩分布的是晚期海相新砂堤沉积及近代海滩沉积,岩性为浅黄色或灰白色中粗砂[14]。该区植物群落属于热带雨林植被类型中的海岸单优青皮群落,植物种类繁多,野生植物资源有69科143属 172种之多[13]。群落乔木层以青皮为优势种,灌木层主要由乔木层树种的幼树组成,其中以青皮的幼树占绝对优势;草本层植物种类与数量均较少,层间植物主要由木质藤本植物组成。
2 研究方法
2.1 繁殖物候观测方法
以石梅湾海岸青皮林核心区作为研究对象,采用种群整体观测与单株树木定位观测相结合的方法。种群整体观测主要记录青皮种群繁殖物候的变化期,单株树木定位观测采用系统抽样与典型抽样相结合方法。2010年选取16株生长良好的青皮作定位观测个体,2011年定位观测个体增加到30株。观测个体平均间距约为10 m,记录其胸径,并在每株树上选择2个基本一致的小枝编号挂牌进行定期观测,观测期为3a,从2010—2012年。为了观测、记录方便,仅选择生长在路旁林缘的青皮树作为定位观测木,挂牌小枝位于树的中下部,标记牌号统一挂在距小枝顶端约15 cm处(图1-i)。根据青皮繁殖物候期各阶段生长状况的不同,观测间隔期为1—2周。每次观测记录青皮种群整体的繁殖物候情况,定位观测木的花期生长及结实情况,并对标记小枝的花蕾、开花及结实等情况进行详细计数,直至繁殖期结束种子全部落地。
2.2 繁殖物候期的划分标准
根据热带森林生态系统定位观测指标体系标准(LY/T 1687—2007)[15],将青皮繁殖物候期划分为6个阶段,即花蕾期、始花期、盛花期、凋谢期、幼果期和成熟期。(1)花蕾期:观测树的花序露出花苞长约2—10 mm 时即属于花蕾期;(2)始花期:观测树有一朵或同时几朵的花瓣开始完全开放,即为开花始期;(3)盛花期:观测树有50%的花蕾都展开花瓣为开花盛期。统计标记小枝上开花数,当开花数占花蕾总数一半以上的时候即为盛花期;(4)凋谢期:观测树上的花瓣凋谢脱落,只剩下5%开花数为凋谢期;(5)幼果期:青皮花瓣凋落后,花托开始膨大发育,种子雏形显现,后期继续生长,种子形状明显;(6)成熟期:种子形状饱满,颜色由绿色转为褐色,即进入种子成熟期。
2.3 数据处理
小枝平均花蕾数 = 标记小枝的花蕾总数/有花蕾的小枝样本数
小枝平均幼果数 = 标记小枝的幼果总数/有幼果的小枝样本数
小枝平均结实数 = 标记小枝的成熟种子总数/有花蕾的小枝样本数
幼果坐果率 = 幼果数/总花蕾数
结实率 = 成熟种子数/总花蕾数
使用Excel2003和SPSS16.0软件分别对标记的青皮小枝、个体以及种群的开花物候期及座果率等指标作相关的统计分析。
3 结果和分析
3.1 单花开花进程
