鄱阳湖区稻田生境中灰鹤越冬行为的时间分配与觅食行为
2015-03-10蒋剑虹戴年华邵明勤黄志强
蒋剑虹,戴年华,邵明勤,黄志强,卢 萍
1 江西师范大学生命科学学院,南昌 330022 2 江西省科学院生物资源研究所,南昌 330096 3 江西省野生动植物保护管理局,南昌 330038
鄱阳湖区稻田生境中灰鹤越冬行为的时间分配与觅食行为
蒋剑虹1,2,戴年华2,*,邵明勤1,黄志强3,卢 萍2
1 江西师范大学生命科学学院,南昌 330022 2 江西省科学院生物资源研究所,南昌 330096 3 江西省野生动植物保护管理局,南昌 330038
2013年11月—2014年2月,采用瞬时扫描和焦点动物法研究了鄱阳湖区稻田生境中灰鹤越冬行为的时间分配、活动节律与觅食行为。结果显示,觅食(64.09%)行为所占比例最大,其次为警戒(15.97%)、飞行(8.67%)和修整(7.37%)行为。4种主要行为中,觅食行为时间分配随越冬前期(11月)、中期(12月—翌年1月)、后期(2月)逐渐增加,其余行为时间均逐渐减少。各环境因子对主要行为的影响存在显著交互效应,修整行为随各环境因子变化最为明显,日最低温度升高、日最高温度降低、日照长度增加及湿度降低都会使修整行为增加;日照长度增加和湿度降低时,觅食行为增加;日照长度增加时,警戒行为减少。环境因子对成鹤影响效果与总体相同。环境因子仅对幼鹤的觅食行为影响显著,即日照长度增加和湿度降低,幼鹤觅食行为增加。环境因子对行为的影响为非线性关系,致使其影响趋势在不同范围内有所变化。行为节律上,灰鹤昼间各时段觅食行为保持较高水平,觅食高峰出现在11:00—11:59和17:00—17:30。灰鹤觅食生境与其夜宿地分离,致其上午觅食高峰有所推后。幼鹤昼间各时段行为节律与成鹤有较大差异,且各时段觅食行为比例均高于成鹤。灰鹤越冬期在稻田生境的平均啄食频率为(32.06±0.47)次/min,平均步行频率为(6.55±0.35)步/min。啄食频率与步行频率呈极显著负相关。时段和集群类型对啄食频率的影响存在显著交互效应。稻田中食物资源的可利用性逐渐下降,灰鹤的啄食频率随时间逐渐降低,为保证越冬期间获取足够的能量供应,灰鹤采取逐渐增加步行频率和觅食时间的策略。有觅食间隔的抽样单元中,平均警戒次数为(1.37±0.04)次/单元,平均警戒持续时间为(6.02±0.37)s/单元。成鹤花费在警戒的时间多于幼鹤,家庭群中的个体警戒持续时间多于聚集群中的个体。
稻田生境;灰鹤;时间分配;觅食行为
动物时间分配与活动节律是动物行为学研究的重要内容[1]。鸟类的时间分配与活动节律直接与其新陈代谢与能量需求相关,鸟类会在不同栖息环境和天气状况(如温度、湿度、日出日落时间)改变其时间分配与活动节律,以适应不同环境条件下的能量需求[2- 5]。幼鸟因缺乏经验会表现出不同于成鸟的时间分配与活动节律[6- 8]。鸟类越冬期的主要行为一般与存活有关,觅食及其相关行为与存活密切相关,在鸟类越冬期通常占很大比例[2]。因此,了解物种的觅食对策及其影响因子对该物种保护与管理具有极其重要的理论和实践意义[9- 10]。鸟类的觅食行为会受食物资源丰富度、时段、年龄和人为干扰等多种因素的影响[7,11- 14]。最优取食理论表明,鸟类会在不同条件下采取特定的行为策略,以达到最佳的能量摄入效率[15]。
灰鹤Grusgrus是国家II级重点保护野生动物,列入《濒危野生动植物种国际贸易公约》 (CITES) 附录Ⅱ中[16]。湿地国际2006年估计全球灰鹤种群为360000—370000只,其中,中国大陆越冬种群为10000—12000只[17]。鄱阳湖地区为中国境内灰鹤重要的越冬栖息地,Li 等2011年冬季在鄱阳湖记录灰鹤8408只[18]。灰鹤冬季的觅食地主要为农田和湿地草滩,其食性受当地食物资源的影响会发生变化[17,19- 21]。鄱阳湖区近年来常见灰鹤由湖滩草洲转移至湖滨稻田中觅食收割后散落的谷物。灰鹤在自然湿地生境、玉米地等非稻田生境及笼养条件下时间分配及活动节律已有较多研究[11,16- 17,22- 25],但冬季稻田生境的食物资源特征、分布及变化规律与非稻田生境明显不同,灰鹤的行为分配模式及觅食行为必然发生较大变化。本文通过对鄱阳湖区稻田生境中灰鹤越冬行为的时间分配、行为节律和觅食行为进行研究,揭示越冬灰鹤在该生境条件下的行为模式、觅食行为特征和能量需求。本文还分析了环境因子(温度、日照长度、湿度)对灰鹤行为分配和觅食行为的影响,揭示灰鹤的生存对策,为该物种的保护管理提供科学的理论依据。
