象草和巨菌草的核型分析
2015-02-28朱丹丹王培丹林兴生苏德伟林占熺
朱丹丹, 王培丹, 林兴生, 林 辉, 苏德伟, 林占熺*
(1.福建农林大学 生命科学学院, 福建 福州 350002; 2.国家菌草工程技术研究中心, 福建 福州 350002)
象草和巨菌草的核型分析
朱丹丹1, 王培丹1, 林兴生2, 林 辉2, 苏德伟2, 林占熺2*
(1.福建农林大学 生命科学学院, 福建 福州 350002; 2.国家菌草工程技术研究中心, 福建 福州 350002)
为巨菌草的种质鉴定、遗传育种和亲缘关系提供一定的细胞学基础,采用传统压片法,对象草(Pennisetumpurpureumschumach)和巨菌草〔Pennisetumgiganteumz.x.lin(暂定名)〕的染色体数目及核型进行分析。结果表明:象草和巨菌草的染色体形态较为一致,大部分由中部(m)和近中部(sm)着丝粒的染色体组成,象草和巨菌草的核型公式分别为2 n=4 x=28=20 m+8 sm和2 n=4 x=28=16 m+8 sm+4 st;染色体相对长度组成分别为2 n=4 x=28=6 L+6 M2+10 M1+6 S和2 n=4 x=28=4 L+8 M2+10 M1+6 S,染色体基数和总数目一致,核型分类均为2 A型,不对称系数为58%~60%,均属于较原始的类型。结论:象草和巨菌草的的染色体无明显差异性,从细胞染色体水平上认为两者亲缘关系较近。
象草; 巨菌草; 染色体; 核型
在遗传多样性分析研究中,不仅有形态学分析、细胞学分析、分子标记等常规方法,同时还包括流式细胞技术和荧光原位杂交等新技术被广泛运用。植物染色体信息与植物的其他特性相比,不同的是其能保持一致性,因此在植物分类学、进化分析学和进化生物学方面是较为关键的参数[11]。目前,关于狼尾草属(Pennisetum)植物核型方面的研究已有报道,如周自玮等[12]研究了云南德宏地区逸生狼尾草(NaturalizedPennisetumsp.)的分类学鉴定,张伦等[13]对长序狼尾草(PennisetumlongissimumS.L.Chen et Y. X. Jin)的核型进行了分析。但前人的研究只是对狼尾草属植物染色体数目及基数的确定,并未对其染色体的着丝粒组成、染色体长度组成、核型类型划分等进行相关的细胞核型分析。目前,从国外引进的狼尾草属植物较多,常出现同名异物和同物异名的现象。因此,笔者以象草和巨菌草为试验材料,通过对其进行染色体数目、形态观察以及核型分析,深入探讨2种植物之间的亲缘关系,并为其遗传育种、引种以及巨菌草种的起源进化、系统分类等提供一定的细胞学理论依据。
1 材料与方法
1.1 材料
象草和巨菌种植于福建农林大学菌草研究所菌草圃。
1.2 材料处理
选取长势较好的象草和巨菌草,将其茎每2节截成段,浸入盛有清水的桶中培养生根,每2 d换1次清水,pH自然条件下培养。
1.3 细胞染色体制片