青皮为两性花,单花开放过程可分为始蕾、初开、盛开、落花、幼果、成熟6个阶段,每个阶段的持续时间变化大。圆锥花序生于上部叶腋内,花序密被星状短绒毛,花蕾从叶腋处或花芽中冒出,鳞片紧密包裹着花瓣,花瓣露白部分长至8—10 mm,原来紧紧包裹的花瓣开始逐渐松开,但尚未裂开,历经约10—15 d,此为花蕾期;花瓣继续松动瓣状裂开,花蕾顶端出现小口,但未能看到雌雄蕊,花型呈线状匙形,持续1—2 d此为初开阶段;花朵继续开放,花瓣完全裂开向外伸展,花瓣5片呈辐射状,可见伸长的乳白色柱头,持续3—5 d此为盛开阶段;花瓣变黄萎蔫,花朵开始凋落,花瓣脱落,花瓣基部可见淡黄色雄蕊,连同宿萼包裹子房,子房球形,密被星状微柔毛,柱头外露,呈头状,历经3—7 d,雌雄蕊干枯掉落,此为落花阶段。花朵开放时,一般开口朝下,有淡淡清香味。花瓣掉落初期,形成球形幼果,萼片大小一致,幼果在成熟之前发育很慢,期间要经过2个月左右的成长期,直到7月下旬种子成熟,幼果发育过程中,宿萼扩大成果翅时,通常是5片果翅,2长3短,从原来的3—4 mm长至3—5 cm不等,并且逐渐由绿色变深红色,最后变为黄褐色或棕褐色即标志着种子成熟。
3.2 小枝开花数量、物候及特征
3a标记的小枝平均数为48枝,标记小枝的平均开花率为61.9%,其中,2011年小枝的开花率最高,为96.4%,2012年最低,仅为25.0%;3a标记的小枝平均花蕾数、平均幼果数和平均成熟种子数分别为133.3个、50.3个和4个,其中,2010年小枝平均花蕾上最多,为203个,2011年小枝平均幼果数和成熟种子数最多,分别为82个和8个;3a小枝年均幼果座果率和结实率分别为42.4%和4.4%,其中,2011年幼果坐果率和结实率最高,分别为71.9%和9.8%,2010年幼果坐果率和结实率最低,分别为19.2%和1.0%(表1)。3a标记小枝的开花同步性比较明显,两枝同步开花的比例平均为62.9%,两枝不同步开花比例为6.9%,而只开1枝的比例为30.2%,不同步时间约相差2.3周,其中,2011年小枝开花同步性最高,为82.1%,2012年同步性最差,仅为40%,且不同步间隔期长达6周(表2)。
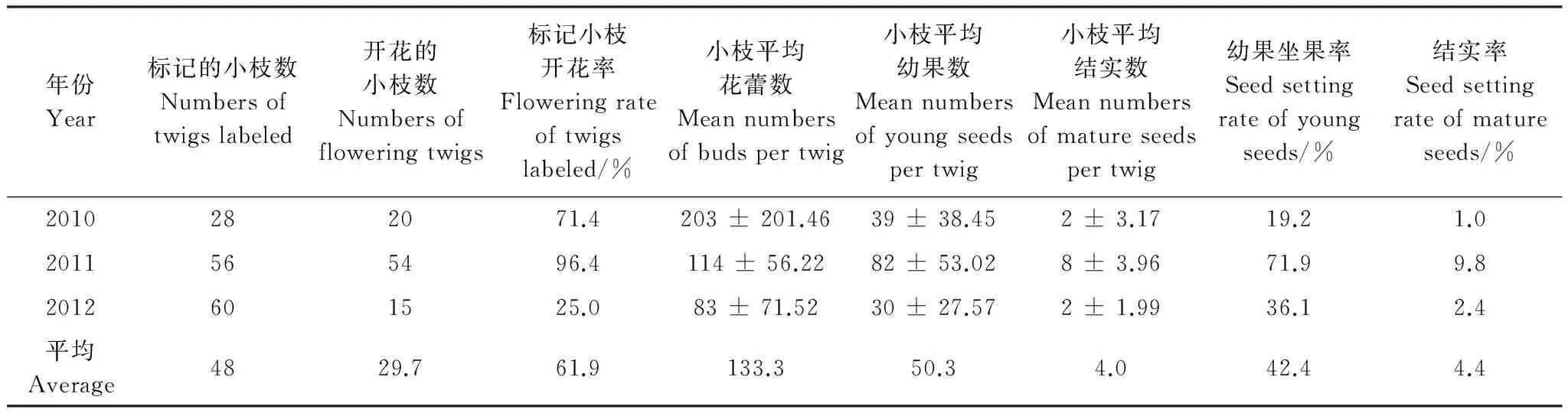
表1 小枝花蕾及结实数量Table 1 The quantity of buds and mature seeds in twigs

表2 小枝3a开花同步性比较Table 2 The comparison of flowering synchronicity in twigs for 3 years
3.3 个体物候及特征
青皮繁殖物候的花蕾期、始花期、盛花期、凋谢期、幼果期、种子成熟期6个时期的形态特征详见图1,其中现花蕾期是花芽鳞片未展、花苞未露阶段,属于花蕾期前期;幼果初期与幼果中期皆是表示一个幼果变大果翅变长并发生颜色由绿变深红的一个动态变化,但均属于幼果期。
