不同生育期滇重楼光合日变化及光响应特征1)
2015-03-06高成杰刘方炎王海林张春华孙永玉李昆
高成杰 刘方炎 王海林 张春华 孙永玉 李昆
(中国林业科学研究院资源昆虫研究所,昆明,650224)
责任编辑:任 俐。
滇重楼(Pairs polyphylla var.yunnanensis)隶属延龄草科(Trilliaceae)重楼属(Paris L.)多年生草本植物,主要分布于我国西南部的云南、四川和贵州一带,主产于云南,多生长于海拔1 400~3 100 m的常绿阔叶林、云南松林、竹林、灌草丛坡背阴坡或阴湿山谷中[1]。其根茎具有清热解毒、消肿散瘀、凉肝定惊等功效,被2010年版《中华人民共和国药典》收载[2]。由于滇重楼药用价值高,全国的各大制药厂、医院、中药材交易市场对滇重楼的需求量有增无减,而主产区长期掠夺式的采挖,已使野生滇重楼资源日益减少、已近枯竭,现已列为云南省30种稀缺濒危天然药物之一[3]。从实施可持续发展战略和提高产业化生产的原料供应保障率的角度出发,进行滇重楼人工栽培是一条必然之路。近几年,研究人员开始注重滇重楼的引种驯化和栽培技术研究,一些学者研究了不同遮阴程度[4]、土壤类型及肥力[5-6]等条件下滇重楼的生长状况,并取得了一定进展,然而,滇重楼生理生态基础研究远落后于栽培技术等方面的研究。
光合作用是植物生产力构成的最主要因素,通过光合作用所积累的有机物占植物总干物质的90%~95%[7]。通过对植物光合作用的研究,有助于采取适当的栽培管理措施,从而提高经济作物产量。植物在不同年龄和生长阶段的光合生产力变化不同[8-10],如Biswas et al.[11]对不同生育期黑绿豆光合气体交换特征的研究发现,黑绿豆在花期表现出最大的光合速率和气孔导度以及较低的胞间CO2摩尔分数,而最大的呼吸速率出现在豆荚鼓粒期。而徐克章等[12]对不同生育期人参叶片光合作用变化的研究表明,尚未形成生殖器官的1年生人参净光合速率(Pn)在叶片完全展开后即达最大值,此后缓慢下降;2~6年生人参叶片完全展开后达第1个高峰,开花期略有下降,绿果期出现第2高峰。目前为止,关于滇重楼光合特性的研究仅见于苏文华等[13]在人工气候箱下,进行温度、湿度对光合速率的单因子影响研究,以及刘维暐等[14]对滇重楼光响应和CO2响应参数的研究,但这些研究多是对7年生以上进入生殖期的滇重楼叶片在1 a中的某一生长季节进行的相关研究,而幼龄期滇重楼以及成熟期滇重楼在1 a中不同生长发育阶段的光合作用变化还未见报道。对不同生育期滇重楼光合特性进行研究,有助于了解滇重楼整个生育期光合变化规律,从而对不同龄期滇重楼或在滇重楼不同生长季节采取合理的管理措施,提升滇重楼产量。基于此,文中以人工荫棚种植下滇重楼为研究对象,分别在6月(花期)、8月(果实膨大期)和9月(果实成熟期)份对幼龄期(3年生)和成熟期(8年生)滇重楼光合日变化和光响应特征进行测定,同时结合不同时期环境因子,探讨不同生育期滇重楼光合参数之间的差异以及光合作用与不同季节环境因子的关系,旨在为滇重楼的栽培管理和高效种植模式提供基础理论依据。
1 研究区概况
试验点位于云南省玉溪市新平县者竜乡(24°16'~24°20'N,101°19'~101°26'E),海拔985 m,年均气温18.5℃,无霜期312 d,年均降水量达1 800 mm,6—11月份雨量集中,占全年降水的89.1%。
2 材料与方法
试验材料为3年生幼龄期和8年生成熟期的滇重楼,种源来自云南省普洱市景东县哀牢山(海拔1 300~2 000 m)地区。滇重楼实生苗一般在生长6 a后开始开花结实,每年的11月中下旬倒苗后即进入越冬,翌年3月份萌动,5月份叶展平,6月份进入盛花期,高生长停止,8月份为果实膨大期,并在茎基前端形成白色越冬芽,9月份进入果实成熟期。