选取生长旺盛的根尖(1 cm左右),离体处理后即刻放入0.05%秋水仙素溶液中2 h,用卡诺氏固定液(V无水乙醇∶V乙酸=3∶1)固定24 h,预处理后将材料置1 mol/L盐酸常温解离10 min。水洗约10 min,转入改良苯酚品红染色液中染色1~2 h。去染液,用蒸馏水洗约30 min后,再用2%纤维素酶和果胶酶(质量比2∶1)混合液在37℃恒温箱中解离30~60 min。清水洗去酶液后,再用卡诺氏固定液固定10 min,最后用45%乙酸或稀改良苯酚品红染色液染色和压片[14]。
1.4 显微镜观察
在新时代发展期,以“软性协商”为核心的“软治理”方式可有效解决矛盾冲突,达成多方共赢的局面。对于城市社区而言,可应用调解、协商、讨论、引导、说服等灵活手段和较强的谈判指导功能,化解社会矛盾,实现城市社区的共同治理[27]。对于新农村社区而言,可在“软法和软权力”的基础上,利用心理疏导、人文关怀等“柔性”执法手段,实现乡村社会的治理有序[28]。而本文所提及的基于压力型体制形成的“不出事治理术”,其实即是新时期一种以“软手段”为实践表征的权宜性治理策略。
将象草和巨菌草根尖细胞临时装片,利用尼康NI-U型号显微镜观察,在低倍镜(100倍)下找到视野后用油镜(1000倍)观察,选择细胞分裂中期,染色体较粗、分散、数目全的染色体进行显微摄影,2种草各选择30个染色体分散良好、轮廓清晰、数目较全的细胞染色体中期相拍照。
1.5 核型分析及相关公式
分别选择30个象草和巨菌草细胞进行染色体数目计数,核型取30个细胞的平均值。核型分析按照李懋学与陈瑞阳[15]方法,染色体排序参照Levan等[16]提出的命名系统,染色体相对长度系数用Kuo等[17]的方法计算,核型不对称系数按Arano[18]提出的方法确定,核型分类按照Stebbins[19]的分类标准。利用核型分析软件Karyo3.1(美国东乐自然基因生命科学公司提供)处理图片、确定着丝粒位置及测量染色体长臂和短臂,统计染色体数目,计算下列值,排序和编号染色体,并划定染色体类型。相关公式如下:
臂比(r)=长臂(S)/短臂(L)
染色体相对长度=(染色体长度/染色体组总长度)×100%
染色体相对长度指数(I.R.L)=染色体长度/全组染色体平均长度
核型不对称系数(As.k, %)=(长臂总长/全组染色体总长)×100%
2 结果与分析
2.1 象草的细胞染色体
由图1可见,其中的细胞染色体数目为28条、26条和30条分别有25个、2个和3个细胞,细胞分别占计数总数的83.3%、7.7%和10%。根据核型分析的标准化建议[23],确定象草的染色体数目为2n=4x=28。
2.2 象草的核型
由表1和表2可知,象草染色体的数目为2 n=4 x=28,其中,第3号、第4号、第11号、第12号为近中部着丝粒染色体(sm),其余均为中部着丝粒染色体(m)。核型公式为2 n=4 x=28=20 m+8 sm。细胞染色体相对长度为3.31%~12.28%,其组成为2 n=4 x=28=6 L+6 M2+10 M1+6 S,最长和最短染色体比为3.71,染色体臂比大于2∶1的染色体占6.67%,按照Stebbins[19]提出的核型分类标准,象草的核型属于2A型,核型不对称系数为58.77%。

注:a为象草染色体中期相,b为象草核型图。
Note: a, Metaphase phase of chromosomes; b, chart of chromosome karyotype.