青皮个体的开花率较高,3a平均值达90%以上,其中,2010年、2011年和2012年个体开花比例分别为100%、 93.5%和91.2%。因气候条件的不同,年际间青皮各物候期的起始时间及整个物候期的持续时间均会出现小幅的变化。青皮从花蕾期至种子成熟整个繁殖物候期经历的时间较长,平均经历101 d,变化区间为98 —105 d,即年际间繁殖物候期会因气候条件的不同而延长或缩短1—2周;青皮个体的花蕾期在4月中下旬—5月下旬,持续期约为5周,盛花期在5月中下旬或6月中下旬,持续期较短,约为1周,幼果期持续时间较长,约为5—6周,种子成熟期在7月下旬(表3)。此外,不仅个体中的小枝开花存在不同步性,青皮个体之间开花也存在不同步性,且同一个体的树冠顶部与底部也存在不同步性,处于遮荫处部分开花一般要延迟1—3周。

图1 青皮开花物候进程Fig.1 Vatica mangachapoi flowering phenology processa: 现花蕾期early period of buds;b: 花蕾期bud period;c: 始花期initial period of flowering;d: 盛花期full flowering period;e: 凋谢期withering period;f: 幼果初期early period of young seeds;g: 幼果中期(萼片变红)medium period of young seeds (calyx lobes changed red);h: 种子成熟期mature period of seeds; i: 小枝标记部位sites labeled in twigs
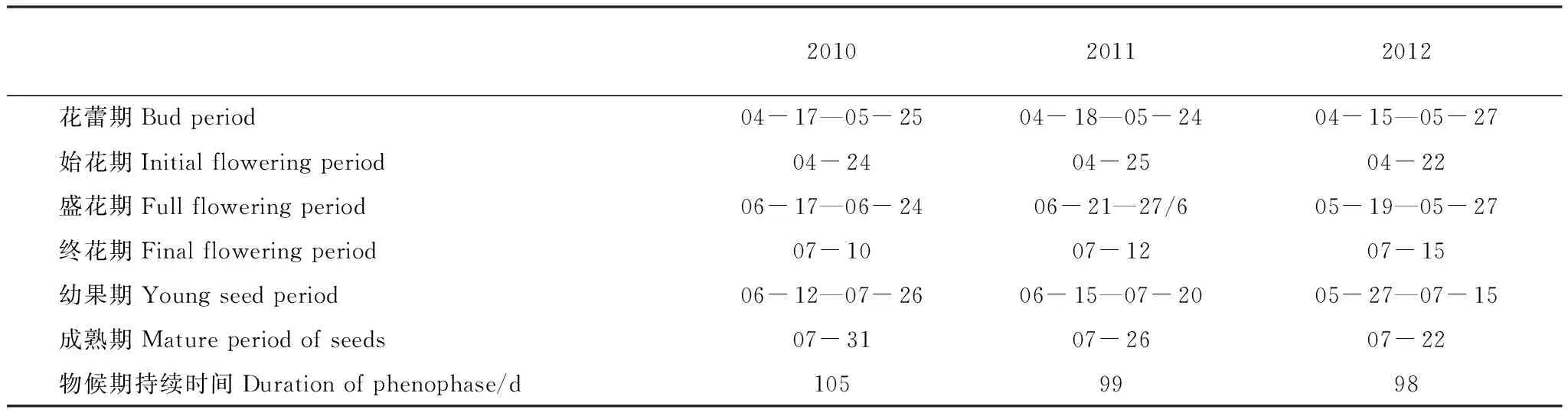
表3 青皮个体繁殖物候期(月-日)Table 3 Individual reproductive phenophases of Vatica mangachapoi
3.4 种群物候及特征
由于青皮个体花期的不同步性,使青皮种群的繁殖物候期要比个体的繁殖物候期长。种群繁殖物候期主要集中在4月中下旬至9月上旬,物候期持续约4个月,期间两次开花数量达到高峰期,即有两批盛花期,与其相对应有2批花蕾期。表4所示,第1批花蕾期出现在4月中下旬,花蕾持续时间2—6周,平均约3周。55.9%的个体间隔1—3周后,出现了第2批花蕾,第2批花蕾期出现在5月下旬或6月中旬,持续时间是1—7周,平均约3周。第1批盛花期发生在5月上旬或中旬,花量大,满树开白花,持续时间为3—5 d,于5月下旬或6月中下旬期间出现第2批盛花期,花量较第1个盛花期少,持续时间为7—10 d。2010年和2012年第1批盛花期相近,均出现在5月上旬,而2011年的第一批盛花期则推迟了约10d;第2批盛花期2010年出现在6月中旬,2011年出现在6月下旬,而2012年较前两年提前了3—4周,出现于5月下旬。花凋谢期出现在7月中旬。花朵凋谢后进入幼果期,幼果期发育缓慢,果翅逐渐曾长并变为深红色,直到7月下旬种子才逐渐成熟。由于开花的不同步性,种子成熟也呈现出不同步性,其中,第1批种子成熟集中在7月下旬,持续时间约为1周,第2批种子成熟期发生在8月中下旬,持续期也约为1周。因此,种子最佳采摘时间是在7月下旬或是8月上旬,这时种子数量较多。此外,种群几乎全年都有植株开花,但花量极少,开花不明显,且仅见于个别小枝。