于2013年3月份选取大小均匀一致的3年生根茎和8年生根茎按照株行距15 cm×20 cm移栽于玉溪市新平县者竜乡。滇重楼所在荫棚均采用双层遮阴网进行遮阴,遮阴率为70%左右。移栽后不施肥,其后田间人工适时除草、浇水。分别于2013年6月下旬(花期)、8月下旬(果实膨大期)和9月下旬(果实成熟期)对幼龄期滇重楼叶片和成熟期滇重楼叶片与花萼的光合日变化、光响应特征及其环境因子日变化进行观测,并对数据进行统计分析。
光合作用日变化测定:在连续的晴天,选择长势良好的幼龄期和成熟期滇重楼各6株,每株随机选择1片叶(花萼),从北京时间08:00—18:00,每隔2 h,采用LI-6400便携式光合作用测定系统(Li-Cor,Lincolin,NE,USA)对滇重楼各项光合生理指标进行测定。每个时期均连续测定2 d,每片叶(花萼)重复3次,结果取平均值。分析测定指标为净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)、大气CO2摩尔分数(Ca)以及人工荫棚内大气温度(Ta)、相对湿度(RH)和光合有效辐射(PA,R)等环境因子。根据测定的气体交换指标计算叶片和花萼气孔限制值(Ls=1-Ci/Ca)和水分利用效率(WU,E)(WU,E=Pn/Tr)[15]。
光响应曲线测定:在北京时间09:00—11:00,用便携式光合作用测定系统LI-6400-02B叶室对幼龄期滇重楼叶片和成熟期滇重楼叶片与花萼进行光响应曲线的测定。测定时设空气流速为500μmol·s-1,设置恒定温度为25℃,相对湿度为70%左右[16],参比室CO2摩尔分数以环境CO2摩尔分数为准。光照梯度自高到低设定为1 500、1 200、1 000、800、600、400、200、100、50、20、0μmol·m-2·s-1,每个光强下控制测定时间为120 s。每个时期测定2 d,每次测定为3次重复,结果取平均值。光响应曲线的相关参数按照直角双曲线修正模型进行拟合[17],模型表达式为:
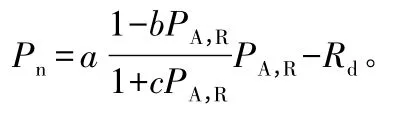
式中,Pn为净光合速率;PA,R为光合有效辐射;a为光响应曲线中的初始斜率,也称为初始量子效率;b为光抑制项;c为光饱和项;Rd为暗呼吸速率。由于该模型是一个具有极值的函数,因此,通过该模型可以直接求取光补偿点(LC,P)、光饱和点(LS,P)和最大净光合速率(Pnmax)。
数据处理:通过单因素方差分析(LSD法)比较不同季节幼龄期滇重楼叶片、成熟期滇重楼叶片和花萼光合生理指标差异显著性。所有数据统计和分析在SPSS 13.0和Excel 2003软件下完成。
3 结果与分析
3.1 环境因子日变化
滇重楼在不同生长季节所处环境因子日变化趋势基本一致(表1)。PA,R呈先升高后降低趋势,峰值在12:00—14:00出现,由大到小的顺序为8月份、9月份、6月份。6、8、9月份Ta从08:00一直上升到14:00时的最高值(分别为35.09、31.86、31.35℃),随后缓慢下降,08:00—18:00日均Ta值由大到小的顺序为6月份(31.00℃)、8月份(28.30℃)、9月份(27.40℃)。RH日变化基本与Ta呈相反趋势,从08:00的最高值持续下降,6月份RH最低值出现在12:00,而8、9月份分别出现在16:00和18:00,08:00—18:00日均RH由大到小的顺序为9月份(71.6%)、8月份(68.3%)、6月份(57.9%)。

表1 不同季节滇重楼所处环境因子日变化
3.2 滇重楼气孔气体交换日变化
3.2.1 成熟期滇重楼叶片气孔气体交换日变化