1 研究区域概述

图1 鄱阳湖调查区域示意图Fig.1 The survey area of Poyang Lake
鄱阳湖(115°49′—116°46′E,28°11′—29°51′N)位于长江中游和下游交界处,长江南岸,江西北部,是中国第一大淡水湖[26- 27]。鄱阳湖属典型的亚热带季风气候,夏季盛行偏南风,炎热多雨;冬季盛行偏北风,气温低而降雨少;年平均气温17.6℃,年平均降水量1450—1550mm,多集中于4—6月,年日照时数1258.8—1794.5 h,年平均相对湿度68%—74%[6,28]。鄱阳湖周边土地肥沃,拥有稻田面积为7.62×105hm2,是中国重要的商品粮基地之一[28]。鄱阳湖每年枯水期间广阔的洲滩次第露出,湿地植被开始发育,湖泊水深也开始变浅,为鄱阳湖的越冬候鸟提供了大量的食物和良好的越冬环境,吸引了白鹤G.leucogeranus、东方白鹳Ciconiaboyciana、白头鹤G.monacha、黑鹳Ciconianigra、白枕鹤G.vipio、灰鹤、小天鹅Cygnuscolumbianus和白额雁Anseralbifrons等在内的数十多万只水鸟来此越冬[18,29]。本次研究区域位于鄱阳湖东岸的鄱阳县,为笔者近年鄱阳湖水鸟调查时,稻田生境中灰鹤最多且集中的区域。调查点位于鄱阳县莲湖乡表恩村、茭溪村和向阳村附近的约400hm2的稻田内,与鄱阳湖冬候鸟越冬重要的栖息地之一——大莲子湖仅有围堤之隔(图1)。每年稻田收割后谷粒散落其间,吸引了大量灰鹤来此取食,调查期间该稻田最多可见1000 余只灰鹤栖息觅食,是鄱阳湖区稻田生境觅食的越冬灰鹤种群的典型样地。
2 研究方法
2.1 调查方法
2013年11月—2014年2月,借助SWAROVSKI (20—60×)单筒望远镜,采用瞬时扫描法对鄱阳湖稻田中灰鹤的行为进行观察,即每5min记录1次成鹤与幼鹤的行为类别[6]。数量较多时,随机记录10—15只个体行为。采用焦点动物法,以1min为1抽样单元,每隔5min抽样1次灰鹤的啄食频率(啄食次数/min)和步行频率(步数/min)[30]。由于灰鹤两次觅食的间隔期常伴随短时间警戒、修整和社会行为,将两次觅食之间发生的这些行为定义为觅食间隔行为。若长时间无觅食行为(>30s),则放弃。根据鄱阳县湖畔冬季日出日落时间的实际情况,观察时间为每天07:00—17:30。环境因子(日最高温度(℃)、日最低温度(℃)、日照长度(min)、湿度(%))直接从江西省气象台发布的气象信息读取。
2.2 集群、行为及越冬时期的分类与定义
本次记录了灰鹤家庭群和聚集群的觅食行为。其中,家庭群由亲鹤和幼鹤组成,分2成、1成1幼、2成1幼和2成2幼4种形式。个体数量大于或等于5的群体定义为聚集群[16,31]。越冬期分越冬前期(10—11月)、中期(12月—翌年1月)、后期(2月)[5]。将灰鹤越冬行为分为7类(表1)[5,17,32]。
2.3 数据处理
以各行为发生频次占总行为频次的百分比测度行为时间分配,以各时间段内各类行为发生频次占总行为频次的百分比来测度日活动节律,时间分配与活动节律均以每天平均值计算[32]。先用Kolmogorov-Smironov检验数据的正态性,若符合或能转换成正态分布的因素,则利用General Linear Model (GLM) 中的多因素方差分析(MANOVA)检验各环境因子间对主要行为分配和年龄组成、集群类型、越冬时期对啄食频率主效应及交互效应[32]。使用单因素方差分析(one-way ANOVA)检验越冬各时期主要行为分配的差异。不符合且不能转换成正态分布的因素,则使用 Mann-Whitney U 检验(双独立样本)或Kruskal-Wallis H 检验(多独立样本)[5]。利用偏相关分析环境因子(日最高温度、日最低温度、日照长度、湿度)对单一环境因子与其余因子在控制和不控制条件下单一环境因子对行为分配、啄食频率和步行频率的影响[4]。采用Spearman相关性检验(双侧)啄食频率与步行频率、啄食频率和步行频率与时间、平均警戒次数与平均警戒持续时间的相关性。采用回归分析中的曲线估计(Curve Estimation)建立环境因子与行为分配之间的回归模型。文中数据表示为平均值±标准误(x±SE),显著性水平设置为α=0.05。所有统计分析借助Excel 2007和SPSS19.0完成。