表4 青皮种群繁殖物候期动态变化Table 4 Dynamic change of Vatica mangachapoi reproductive phenophase
3.5 气候对开花物候的影响
研究结果表明植物开花物候与花期前特定时期的平均气温显著相关,与整个前生长期的总度日数不相关[16]。青皮花蕾盛期通常发生在4—5月份,因此本文选择青皮花蕾盛期前一个月,即3、4月份的气象资料分析气候对开花物候的影响。3、4月份月平均气温数据表明,2010年气温与3a的平均气温接近,2011年气温偏低,3、4月份月平均气温比2010年同期分别低2.7 ℃ 和1.2 ℃,而2012年气温有些偏高,尤其是4月份气温比2010年同期高0.8 ℃(表5)。由于2011年比2010年 3、4月份气温偏低,2011年两批盛花期比2010年分别推迟了9 d 和7 d;2012年与2010年的3月份气温相当,第1批盛花期也基本相同,但4月份气温与2010年同期高0.8 ℃,第2批盛花期提前了23 d。结果表明温度升高远比温度降低对开花物候期的影响敏感。3、4月份降雨量对盛花期的影响不明显,这是因为2011年 2批盛花期均推迟,但3、4月份的降雨量出现完全相反的结果,2011年 3月份降雨量为3个年度同期降雨量的最大值,而4月份的降雨量为3个年度同期的最小值。2012年3、4月份的降雨量比2010年 同期分别高66 mm和45.4 mm,但第1批盛花期相同,第2批盛花期却提前了23 d。
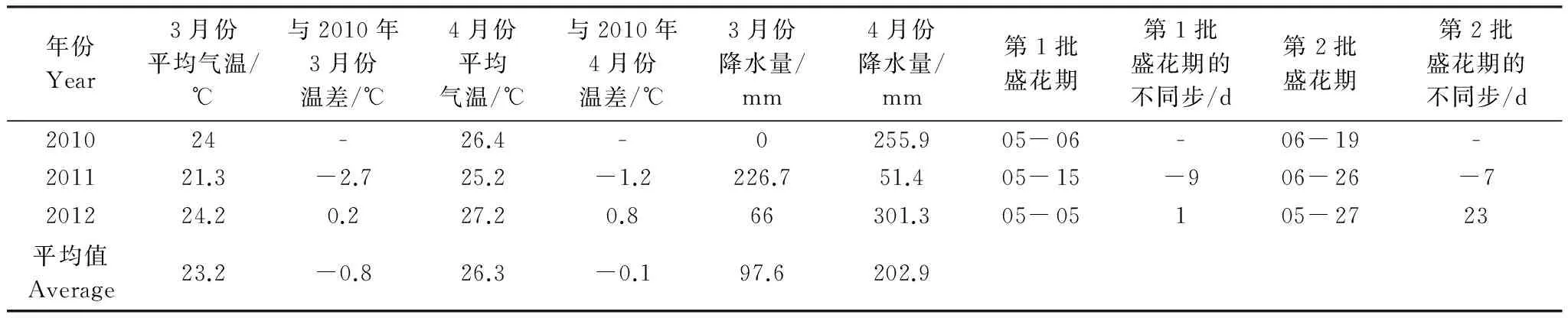
表5 温度与降雨量对盛花期的影响Table 5 The impact of temperature and rainfall on the full flowering period
3月份平均气温Average temperature in March; 与2010年3月份温差Temperature difference with March of 2010; 4月份平均气温Average temperature in April; 与2010年4月份温差Temperature difference with April of 2010; 3月份降水量Rain fall in March; 4月份降水量Rain fall in April; 第1批盛花期Full flowering on first batch; 第1批盛花期的不同步Asynchronous dates of full flowering period on first batch; 第2批盛花期Full flowering on second batch; 第2批盛花期的不同步Asynchronous dates of full flowering period on second batch