成熟期滇重楼叶片Pn日变化在6月份仅一个峰值,峰值出现在10:00左右;8、9月份则出现两个峰值,峰值分别出现在10:00和14:00,第一峰值明显高于第二峰值,在正午出现光合午休现象(表2)。同时,气孔导度(Gs)与Pn日变化趋势基本一致,在正午前较高,午后一直维持在较低水平。不同时期滇重楼叶片胞间CO2摩尔分数(Ci)均呈先下降后上升的趋势,6月份Ci最低值出现在16:00,8、9月份均出现在14:00;而气孔限制值(Ls)则与Ci变化呈相反趋势。Farquhar et al.[18]认为,Ci降低和Ls升高表明,Pn的降低主要是由气孔限制因素造成的;Ci升高和Ls降低则表明,Pn的降低主要是由气孔限制因素造成的。在6月份的10:00—16:00,滇重楼叶片Ci随Gs的下降而下降,Ls随Gs的下降而升高,该时段Pn的降低是由气孔限制因素造成的,在16:00以后,Pn的降低则是由非气孔限制因素造成的,即弱光引起的叶肉细胞活性的下降;而引起8、9月份叶片出现光合午休的主要因素则是气孔限制。6—9月份,滇重楼叶片Tr仅一个峰值,峰值分别出现在12:00和14:00,在8月份则出现两个峰值,峰值分别出现在10:00和14:00,与Gs日变化趋势一致。6月份,叶片WU,E峰值出现在16:00,而8、9月份均出现在10:00。
比较日均气体交换参数,成熟期滇重楼叶片在6、8、9月份日均Pn分别为(3.48±0.32)、(3.84±0.10)、(3.11±0.07)μmol·m-2·s-1,8月份的分别显著高于6、9月份的(P<0.05)。日均Tr以6月份的最高((1.40±0.12)mmol·m-2·s-1),其次是8月份的((1.35±0.08)mmol·m-2·s-1),9月份的最低((1.10±0.12)mmol·m-2·s-1)。日均WU,E以8月份((2.75±0.18)μmol·mmol-1)和9月份((2.76±0.14)μmol·mmol-1)显著高于6月份((2.07±0.20)μmol·mmol-1)的(P<0.05)。
3.2.2 幼龄期滇重楼叶片气孔气体交换日变化
幼龄期滇重楼并未进入生殖生长阶段,因此,其光合作用的主要器官仅为叶片,其Pn日变化在6月份仅一个峰值,峰值出现在10:00左右,在8、9月份出现两个峰值,峰值分别出现在10:00和14:00,正午出现光合“午休”现象(表2)。6月份,幼龄滇重楼叶片Gs在10:00最高,之后迅速下降到18:00的最低值;而8、9月份,Gs从10:00的最高值下降到12:00的最低值后又有升高的趋势。从Ci和Ls的变化趋势看,幼龄滇重楼叶片在6月份10:00—16:00Pn下降是由气孔关闭引起的,而在8、9月份14:00以后幼龄滇重楼叶片Ls随Ci的升高而降低,此期间Pn下降可能是由叶肉细胞羧化活性的降低所造成的。幼龄滇重楼叶片Tr日变化与Pn日变化趋势一致,在6月份仅一个峰值,峰值出现在Pn最高时段,在8、9月份则出现两个峰值,在光合出现“午休”时处于低谷。幼龄滇重楼叶片WU,E在不同生长时期均只出现一个峰值,与成熟滇重楼叶片类似,6月份峰值出现在16:00左右,8、9月份峰值出现在10:00左右。
比较日均气体交换参数,幼龄滇重楼叶片在6、8、9月份日均Pn分别为(3.39±0.31)、(3.71±0.17)、(2.53±0.14)μmol·m-2·s-1,6、8月份的分别显著高于9月份的(P<0.05)。日均WU,E分别为(2.50±0.30)、(2.43±0.17)、(2.79±0.26)μmol·mmol-1,差异均不显著。日均Tr以8月份的最高((1.66±0.12)mmol·m-2·s-1),其次是6月份的((1.40±0.10)mmol·m-2·s-1),9月份的最低((0.85±0.10)mmol·m-2·s-1),6、8月份的分别显著高于9月份的(P<0.05)。

表2 不同季节成熟期和幼龄期滇重楼叶片光合生理指标日变化
3.2.3 滇重楼花萼气孔气体交换日变化