图1 象草染色体中期相和核型图
Fig.1 Metaphase chromosomes and karyotype ofP.purpureum
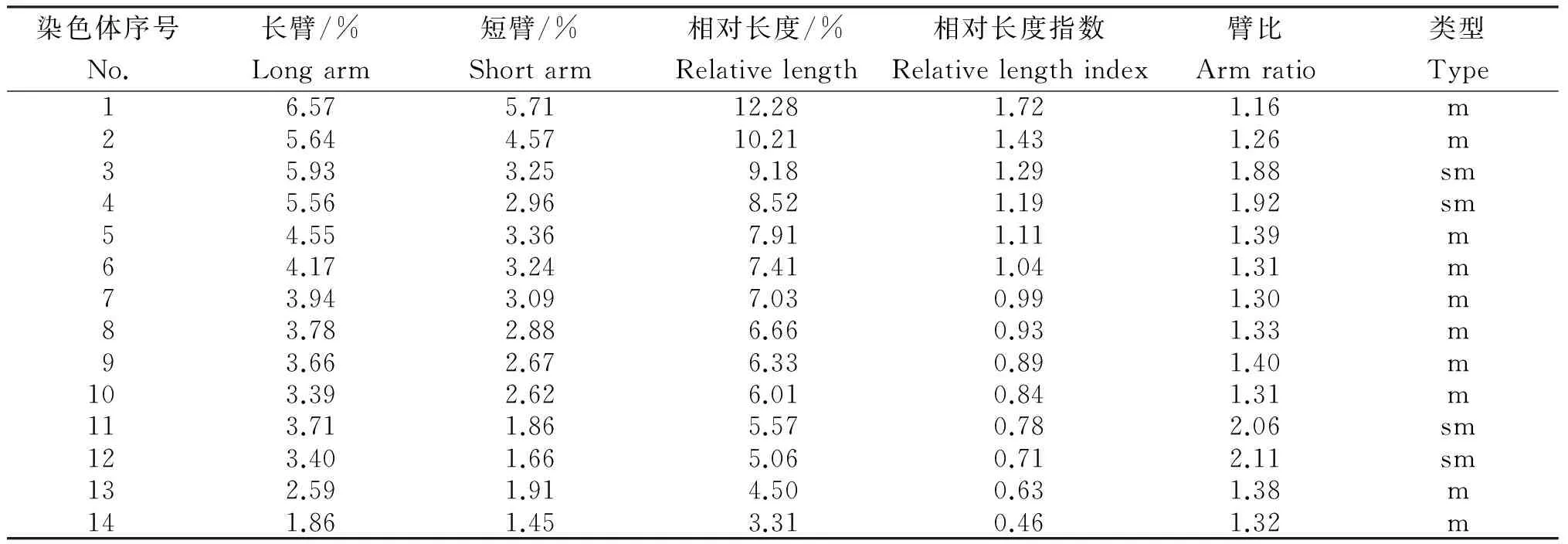
表1 象草的染色体核型参数

表2 象草与巨菌草染色体的核型特征
2.3 巨菌草的细胞染色体
由图2可见,30个巨菌草染色体分散较好的细胞染色体数目为28条和26条分别有28个和2个细胞,分别占计数总数的93.3%和7.7%,按照核型分类标准[23],最终确定巨菌草染色体数目为2n=4x=28。
2.4 巨菌草的核型
由表2和表3可知,巨菌草染色体的数目为2 n=4 x=28,其中第3号、第4号、第11号、第12号为近中部着丝粒染色体(sm),第13号、第14号为近端部着丝粒染色体(st),其余均为中部着丝粒染色体(m)。核型公式为2 n=4 x=28=16 m+8 sm+4 st。核染色体相对长度在4.13%~11.78%,其组成为2 n=4 x=28=4 L+8 M2+10 M1+6 S,最长和最短染色体比为2.85,染色体臂比大于2∶1的染色体占6.67%。按照核型分类标准,巨菌草的核型属2A型,核型不对称系数为59.86%。