4 结果与讨论
(1)青皮种群几乎全年都有开花现象,但其繁殖物候期主要集中在4月中下旬至9月上旬。在此期间,有两次开花高峰期,发生在5—6月份,持续时间近2个月,花量呈现出一强一弱的现象。非集中开花期,青皮只是各别小枝的少量开花,开花现象不明显。青皮这种开花模式与长柄双花木(Disanthuscercidifoliusvar.longipes)相类似,均属于“大量集中开花”模式[17],而非“持续开花”模式,如假叶树(Ruscusaculeatus)具有7个月的开花期[18]。青皮在个体和小枝之间存在开花物候不同步现象,因此在单花、个体和种群的始花期和开花持续期等方面存在差异,其原因是受生物因子的影响,这也与长柄双花木具有同样的特征[17,19]。青皮个体开花物候的不同步性是随机现象,还是遗传特性决定一些个体总是早些开花,而另一些个体总是晚些开花还有待于深入研究。
(2)青皮年际间开花数量及坐果率存在显著差异,表明青皮具有种子丰年和歉年现象,其中2011年开花数量和坐果率均显著高于其他2个年份,为种子丰年,相比之下2010年和2012年为种子歉年。花蕾数量、幼果数量与结实数量的比例为33.2∶12.5∶1,说明青皮的结实量远低于开花数量。其主要原因是由于海南台风、暴雨等恶劣气候使得大量花蕾、花朵以及幼果被刮落,加之虫害严重,从而造成青皮的结实率很低。
(3)3a的观测数据表明青皮开花物候年际间变化对气温变化影响明显,对降雨量影响则不明显。3、4月份平均气温分别高于24 ℃ 和26.4 ℃ 时,开花期提前明显;低于此温度,开花期推迟相对缓慢,这一现象证实了青皮物候期的提前与推迟对温度的上升与下降的响应也是非线性的[20- 21]。温度升高可促进酶的活性,加快植物物候进程,特别在植物各生长发育期的前期,各种物候期的开始日期与其前期气温之间有显著的相关性[22],因此3、4月份平均气温高于基准温度,青皮开花期提前明显。另外,研究结果表明低纬度热带地区四季气温没有明显差异,植物发育进程主要由旱季的干旱程度及其持续时间决定,因而使得许多热带植物物候行为主要取决于降水量[23],但青皮物候期则属于例外,这可能归因于海南旱季时间较短的缘故。
(4)青皮种子属于玩拗型种子,无休眠期,失水约7—14 d便会死亡[24],因此落种期遇上雨季尤为重要。青皮双峰开花模式使种子成熟期分散,增加了落种期恰逢雨季的机会,这是青皮种群能在滨海沙滩成功繁殖的重要策略之一。
(5)青皮幼果果翅变红期因个体差异而不同。通常幼果发育到中期,大部分果翅由原先的鲜绿色转变为深红色,当变为棕褐色时种子达到成熟,也有少量个体的幼果果翅一直保持鲜绿色,直到种子近熟期才变为深红色。游客来石梅湾旅游区观看青皮花蕾和开花的最佳时期为4月中下旬至6月上旬,观看青皮种子红色萼片的最佳时期为5月下旬至7月中下旬。
[1] 陆佩玲, 于强, 贺庆棠. 植物物候对气候变化的响应. 生态学报, 2006, 26(3): 923- 929.