成熟期滇重楼生殖生长可达5个月以上[1],在生长过程中,滇重楼花萼Pn日变化与叶片相似,6月份仅出现一个峰值,峰值出现在12:00;8、9月份均出现两个峰值,正午出现光合午休现象(表3)。滇重楼花萼在6月份午后Pn降低的生理因素与叶片类似,Ci在12:00—16:00随Gs的下降而下降,Ls随Gs的下降而升高,该时段Pn的降低是由气孔限制因素造成的,16:00以后花萼Pn下降的主要因素则是非气孔限制。8、9月份12:00左右Gs和Ci均降低,但Ls升高,表明花萼发生光合午休的生理因素为气孔限制,而14:00以后Pn的降低则可能是由于叶肉细胞活性下降所导致的。花萼Tr日变化在6月份的10:00达到最高值,随后一直下降;8、9月份出现两个峰值,峰值分别出现在10:00和14:00,与Pn日变化一致。花萼WU,E日变化在6月份出现两个峰值,峰值分别出现在12:00和16:00,而8、9月份均仅出现一个峰值,峰值出现在12:00。
比较日均气体交换参数,成熟滇重楼花萼在6、8、9月份日均Pn分别为(2.79±0.22)、(2.99±0.19)、(2.63±0.10)μmol·m-2·s-1,差异均不显著。日均Tr和WU,E均以8月份最高,其次是6月份,9月份最低。
3.3 不同生育期滇重楼光合速率对光强的响应
成熟期滇重楼叶片与花萼在6、8、9月份PA,RPn响应曲线呈相似的变化趋势(图1)。当成熟期滇重楼叶片PA,R为0~200μmol·m-2·s-1时,Pn随PA,R的增强呈线性增加;当PA,R为200~400μmol·m-2·s-1时,Pn随PA,R呈曲线缓慢上升;随着PA,R的继续增强,成熟期滇重楼叶片Pn不断缓慢升高,在光强达到1 500μmol·m-2·s-1时,也无明显的强光抑制现象。而滇重楼花萼Pn在PA,R达到800μmol·m-2·s-1时,开始缓慢下降,出现微弱的光抑制现象。不同生长季节幼龄滇重楼叶片在PA,R为0~200 μmol·m-2·s-1时,Pn差异并不明显,但随着PA,R的不断升高,不同生长季节幼龄期滇重楼叶片Pn出现差异,以8月份的高于6、9月份的,在光强达到1 500 μmol·m-2·s-1时,不同季节幼龄滇重楼叶片并无明显的强光抑制现象,与成熟期滇重楼叶片类似。

表3 不同季节滇重楼花萼光合生理指标日变化
通过直角双曲线修正模型对不同龄期滇重楼在不同季节光合作用的光响应曲线进行拟合,经F检验,相关性均达到显著水平(P<0.001),且R2>0.990(表4)。从不同季节来看,成熟期滇重楼叶片Pnmax以8、9月份的显著高于6月份的;而初始量子效率和LS,P均以9月份的显著高于6、8月份的(P<0.05);Rd和LC,P均以6月份的最高,其次是8月份的,9月份的最低(P<0.05)。不同季节花萼LS,P差异不显著(P>0.05),但Pnmax、LC,P和Rd均以9月份的显著高于6、8月份的,而初始量子效率则显著低于6、8月份的。幼龄期滇重楼叶片Pnmax在8月份的最高,其次是9月份的,在6月份的最低(P<0.05),而LS,P、LC,P和Rd则以8、9月份的大于6月份的,初始量子效率以6月份的显著大于8、9月份的(P<0.05)。
从不同龄期来看,幼龄期6、8月份的滇重楼叶片表观量子效率要高于成熟期叶片(P<0.05)的,而LC,P和Rd则低于成熟期叶片的,且在6月份和9月份达到显著水平(P<0.05)。不同季节幼龄期滇重楼叶片Pnmax和LS,P均低于成熟期叶片的。
滇重楼花萼具有同幼龄滇重楼叶片相似的光合性能,6、8月份的初始量子效率与幼龄期叶片的相差不大,但高于成熟期叶片的,9月份低于成熟期和幼龄期叶片的(P<0.05);不同季节花萼Pnmax和LS,P均低于成熟期叶片和幼龄期叶片的;LC,P和Rd与幼龄期叶片相差不大,但在6、8月份低于成熟期叶片的,9月份高于成熟期叶片的。

表4 不同季节滇重楼叶片与花萼光响应参数