表1 灰鹤越冬行为分类与定义Table 1 Description of various behavior types of the common cranes during wintering period
3 结果
3.1 灰鹤越冬行为的时间分配
本研究共扫描1461次,15577只次,包括11272只次成鹤和4305只次幼鹤。觅食(64.09%)行为所占比例最大,其次为警戒(15.97%)、飞行(8.67%)和修整(7.37%)行为,其他各行为均不足3.00%。成幼鹤行为分配差异较大,成鹤的觅食(60.70%)和飞行(8.90%)行为低于幼鹤(70.21%、9.47%),成鹤的警戒(19.05%)和修整(7.43%)行为高于幼鹤(14.21%、7.16%)(图2)。
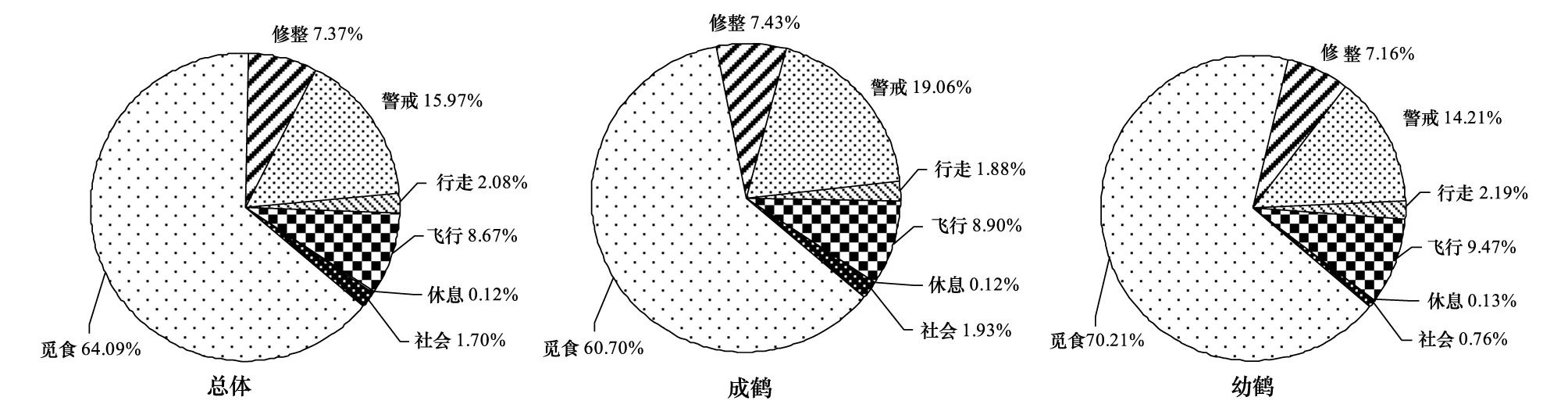
图2 鄱阳湖稻田灰鹤成幼鹤行为分配Fig.2 Time budgets of adult and juvenile common cranes inhabiting rice fields of Poyang Lake
4种主要行为中,觅食行为比例在越冬前期、中期、后期逐渐增加,修整、警戒和飞行行为比例则逐渐减少(图3)。觅食行为(F2,11=32.929,P=0.000)和飞行行为(F2,11=30.608,P=0.000)各时期差异极显著,修整行为各时期差异不显著(F2,11=2.160,P=0.162),警戒行为各时期差异显著(F2,11=4.697,P=0.034)。

图3 鄱阳湖稻田灰鹤4种主要行为在不同越冬期的变化 Fig.3 The variation of common crane′s four main behaviors during different wintering stages inhabiting rice fields of Poyang Lake
3.2 灰鹤主要行为与环境因子的关系
GLM检验显示,各环境因子对行为分配的影响均存在极显著交互效应(P=0.00)。偏相关分析显示,总体上,觅食行为与日照长度(r=0.635,P=0.036, df=9)呈显著正相关,与湿度(r= -0.659,P=0.027, df=9)呈显著负相关;警戒行为与日照长度(r= -0.605,P=0.049, df=9)呈显著负相关;修整行为与日最低温度(r=0.760,P=0.007, df=9)、和湿度(r=0.808,P=0.003, df=9)呈极显著正相关,与日最高温度(r=0.603,P=0.050, df=9)呈显著负相关,与日照长度(r= -0.793,P=0.004, df=9)呈极显著负相关。其它行为与环境因子相关性不显著。
成鹤的觅食行为与日照长度(r=0.709,P=0.014, df=9)呈显著正相关,与湿度(r= -0.688,P=0.019, df=9)呈显著负相关;警戒行为与日照长度(r= -0.778,P=0.005, df=9)呈极显著负相关;修整行为与日最低温度(r=0.733,P=0.010, df=9)呈显著正相关,与日最高温度(r= -0.669,P=0.024, df=9)呈显著负相关,与日照长度(r= -0.755,P=0.007, df=9)呈极显著负相关,与湿度(r=0.776,P=0.005, df=9)呈极显著正相关。其它行为与环境因子相关性不显著(表2)。
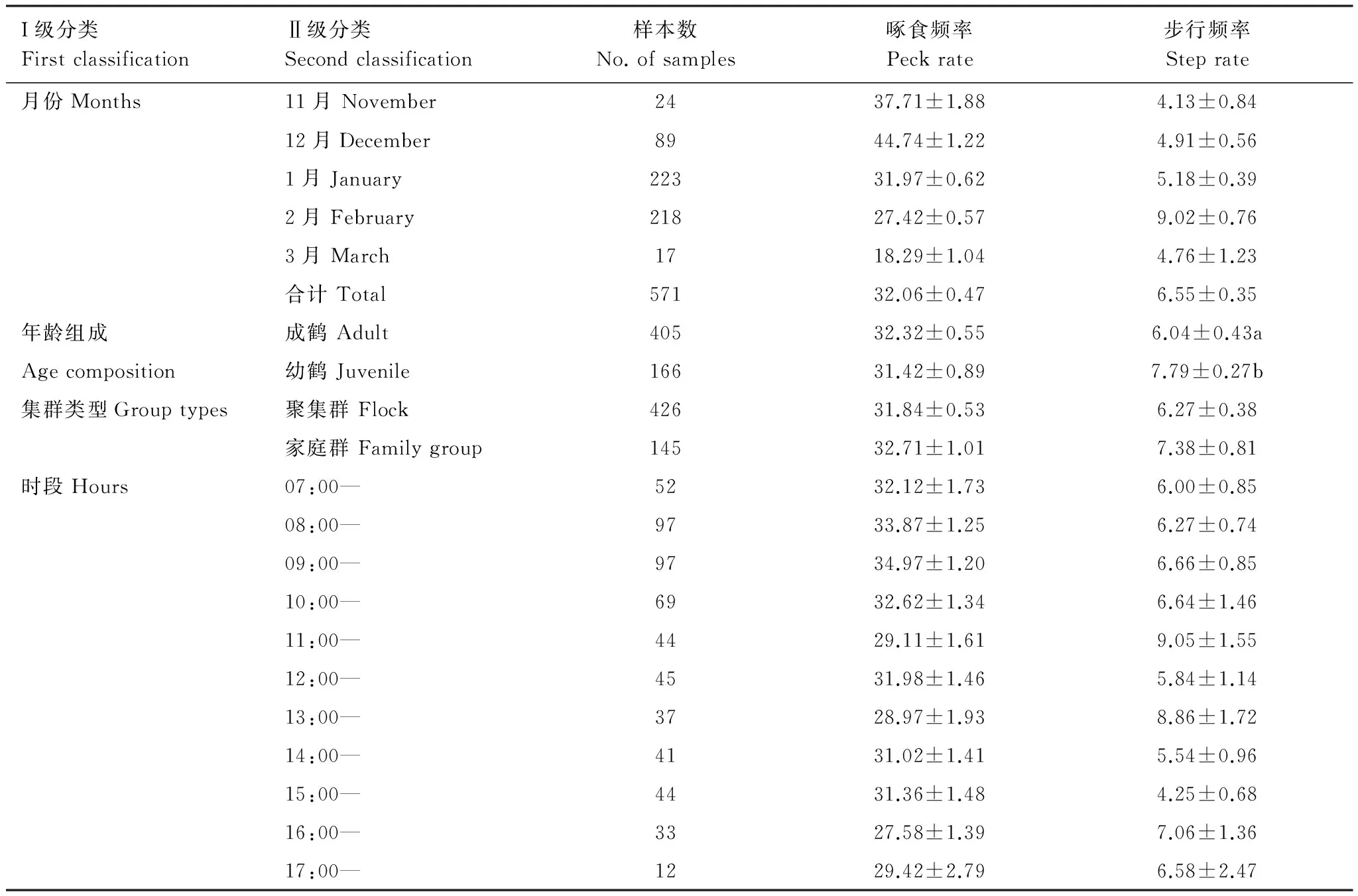
表2 鄱阳湖稻田灰鹤的啄食频率与步行频率Table 2 The peck and step rates of common crane inhabiting rice fields of Poyang Lake
幼鹤的觅食行为与日照长度(r=0.655,P=0.029, df=9)呈显著相关,与湿度(r= -0.724,P=0.012, df=9)呈显著负相关。其它行为与环境因子相关性不显著。
3.3 行为节律