[2] Penuelas J, Filella I. Phenology: responses to a warming world. Science, 2001, 294(5543): 793- 795.
[3] Sparks T H, Jeffree E P, Jeffree C E. An examination of the relationship between flowering times and temperature at the national scale using long-term phenological records from the UK. International Journal of Biometeorology, 2000, 44(2): 82- 87.
[4] 王连喜, 陈怀亮, 李琪, 余卫东. 植物物候与气候研究进展. 生态学报, 2010, 30(2): 447- 454.
[5] Sakai S, Momose K, Yumoto T, Nagamitsu T, Nagamasu H, Hamid A A, Nakashizuka T. Plant reproductive phenology over four years including an episode of general flowering in a lowland dipterocarp forest, Sarawak, Malaysia. American Journal of Botany, 1999, 86(10): 1414- 1436.
[6] 郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1584- 1587.
[7] 张福春. 气候变化对中国木本植物物候的可能影响. 地理学报, 1995, 50(5): 403- 408.
[8] Menzel A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Climatic Change, 2003, 57(3): 243- 263.
[9] Prieto P, Peuelas J, Ogaya R, Estiarte M. Precipitation-dependent flowering ofGlobulariaalypumandEricamultiflorain Mediterranean shrubland under experimental drought and warming, and its inter-annual variability. Annals of Botany, 2008, 102(2): 275- 285.
[10] 中国科学院华南植物研究所. 广东植物志 (第三卷). 广东科技出版社, 2005: 158- 159.
[11] 中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1990, 50(2): 128- 131.
[12] 刘淑菊, 郝清玉. 石梅湾青皮林群落结构及天然更新特征. 林业资源管理, 2011, (2): 54- 59.
[13] 汪永华, 胡玉佳, 翁应云. 海南岛青皮林自然保护区. 植物杂志, 2003, (5): 8- 8.
[14] 梁淑群, 林英, 杨小波, 黄世满, 符史新, 谢国干. 海南万宁礼纪青梅林. 海南大学学报: 自然科学版, 1993, 11(4): 1- 9.
[15] 国家林业局. LY/T 1687- 2007 热带森林生态系统定位观测指标体系. 北京: 中国标准出版社, 2007.
[16] Thórhallsdóttir T E. Flowering phenology in the central highland of Iceland and implications for climatic warming in the Arctic. Oecologia, 1998, 114(1): 43- 49.
[17] 肖宜安, 何平, 李晓红. 濒危植物长柄双花木开花物候与生殖特性. 生态学报, 2004, 24(1): 14- 21.
[18] Martinez-Palle E, Aronne G. Flower development and reproductive continuity in MediterraneanRuscusaculeatusL. (Liliaceae). Protoplasma, 1999, 208(1/4): 58- 64.
[19] 刘志民, 蒋德明. 植物生殖物候研究进展. 生态学报, 2007, 27(3): 1233- 1241.
[20] 徐雨晴, 陆佩玲, 于强. 气候变化对植物物候影响的研究进展. 资源科学, 2004, 26(1): 129- 137.
[21] 张学霞, 葛全胜, 郑景云, 张福春. 近150年北京春季物候对气候变化的响应. 中国农业气象, 2005, 26(3): 263- 267.
[22] 李荣平, 刘晓梅, 周广胜. 盘锦湿地芦苇物候特征及其对气候变化的响应. 气象与环境学报, 2006, 22(4): 30- 34.
[23] Stevenson P R, Castellanos M C, Cortés A I, Link A. Flowering patterns in a seasonal tropical lowland forest in Western Amazonia. Biotropica, 2008, 40(5): 559- 567.