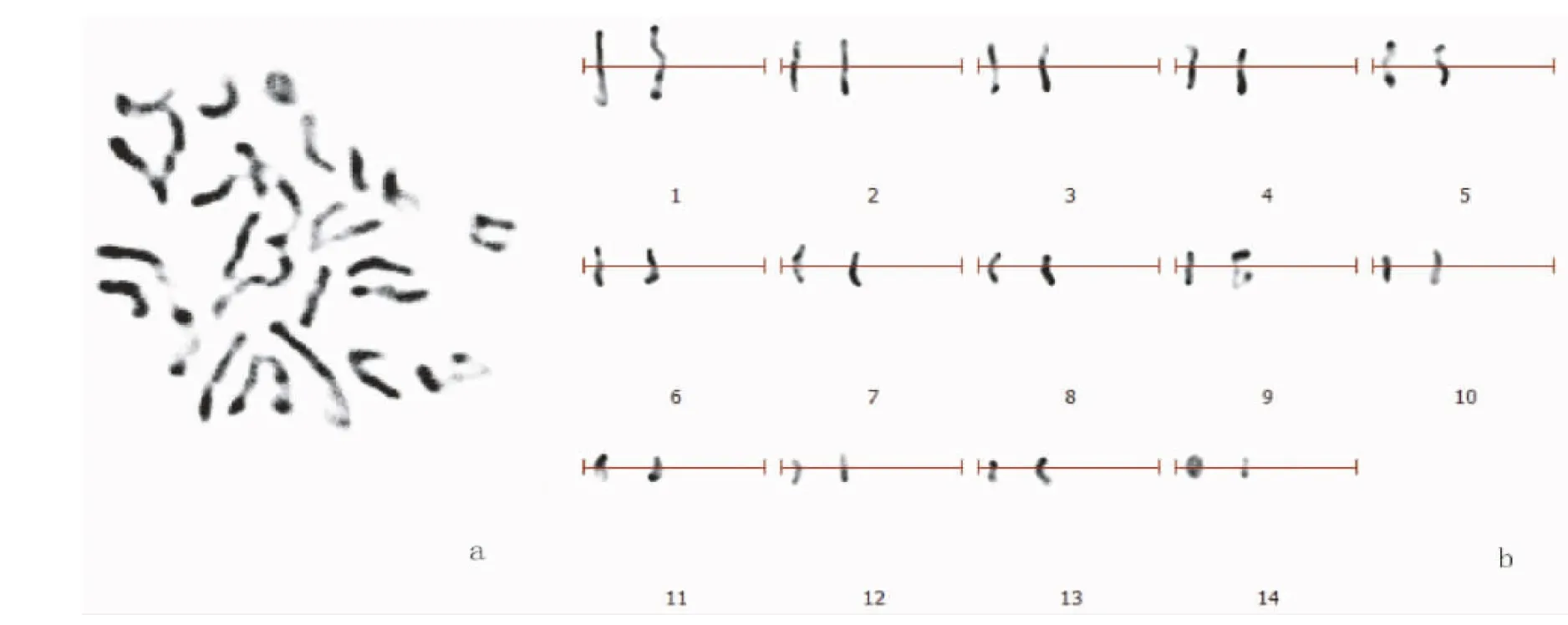
注:a为巨菌草染色体中期相,b为巨菌草核型图。
Note: a, metaphase phase of chromosomes; b, chart of chromosome karyotype.
图2 巨菌草染色体中期相和核型图
Fig.2 Metaphase chromosomes and karyotype pairs ofP.giganteum
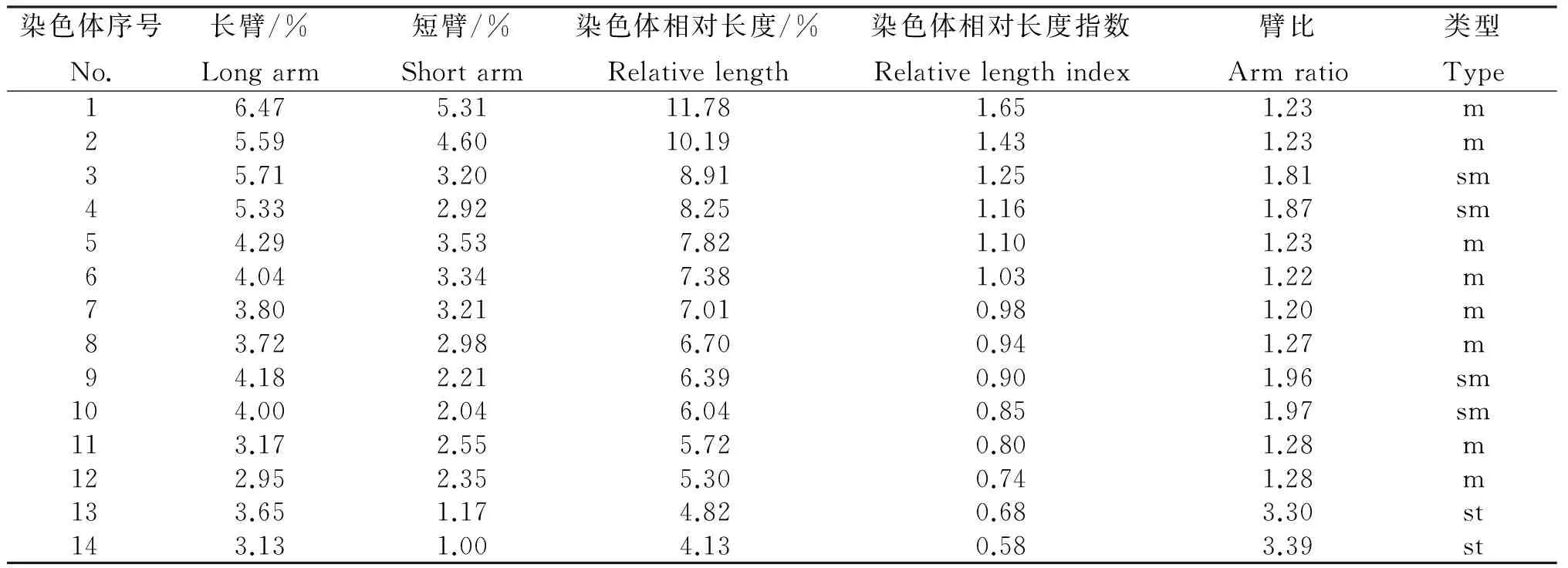
表3 巨菌草的染色体核型参数
3 结论与讨论
每一个物种染色体的结构、数目和形态不同。象草和巨菌草的体细胞染色体总数目均为2 n=4 x=28,无随体现象出现。其中象草的核型公式为2 n=4 x=28=20 m+8 sm,相对长度为3.31%~12.28%,组成为2 n=4 x=28=6 L+6 M2+10 M1+6 S,属于2A型,核型不对称系数为58.77%;巨菌草核型公式为2 n=4 x=28=16 m+8 sm+4 st,相对长度为4.13%~11.78%,组成为2 n=4 x=28=4 L+8 M2+10 M1+6 S,也属于2A型,不对称系数为59.86%。