本研究可用作行为节律分析的累计有效时间为14d(11月和12月份各4d, 1月和2月份各3d),总体上,灰鹤昼间各时段觅食行为保持较高水平,觅食高峰出现在11:00—11:59和17:00—17:30,分别为71.32%和70.58%,低谷出现在07:00—07:59,占54.74%;警戒行为昼间各时段波动不大,警戒高峰出现在07:00—07:59和16:00—16:59,分别为22.68%和21.77%,低谷出现在11:00—11:59,占12.58%;飞行行为高峰出现在07:00—07:59和10:00—10:59,分别为17.14%和11.14%;修整行为近似呈现“正态分布”,其最高峰出现在12:00—12:59,为10.29%,低谷出现在07:00—07:59和16:00—16:59,分别为2.24%和3.60%(图4)。
成鹤各时段行为节律的高峰与低谷出现时段和比例与总体状况相似。觅食行为2个高峰和低谷比例有所降低,分别为68.53%、69.19%和51.33%;修整行为高峰有所改变,出现在08:00—09:59和12:00—12:59(图5)。

图4 鄱阳湖稻田灰鹤总体行为节律 Fig.4 The overall behavior rhythm of common crane inhabiting rice fields of Poyang Lake

图5 鄱阳湖稻田灰鹤成鸟行为节律 Fig.5 The behavior rhythm of adult common crane inhabiting rice fields of Poyang Lake
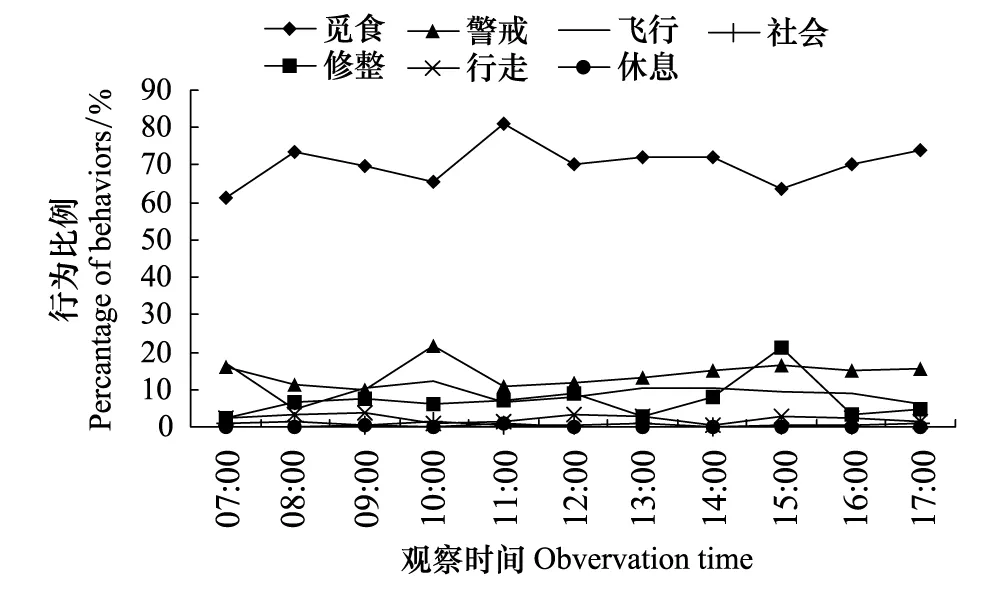
图6 鄱阳湖稻田灰鹤幼鸟行为节律 Fig.6 The behavior rhythm of juvenile common crane inhabiting rice fields of Poyang Lake
幼鹤各时段的觅食行为比例均高于成鹤,高峰和低谷出现时段与成鹤相同,高峰时占80.89%和73.95%,低谷时占61.06%;警戒行为昼间各时段波动较成鹤大,警戒高峰出现在07:00—07:59、10:00—10:59和16:00—16:59,分别为16.13%、21.90%和16.69%,低谷出现在09:00—09:59和11:00—11:59,分别为9.98%和10.78%;飞行行为与成鹤相似;修整行为高峰出现在15:00—15:59,为21.07%,低谷出现在07:00—07:59、13:00—13:59和16:00—16:59(图6)。
3.4 觅食行为
3.4.1 觅食频率与步行频率
共观察稻田灰鹤越冬觅食行为数据571min,灰鹤平均啄食频率为(32.06±0.47)次/min。GLM分析显示,时段和集群类型对啄食频率的影响存在显著交互效应(F=2.037,P=0.034)。啄食频率与时间呈极显著负相关(r= -0.754,P=0.002,N=14)(图7)。成鹤啄食频率((32.32±0.55) 次/min)略高于幼鹤((31.42±0.89) 次/min),但差异不显著(F1,569=2.541,P=0.112)。聚集群个体啄食频率((31.84±0.53) 次/min)略低于家庭群((32.71±1.01) 次/min),差异也不显著(F1,569=0.462,P=0.497)。各时段间啄食频率差异显著(F10,560=2.337,P=0.011),在08:00—08:59、09:00—09:59时段最高,分别达到(33.87±1.25)和(34.97±1.20)次/min,在16:00—16:59最低,为(27.58±1.39)次/min(表3)。偏相关分析显示,啄食频率与湿度(r= -0.685,P=0.020,df=9)呈显著负相关,其它环境因子与啄食频率和步行频率无显著相关性(P>0.05)。
稻田灰鹤越冬期平均步行频率为(6.55±0.35)步/min。啄食频率与步行频率呈极显著负相关(r=0.360,P=0.000,N=571)(图8)。步行频率与时间相关虽不显著(r=0.521,P=0.056,N=14),但随时间有一定的上升趋势(图7)。由于步行频率不符合正态分布,未考虑交互效应的影响,成鹤步行频率((6.04±0.43) 步/min)极显著低于幼鹤((7.79±0.27) 步/min)(Z= -3.666,P=0.000)。灰鹤在聚集群((6.27±0.38) 次/min)中的步行频率低于家庭群((7.38±0.81) 步/min),不同集群类型步行频率(Z= -1.556,P=0.120)和各时段间步行频率(χ2=10.086,P=0.433,df=10)差异均不显著(表2)。