[24] 刘文明, 宋学之. 青皮种子主要储藏条件的研究I. 种子含水量与测控. 林业科学研究, 1989, 2(3): 214- 220.
Reproductive phenological characteristics of hainan coastalVaticamangachapoiforest
LÜ Bing, WANG Na, LIU Shuju, HAO Qingyu*
MinistryofEducationKeyLaboratoryforTropicalAnimalandPlantEcology,CollegeofLifeSciences,HainanNormalUniversity,Haikou571158,China
Vaticamangachapoibelongs to theVaticaLinn genus of theDipterocarpaceaefamily and was once extensively distributed over Hainan Island, from low to medium altitude. It is now an endangered species and a Ⅱ grade key protected plant in China,due to over cutting for its valuable wood. It exhibits several excellent characteristics, such as strong adaptability to growth conditions, especially having the ability to colonize poor sandy beach or rocky areas, eventually easily forming a singleVaticamangachapoidominant community due to strong natural regeneration, ability to compete and strong wind resistance. Therefore, it is no surprise that theVaticamangachapoiforest is distributed over the sandy beach of Shimei Bay, Hainan Island, China, and is the only patch of natural forest growing on sandy beach environments in Hainan Island. Now it has become a significant model forest for the regeneration and transformation of manmade shelterbelts and as a scenic forest in the Shimei Bay tourism area in Hainan Island. The reproductive phenology study ofVaticamangachapoiforest will help to reveal the biological characteristics of Vatica mangachapoi that allows it to successfully settle and reproduce in arid coastal beach environments. Our primary objectives were to: (1) Investigate the biological characteristics ofVaticamangachapoireproductive phenology, and (2) analyze the response ofVaticamangachapoireproductive phenology to temperature changes in the coastalVaticamangachapoiforest of Shimei Bay, so as to supply a theoretical basis for the protection and management of theVaticamangachapoiforest and to predictVaticamangachapoifluorescence, formation of red seed calyx, seed rain dispersal periods, etc. The reproductive phonological observations were conducted on the three levels of population, individuals and twigs continuously for three years. Thirty individualVaticamangachapoiwere selected based on systematic sampling and sixty twigs were labeled with two twigs per individual. 3a meteorological data were obtained from Hainan meteorological Bureau to analyze the interannual phenophase variation. The results showed that the average reproductive phenophase, numbers of buds in a twig, seed setting rate of young seeds, and seed setting rate of mature seeds ofVaticamangachapoiwere 101d, 133.3, 42.4% and 4.4%, respectively. The population ofVaticamangachapoiwas of 1a continuous flowering type, but exhibited the obvious concentrated florescence with a bimodal. There were many flowers in the period of concentrated flowering and few in the non-concentrated flowering period; the flowering time exhibited an obvious asynchronicity among individuals and between twigs on each individual. There was a significant difference in the numbers of flowers and seeds between the harvest and lean years; the rainfall in one month before buds occurred had no obvious effect on the flowering phenophase, but the monthly average temperature did have an obvious effect. When the monthly average temperature in March and April in the month before buds appeared was higher than 24℃ and 26.4℃, respectively, the flowering phenophase progression was obvious, but the flowering phenophase was delayed when the temperature was lower. These results demonstrate that the response of the flowering phenophase progression and delay based on the rise and fall in temperature was nonlinear, and the bimodal phenophase and seed dispersal period were a benefit to successfulVaticamangachapoiseed germination and growth on the seasonal dry sandy beach.
CoastalVaticamangachapoiforest; reproductive phenology; Hainan Island
国家自然科学基金资助项目(31060092)
2013- 04- 01;
日期:2014- 03- 25
10.5846/stxb201304010572
*通讯作者Corresponding author.E-mail: hnhaoqy@126.com
吕冰,王娜,刘淑菊,郝清玉.海南海岸青皮林繁殖物候特征.生态学报,2015,35(2):416- 423.
Lü B, Wang N, Liu S J, Hao Q Y.Reproductive phenological characteristics of hainan coastalVaticamangachapoiforest.Acta Ecologica Sinica,2015,35(2):416- 423.