图1 不同季节8年生(成熟期)与3年生(幼龄期)滇重楼光合-光响应曲线
4 结论与讨论
光合作用是植物物质积累与生理代谢的基本单元,与生长发育阶段和环境因子密切相关[9-10,19],研究表明,3年生幼龄期和8年生成熟期滇重楼叶片Pn变化日进程相似,但不同生长季节滇重楼叶片Pn变化日进程有差异,在6月份仅一个峰值,在8、9月份出现两个峰值,在中午出现“午休”现象。许大全[20]认为,当植物“午休”现象严重时,光合日进程中没有下午的第二个峰值出现,其峰值不是在中午,而是在上午的早些时候。幼龄期和成熟期滇重楼叶片Pn日进程在6月份并不同于典型的“单峰”型,其峰值出现在10:00左右,表明滇重楼在该时期“午休”现象严重。“午休”现象受生态因子、生理生化因子等多方面控制[21-22],滇重楼发生“午休”现象可能并不是由强光直接引起的,有研究表明,滇重楼最适光合有效辐射可达2 000μmol·m-2·s-1[13-14],本研究中,滇重楼叶片在高光强下也未出现光抑制现象,因此,高光强并不是直接导致滇重楼“午休”的直接原因,但它是引起一天中空气温度和湿度变化的主要因素[8]。苏文华等[13]通过人工气候箱对滇重楼光合作用的研究发现,滇重楼叶片光合速率在20~35℃下随温度升高而下降,本研究中,滇重楼出现光合“午休”时,所处Ta超过31℃,尤其是在6月份,Ta高达35℃,较高的温度可能影响光合作用中酶的活性,降低了光合效能[20]73。一些研究认为,较低的空气湿度也是导致光合“午休”的重要生态因子[21,23],滇重楼叶片在6月份“午休”现象严重,下午并未出现第二个峰值,而在8、9月份下午均有第二个峰值,这可能与8、9月份Ta下降,RH升高有关。
尽管幼龄期和成熟期滇重楼在不同生长季节都具有相似的光合日变化趋势,但不同龄期滇重楼日均气体交换参数有差异。成熟期滇重楼叶片在不同生长季节日均Pn和Pnmax均高于幼龄期叶片,尤其是6月份(花期)和9月份(果实成熟期)。已有研究表明,库的大小对源的活动具有调节作用,植物生殖器官的存在对叶片光合作用有促进作用,这种现象在一些植物中均存在,包括苹果[24]、柑橘[25]和桃[26]。成熟期滇重楼叶片光合速率高于幼龄期可能与库强有关,花和果实的存在需要消耗植物大量营养,从而导致成熟期滇重楼叶片具有较高的光合速率。从不同季节来看,成熟期和幼龄期滇重楼叶片日均Pn在8月份最大,其次是6月份,9月份最低,一方面可能是由于6月份较高的Ta和较低的空气湿度导致滇重楼在该时期出现严重的“午休”现象,而9月份的Ta虽较低,空气相对湿度也升高,但该时期滇重楼已逐渐进入衰老期,其光合作用中酶的活性可能下降,从而导致该时期较低的净光合速率。另一方面,8月份可能是滇重楼根茎产量形成的重要时期,植物在该时期通过较高的光合效率来达到根茎产量的形成[9]。许多研究表明,种子植物生育后期是光合同化物生产与积累的关键性阶段,然而,种子形成过程中伴随着植株的生理衰老[27-28],如花后高粱叶片光合速率呈现出先升高后下降的趋势[29],文中也得出类似的结论。但相较于幼龄期,成熟期滇重楼在9月份日均Pn下降并不明显,仍然具有较高的最大净光合速率,这有利于增加该时期滇重楼果实产量的积累。
滇重楼属耐阴植物,其叶片初始量子效率较高(0.051~0.079),光补偿点较低((3.25~13.10)μmol·m-2·s-1),对弱光的利用率较高,同时其光饱和点较高((918.07~1 440.97)μmol·m-2·s-1),利用的光合有效辐射范围很大,这与前人研究结果类似[13-14]。但不同龄期和不同季节滇重楼光响应参数并不一致,幼龄期滇重楼初始量子效率较高,且LC,P和LS,P均低于成熟期滇重楼的,对弱光的利用率更高,但对强光的利用能力不如成熟期滇重楼。因此,在滇重楼种植过程中,幼龄期滇重楼有必要采取较高遮阴度的遮阴网进行遮阴,或适当增加栽培密度。而对于8年生以上的成熟期滇重楼可适当降低遮阴度,使其获得更大的光合积累。苏文华等[13]认为,滇重楼光和作用的最佳条件是强光照和高湿度,但强光照往往引起生境温度升高及湿度下降,高温和低空气湿度对滇重楼叶片的光合作用不利。从不同季节来看,滇重楼9月份的LS,P高于8月份的,6月份最低,且在滇重楼生长后期,空气温度逐渐降低,相对湿度逐渐升高。因此,滇重楼在一年中的生长发育后期,可适当降低遮阴度,提高光照强度,使其获得更高的有机物积累。