图7 鄱阳湖稻田灰鹤啄食频率和步行频率的时间变化Fig.7 The temporal variation of peck and step rates for common crane inhabiting rice fields of Poyang Lake

图8 鄱阳湖稻田灰鹤啄食频率与步行频率相关分析 Fig.8 Correlation between peck and step rates of common crane inhabiting rice fields of Poyang Lake
3.4.2 觅食间隔时间与警戒
灰鹤觅食间隔中有短时间的警戒、修整及社会行为,其中警戒行为频次占95.56%。有些抽样单元灰鹤一直觅食,即无觅食间隔。有觅食间隔的抽样单元占总抽样单元的49.56% (N=283),成鹤觅食间隔比例大于幼鹤,家庭群大于聚集群。有觅食间隔的抽样单元中,平均警戒次数为(1.37±0.04)次/单元,平均警戒持续时间为(6.02±0.37) s/单元。Spearman相关性检验显示,平均警戒次数与平均警戒持续时间呈极显著负相关(r= -0.444,P=0.000,N=283)。由于警戒次数和警戒持续时间不符合正态分布,未考虑交互效应,成幼鹤警戒次数差异显著(Z=-2.914,P=0.004),但平均警戒持续时间显著不差异(Z= -0.334,P=0.738)。不同集群类型间警戒次数差异不显著(Z= -1.528,P=0.126),但警戒持续时间差异显著(Z= -3.517,P=0.000)(表3)。
4 讨论
4.1 灰鹤越冬行为的时间分配与行为节律
鸟类行为时间分配是对当地环境条件的适应,也是影响动物行为的综合表现[33]。高原地区低温环境增加鹤类的能耗,迫使其增加取食时间以满足越冬期较大的能量需求。丹顶鹤G.japonensis在人工湿地花费的警戒持续时间较原始滩涂高出近1倍[3]。本研究中灰鹤觅食(64.09%)行为比例低于江苏盐城保护区的灰鹤(70.3%)、云南拉市海(高原湖泊生境)的灰鹤(75.53%)和西藏南部(高原湖泊及农田生境)的黑颈鹤G.nigricollis(73.0%),与安徽升金湖(含农田生境)(60.4%)及上海崇明东滩(自然滩涂生境)的白头鹤(67.42%)相似,除盐城保护区的灰鹤(28.1%)外各次调查的警戒行为比例较为相似(14%—16%)[5,16- 17,34- 35]。Jesús等研究发现,西班牙灰鹤从11月至翌年2月,警戒持续时间增加、觅食时间减少[36]。Alonso等也发现越冬期间灰鹤的觅食时间逐渐下降,这与食物可利用性有关[7]。本次研究中,灰鹤越冬期的觅食行为比例则逐渐增加,这与Zhou等的结果相似[5]。主要因为(1)本次研究的稻田秋季(11月)收割后,散落的稻谷丰富度随着灰鹤、放牧家鸭等的取食逐渐下降;(2)降雨使原本在地表的稻谷埋入土中,灰鹤需要更多的时间来翻找食物,为了满足足够的能量需求,必须增加觅食时间以弥补觅食效率的降低;(3)刚到越冬地的越冬前期需要花费更多的警戒持续时间以适应不熟悉的环境,越冬后期需要增加取食以积累足够的能量用于迁飞和繁殖。
表3 鄱阳湖稻田灰鹤觅食间隔中的警戒行为与持续时间
Table 3 Alerting behavior and its duration between two consecutive foraging behaviors of common crane inhabiting rice fields of Poyang Lake

*仅指有间隔的抽样单元
鹤类觅食行为常在上午及下午出现一个高峰[11,16,34- 35]。本研究中灰鹤觅食高峰发生在11:00—11:59,这可能与灰鹤的夜宿地和觅食地分离有关,灰鹤常在07:00—07:59和10:00—10:59时段飞来此处觅食,故上午觅食高峰有所推后,拉市海越冬的灰鹤甚至表现出3个觅食高峰[11,16]。Alonso等研究发现,灰鹤上午比下午有更大的活动性[11]。若将灰鹤的觅食、行走、飞行、社会行为归为活动性行为,则此次研究中上午(80.86%)和下午(79.54%)的活动性相似[37]。
4.2 环境因子对稻田生境中灰鹤越冬行为的影响
鹤类的越冬行为具有一定的基本规律,但常受气候因子、取食条件、水源及安全等因素的制约,在一定的时空范围内产生相应的适应性变化[3- 4,26,34]。此次调查发现,修整、觅食、警戒行为会随多种或一种环境因子变化。笼养灰鹤在繁殖期也表现出理羽时间受天气状况影响显著的现象[23]。孔德军等研究发现,气温高时黑颈鹤增加修整时间减少取食时间,气温低时则减少修整时间增加取食时间[4]。白头鹤在寒冷及日照时间短时,可能会产生增加觅食时间,提高食物摄人量以弥补因抵御寒冷而导致的能量消耗的行为适应现象[34]。但此次研究发现,温度并没有显著影响灰鹤的觅食行为时间分配,日照长度对灰鹤的影响与白头鹤相反。这可能由于各研究中温度的高低、日照的长短并不在同一个范围内且温度和日照长度对行为的影响并不一定呈线性关系,觅食行为时间分配与日照长度存在极显著的二次曲线关系(y=117.644-0.362x+2.79×10-4x2;R2=0.891,F=44.765,P=0.000;y为觅食时间分配比例/%,x为日照长度/min),与日最高温度存在显著三次曲线关系(y=0.286+0.01x2+5.7×10-4x3;R2=0.587,F=7.812,P=0.008;y为觅食时间分配比例/%,x为日最高温度/℃)。由于为非线性关系,日照长度/日最高温度对觅食行为的影响为先减少/增加后增加/减少的趋势。这可能是环境因子对不同的物种影响效果不同,这些内容还有待进一步系统研究。
4.3 觅食行为
此次调查越冬灰鹤啄食频率与日本八代越冬白头鹤的啄食频率相似(约30 次/min),远高于在崇明东滩自然滩涂环境的越冬白头鹤(约5次/min)[34]。此次调查中啄食频率随着时间的推移逐步下降,而步行频率逐步上升,觅食时间分配逐渐增多。与Alonso等研究发现越冬期灰鹤觅食速率不断降低结果一致[7]。这表明随着时间的推移,食物资源的可用性逐渐降低,致其啄食频率不断降低。最优取食理论表明鸟类会在不同条件下采取不同的行为适应以达到最佳的能量摄入效率,由于单位面积的食物丰富度下降,灰鹤必须提高步行频率在更大面积的区域找到足够的食物资源,同时增加觅食时间,弥补觅食效率的下降[15]。12月的啄食频率略高于11月(表2),这可能是灰鹤为了适应飞翔,在迁徙过程中部分消化器官萎缩,刚到达越冬地消化器官尚未恢复,食物的消化和营养物质的吸收都受到了限制,其代谢能的摄入速率较低[38- 39]。因此11月份灰鹤刚到达越冬地的啄食频率并未达到最大值,随着消化器官的恢复,12月份啄食频率有所上升。
4.4 成幼鹤及不同集群的行为差异
幼鹤由于缺乏经验,一般需要亲代抚育以提高其存活率,并增加取食时间获得足够的食物摄取率[7- 8]。此次研究发现,幼鹤比成鹤花费更多的时间取食和更少的时间警戒,与多数研究相似[7- 8,17,32]。越冬期间,家庭群中的个体通常比聚集群中的个体利用更少的时间取食和更多时间警戒,说明较大的群体规模可以降低个体面临的捕食风险[16]。从取食间隔中的警戒次数与警戒持续时间看,成鹤花费在警戒的时间多于幼鹤,在家庭群中的个体警戒持续时间多于在聚集群中的个体,与以往研究结果相符[5,7- 8,22,32,36]。
本次调查时间为1个越冬期,但本文的很多结果均与其他研究结果类似,如灰鹤高的觅食时间和15%左右的警戒时间[5,16- 17,34- 35];越冬期间灰鹤的觅食时间逐渐增加[5];上午和下午各出现一个觅食高峰[11,16,34- 35];成幼鹤的取食和警戒时间差异[7- 8,17,32]等。因此,本文的结果具有普遍意义和科学价值。此外,越冬水鸟的行为研究时间一般均为1个越冬期[2,4- 6,32,37]。
4.5 保护建议
(1)鄱阳湖滨稻田为灰鹤等候鸟提供了重要的越冬觅食地和食物来源,应重视并加强对稻田生境鹤类的保护和管理,规范滨湖地区稻田的农药使用和尽量减少越冬期人为活动(放牛、牧鸭)对鹤类的干扰。可在灰鹤经常活动的区域划规范放牛和牧鸭生产(尤其是取食高峰的11:00—11:59和17:00—17:30),以减少人为干扰对越冬灰鹤行为的影响,也可考虑在稻田中预留小部分稻谷不收割,以保证越冬后期的食物供应;(2)深入研究环境因子对灰鹤行为的影响,理清环境因子变化对灰鹤行为的影响模式,可尝试系统收集行为数据后,建立该地区环境因子对行为分配的影响模型,预测环境变化对灰鹤生存状况的影响,为灰鹤和其他鹤类的保护提供参考依据。
[1] Halle S, Stenseth N C. Activity Patterns in Small Mammals: An Ecological Approach. Berlin: Springer, 2000.