高等植物制造光合产物的主要源器官除了绿色叶片外,几乎所有的营养器官(茎、根系)和繁殖器官(绿色花朵和发育中的果实)都可进行光合作用[30],许多研究中,植物花萼最大净光合速率要比叶片的低得多,如绿花嚏根草(Helleborus viridis)和黑嚏根草(H.niger)等[30-31]。本研究中,滇重楼花萼在6、8、9月份日均净光合速率分别为成熟期滇重楼叶片的80.2%、77.9%和84.6%,为幼龄期滇重楼叶片的82.3%、80.6%和104.0%,Pnmax分别为成熟期和幼龄期叶片的71.9%~82.9%和72.1%~99.6%,可见,滇重楼花萼具有同叶片类似的同化功能,尤其是幼龄期叶片。Yu et al.[32]对海拔2 600 m下滇重楼叶片与花萼光响应特征的研究结果表明,滇重楼在生长旺盛期,花萼与叶片光响应曲线几乎重叠,而本研究中,滇重楼花萼在不同季节光响应参数与叶片相差甚大,其Pnmax和LS,P均低于成熟期和幼龄期叶片,对弱光的利用效率较高,这可能与高海拔(2 600 m)和低海拔下(985 m)所处环境因子(如温度和光合有效辐射等)差异较大有关。
文中在引种栽培的基础上,对海拔985 m地区处于不同生育期的滇重楼光合日变化和光响应特征进行研究,认为该地区空气温度(Ta)偏高(尤其是6月份)引起滇重楼气孔关闭,是导致滇重楼光合“午休”的重要因素。滇重楼在8月份日均Pn最高,该时期可能是滇重楼根茎产量形成的重要时期。幼龄期滇重楼初始量子效率较高,且LC,P和LS,P均低于成熟期滇重楼,对弱光的利用率更高,但对强光的适应能力不如成熟期滇重楼。在滇重楼种植过程中,可将滇重楼分为幼龄期和成熟期,采取不同的遮阴度进行遮阴,成熟期滇重楼较幼龄期可适当降低遮阴度。另外,不同季节滇重楼光响应参数并不一致,9月份的LS,P高于8月份的,6月份的最低,在每年滇重楼生长后期,可适当降低遮阴度,提高光照强度,使其获得更大的光合积累。滇重楼药用部位主要为根茎,进一步的研究需要从光合产物分配的角度探讨不同生育期滇重楼各器官生物量和营养分配的动态变化,从而更好地指导滇重楼种植过程中的栽培管理。
[1]李恒.重楼属植物[M].北京:科学出版社,1998.
[2]国家药典委员会.中国药典[M].北京:中国医药科学技术出版社,2010.
[3]陆辉,许继宏,陈锐平,等.云南重楼属植物资源现状与保护对策[J].云南大学学报:自然科学版,2006,28(S1):307-310.
[4]陈翠,汤王外,谭敬菊,等.不同遮荫方式及遮荫率对滇重楼生长的影响研究[J].中国农学通报,2010,26(10):149-151.
[5]何忠俊,曾波,梁社往.锌对滇重楼生长、养分和总皂甙含量的影响[J].西南农业学报,2012,25(2):665-669.
[6]毛玉东,梁社往,何忠俊,等.土壤pH对滇重楼生长、养分含量和总皂甙含量的影响[J].西南农业学报,2011,24(3):985-989.
[7]夏方山,毛培胜,闫慧芳,等.植物花后光合性能与物质转运的研究进展[J].草地学报,2013,21(3):420-427.
[8]郭春燕,李晋川,岳建英,等.两种高质牧草不同生育期光合生理日变化及光响应特征[J].生态学报,2013,33(6):1751-1761.