[2] 曾宾宾, 邵明勤, 赖宏清, 蒋剑虹, 李畅, 戴年华. 性别和温度对中华秋沙鸭越冬行为的影响. 生态学报, 2013, 33(12): 3712- 3721.
[3] 吕士成, 陈卫华. 环境因素对丹顶鹤越冬行为的影响. 野生动物, 2006, 27(6): 18- 20.
[4] 孔德军, 杨晓君, 钟兴耀, 道美标, 朱勇. 云南大山包黑颈鹤日间越冬时间分配和活动节律. 动物学研究, 2008, 29(2): 195- 202.
[5] Zhou B, Zhou L Z, Chen J Y, Cheng Y Q, Xu W B. Diurnal time-activity budgets of wintering hooded cranes (Grusmonacha) in Shengjin Lake, China. Waterbirds, 2010, 33(1): 110- 115.
[6] 戴年华, 邵明勤, 蒋丽红, 杜建华, 蒋剑虹, 曾宾宾. 鄱阳湖小天鹅越冬种群数量与行为学特征. 生态学报, 2013, 33(18): 5768- 5776.
[7] Alonso J A, Alonso J C. Age-related differences in time budgets and parental care in wintering common cranes. The Auk, 1993, 110(1): 78- 88.
[8] Avilés J M. Time budget and habitat use of the Common Crane wintering in dehesas of southwestern Spain. Canadian Journal of Zoology, 2003, 81(7): 1233- 1238.
[9] Žydelis R, Esler D. Response of wintering Steller′s Eiders to herring spawn. Waterbirds, 2005, 28(3): 344- 350.
[10] Lok E K, Esler D, Takekawa J Y, de la Cruz S W, Sean Boyd W, Nysewander D R, Evenson J R, Ward D H. Stopover habitats of spring migrating surf scoters in southeast Alaska. The Journal of Wildlife Management, 2011, 75(1): 92- 100.
[11] Alonso J C, Alonso J A. Daily activity and intake rate patterns of wintering common cranesGrusgrus. Ardea, 1992, 80(3): 343- 351.
[12] Tatu K S, Anderson J T, Hindman L J, Hindman L. J, Seidel G. Diurnal foraging activities of mute swans in Chesapeake Bay, Maryland. Waterbirds, 2007, 30(1): 121- 128.
[13] Lee S D, Jaboński P G, Higuchi H. Winter foraging of threatened cranes in the demilitarized zone of Korea: behavioral evidence for the conservation importance of unplowed rice fields. Biological Conservation, 2007, 138(1/2): 286- 289.
[14] Henson P, Grant T A. The effects of human disturbance on trumpeter swan breeding behavior. Wildlife Society Bulletin, 1991, 19(3): 248- 257.
[15] Ravn Merkel F, Mosbech A. Diurnal and nocturnal feeding strategies in common eiders. Waterbirds, 2008, 31(4): 580- 586.
[16] 李学友, 杨洋, 杨士剑, 江望高, 彭贵鸿. 云南拉市海灰鹤的越冬行为初步观察. 动物学杂志, 2008, 43(3): 65- 70.
[17] 李忠秋, 王智, 葛晨. 盐城灰鹤 (Grusgrus) 越冬种群动态及行为观察. 动物学研究, 2013, 34(5): 453- 458.
[18] Li F S, Wu J D, Harris J, Burnham J. Number and distribution of cranes wintering at Poyang Lake, China during 2011- 2012. Chinese Birds, 2012, 3(3): 180- 190.
[19] Wood C, Qiao Y, Li P, Ding P, Lu B Z, Xi Y M. Implications of rice agriculture for wild birds in China. Waterbirds, 2010, 33(S1): 30- 43.
[20] 战永佳. 北京野鸭湖湿地自然保护区灰鹤(Grusgrus)的越冬生态研究. 北京:首都师范大学, 2007.
[21] 战永佳, 陈卫, 胡东, 吴秀山, 张金国. 北京湿地越冬灰鹤食性的初步分析. 湿地科学, 2007, 5(1): 45- 50.
[22] 陈文华, 杨洋, 江望高, 杨士剑, 彭贵鸿, 黄庭发. 云南丽江拉市海越冬前、中期灰鹤(Grusgrus)行为初步研究. 云南大学学报: 自然科学版, 2006, 28(S1): 365- 370.
[23] 刘丹, 何相宝, 赵锦霞, 田秀华. 笼养灰鹤繁殖期行为时间分配的研究. 野生动物杂志, 2009, 30(4): 185- 189.
[24] 周同山, 马雪峰, 卢小琴, 田秀华. 笼养灰鹤越冬期行为活动的时间分配和活动节律. 野生动物杂志, 2009, 30(5): 245- 247.
[25] 黄丙辉. 黄河湿地灰鹤越冬集群和觅食地生境特征的研究. 开封: 河南大学, 2013.
[26] 李言阔, 钱法文, 单继红, 李佳, 袁芳凯, 缪泸君, 谢光勇.气候变化对鄱阳湖白鹤越冬种群数量变化的影响. 生态学报, 2014, 34(10): 2645- 2653.
[27] Wang H Z, Xu Q Q, Cui Y D, Liang Y L. Macrozoobenthic community of Poyang Lake, the largest freshwater lake of China, in the Yangtze floodplain. Limnology, 2007, 8(1): 65- 71.
[28] 熊传伟, 黄国勤, 吴孙娟. 鄱阳湖及周边经济区稻田耕作制度的现状调查及综合评价——以余江县为例. 耕作与栽培, 2007, (1): 1- 2.
[29] 崔鹏, 夏少霞, 刘观华, 吴建东, 曾南京, 伍旭东, 文思标, 罗盛金, 纪伟涛, 雷富民. 鄱阳湖越冬水鸟种群变化动态. 四川动物, 2013, 32(2): 292- 296.
[30] 杨秀丽. 安徽升金湖国家级自然保护区白额雁(Anseralbiforns)数量分布,觅食行为和食性变化研究. 合肥: 中国科学技术大学, 2011.
[31] 刘强, 杨晓君, 朱建国, 赵健林, 余红忠. 云南省纳帕海自然保护区越冬黑颈鹤的集群特征. 动物学研究, 2008, 29(5): 553- 560.
[32] 袁芳凯, 李言阔, 李凤山, 李佳, 缪泸君, 谢光勇. 年龄、集群、生境及天气对鄱阳湖白鹤越冬期日间行为模式的影响. 生态学报, 2014, 34(10): 2608- 2616.
[33] 王有辉, 王虹. 中国灰鹤的分布与数量现状. 四川动物, 2003, 22(1): 35- 38.
[34] 崔志兴, 司强. 白头鹤越冬期日间行为初探. 上海师范大学学报: 自然科学版, 2004, 49(S1): 117- 121.
[35] 仓决卓玛, 杨乐, 李建川. 西藏黑颈鹤越冬期昼间行为的时间分配. 野生动物, 2008, 29(1): 15- 20.
[36] Avilés J M, Bednekoff P A. How do vigilance and feeding by common cranesGrusgrusdepend on age, habitat, and flock size?. Journal of Avian Biology, 2007, 38(6): 690- 697.
[37] 桑莉莉, 葛振鸣, 裴恩乐, 徐骁俊, 姜姗, 王天厚. 崇明东滩人工湿地越冬水禽行为观察. 生态学杂志, 2008, 27(6): 940- 945.
[38] Lindström Å, Klaassen M, Kvist A. Variation in energy intake and basal metabolic rate of a bird migrating in a wind tunnel. Functional Ecology, 1999, 13(3): 352- 359.
[39] Klaassen M, Biebach H. Energetics of fattening and starvation in the long-distance migratory garden warbler, Sylvia borin, during the migratory phase. Journal of Comparative Physiology B, 1994, 164(5): 362- 371.
Time budget and foraging behavior of wintering common cranes inhabiting rice fields of Poyang Lake
JIANG Jianhong1,2, DAI Nianhua2,*, SHAO Mingqin1, HUANG Zhiqiang3, LU Ping2
1CollegeofLifeScience,JiangxiNormalUniversity,Nanchang330022,China2InstituteofBiologicalResources,JiangxiAcademyofSciences,Nanchang330096,China3JiangxiProvincialManagementBureauofWildFaunaandFloraConservation,Nanchang330038,China
The common craneGrusgrusis currently listed in the Second Category of National Key Protected Wildlife Species in China and the second appendix of CITES with a global population size estimated at 360000—370000 individuals. In China there is a wintering population of 10000—12000 individuals, of which 8408 individuals are found at Poyang Lake, which represents an important wintering habitat for the species in China. From November 2013 to February 2014, the wintering time budget and foraging behavior of common cranes inhabiting rice fields of Poyang Lake were observed by instantaneous scan and animal focal sampling methods. Common cranes spent most of their time foraging (64.09%), followed by alerting (15.97%), flying (8.67%) and maintenance (7.37%) behaviors. While foraging behavior increased over early, middle and late wintering stages, the other behaviors gradually declined over the same time period. There was a significant interaction effect among environmental factors on behaviors. Maintenance behavior changed most obviously with environmental factors. For example, an increase in maintenance was observed when daily minimum temperature increased, daily maximum temperature decreased, day length increased and humidity decreased. Foraging behavior increased when day length increased and humidity decreased; and alerting behavior increased when day length increased and humidity decreased. The effect of environmental factors on adult individuals was the same as the overall pattern. There was only one significant impact on the foraging behavior of juveniles. In other words, when day length increased and humidity decreased, the foraging behavior of juveniles increased. The impact of environmental factors on behavior may be a nonlinear relationship that results in the influence trends changing in different ranges. In terms of the activity rhythms, foraging behavior was always kept at a high level in each daytime period, peaking at 11:00—11:59 and 17:00—17:30. Because the foraging habitat and communal roosting habitat of common cranes were separated, the foraging peak in the morning was delayed. There was also a large difference in activity rhythms between adults and juveniles, with foraging behavior of juveniles being higher than adults in each observation period. Mean peck rate was (32.06± 0.47) pecks / min, mean step rate was (6.55± 0.35) steps / min, and there was a significant negative correlation between peck rate and step rate. A significant interaction effect between time and group types was found on peck rate. Because the availability of food resources in rice-field habitats decreases gradually over winter, the peck rate of cranes should also decrease gradually over time. To ensure adequate energy supply during the winter, it is likely that common cranes would change their foraging strategy by increasing the step rate and foraging time. Nearly (1.37±0.04) times and (6.02±0.37) s per unit with interval were used for alerting. Adults spend more time alerting than do juveniles and spend more time alerting when in family groups than in flocks. This paper also provides scientific suggestions for the conservation of this species.
rice fields; Common crane; time budget; foraging behavior
国家自然科学基金(31260517,31101651); 江西省科技重大专项(20114ABG01100- 1- 03- 4)
2014- 05- 04;
2014- 10- 29
10.5846/stxb201405040882
*通讯作者Corresponding author.E-mail: dainianhua@jxas.ac.cn; 1048362673@qq.com
蒋剑虹,戴年华,邵明勤,黄志强,卢萍.鄱阳湖区稻田生境中灰鹤越冬行为的时间分配与觅食行为.生态学报,2015,35(2):270- 279.
Jiang J H, Dai N H, Shao M Q, Huang Z Q, Lu P.Time budget and foraging behavior of wintering common cranes inhabiting rice fields of Poyang Lake.Acta Ecologica Sinica,2015,35(2):270- 279.