[9]Hgaza V K,Diby L N,AkéS,et al.Leaf growth and photosynthetic capacity as affected by leaf position,plant nutritional status and growth stages in Dioscorea alata L.[J].Journal of Animal&Plant Sciences,2009,5(2):483-493.
[10]Sajjadinia A,Ershadi A,Hokmabadi H,et al.Gas exchange activities and relative water content at different fruit growth and developmental stages of ON and OFF cultivated pistachio tree[J].Australian Journal of Agricultural Engineering,2010,1(1):1-6.
[11]Biswas D K,Haque M M,Rahman M A.Photosynthetic gas exchange characteristics,leaf area and dry matter accumulation of two blackgram cultivars[J].Journal of Biological Science,2001,1(10):951-954.
[12]徐克章,张美善,武志海,等.人参不同生育期叶片光合作用变化的研究[J].作物学报,2006,32(10):1519-1524.
[13]苏文华,张光飞.滇重楼光合作用与环境因子的关系[J].云南大学学报:自然科学版,2003,25(6):545-548.
[14]刘维暐,陈翠,和荣华,等.四种重楼属植物光合作用特征[J].植物分类与资源学报,2013,35(5):594-600.
[15]Berry JA,Downton WJS.Environmental regulation of photosynthesis//[M].Govindjee.Photosynthesis(VolumeⅡ).New York:Academic Press,1982.
[16]张世华,梁社往,何忠俊,等.10个地理种源滇重楼光合特性的比较研究[J].云南农业大学学报,2012,27(5):708-715.
[17]Ye Z P.A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45(4):637-640.
[18]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33(1):317-345.
[19]王强,张其德,卢从明,等.超高产杂交稻不同生育期的光合色素含量、净光合速率和水分利用效率[J].植物生态学报,2002,26(6):647-651.
[20]许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[21]何文兴,易津,李洪梅.根茎禾草乳熟期净光合速率日变化的比较研究[J].应用生态学报,2004,15(2):205-209.
[22]Küppers M,Wheeler A M,Küppers B IL,et al.Carbon fixation in eucalypts in the field:analysis of diurnal variations in photosynthetic capacity[J].Oecologia,1986,70(2):273-282.
[23]Raschke K,Resemann A.The midday depression of CO2assimilation in leaves of Arbutus unedo L.:diurnal changes in photosynthetic capacity related to changes in temperature and humidity[J].Planta,1986,168(4):546-558.
[24]Palmer J W,Giuliani R,Adams H M.Effect of crop load on fruiting and leaf photosynthesis of‘Braeburn’/M.26 apple trees[J].Tree Physiol,1997,17(11):741-746.
[25]Iglesias D J,Lliso I,Tadeo F R,et al.Regulation of photosynthesis through source:sink imbalance in citrus is mediated by carbohydrate content in leaves[J].Physiol Plant,2002,116(4):563-572.
[26]Li Wei D,Duan Wei,Fan Pei G,et al.Photosynthesis in response to sink-source activity in relation to end products and activities of metabolic enzymes in peach trees[J].Tree Physiol,2007,27(9):1307-1318.
[27]Lu Congming,Lu Qingtao,Zhang Jianhua,et al.Characterization of photosynthetic pigment composition,photosystemⅡphotochemistry and thermal energy dissipation during leaf senescence of wheat plants grown in the field[J].Journal of Experimental Botany,2001,52:1805-1810.
[28]Yong I K,Nilda R B,Tay E H,et al.Antioxidative enzymes offer protection from chilling damage in rice plants[J].Crop Science,2003,43(6):2109-2117.
[29]周紫阳,黄瑞冬.不同籽粒淀粉含量高粱花后叶片光合指标的比较[J].东北农业大学学报,2011,42(4):52-56.
[30]Aschan G,Pfanz H.Non-foliar photosynthesis-a strategy of additional carbon acquisition[J].Flora,2003,198(2):81-97.
[31]Salopek-Sondi B,Kovac M,Ljubesic N,et al.Fruit initiation in Helleborus niger L.triggers chloroplast formation and photosynthesis in the perianth[J].Journal of Plant Physiology,2000,157(4):357-364.
[32]Yu Kun,Fan Qilong,Wang Yan,et al.Function of leafy sepals in Paris polyphylla:photosynthate allocation and partitioning to the fruit and rhizome[J].Function of Plant Biology,2013,40(4):393-399.