两者的核型不对称系数为58%~60%,具有较高的对称性,表明在进化上属于比较原始的类型。普遍认为,在植物界中细胞核型进化的基本趋势由对称向不对称发展[19],巨菌草的核型不对称系数略大于象草,说明巨菌草的进化程度比象草高,此结论国内鲜有报道。
一般来说,核型数据如染色体的数目、倍性、相对长度、随体的数量及位置等参数均是鉴定种和亚种的重要理论依据[20-25]。周自玮等[12]研究云南德宏地区逸生狼尾草的体细胞染色体为2 n=4 x=28,与本试验结果一致,由此推断云南地区的逸生狼尾草与巨菌草、象草亲缘关系较近。阎贵兴等[26]在禾本科7属33种饲用植物的染色体研究结果中表明其基数均为7,且均由m和sm染色体组成〔东方早麦草(Eremopyrum orientale (Linn.) Jaub. et Spach)例外〕,多数种类核型对称程度较高,与本试验结果大体一致。有研究表明,狼尾草属的狼尾草,其染色体有2n=18、14、36,而象草染色体只有2n=28、56[27],可推断巨菌草与象草是不同的象草品种类型。另据巨菌草的 rDNA ITS 序列测定结果发现,巨菌草与牧草蔗、莱牧1号的同源性达92.1%,推测巨菌草是杂交狼尾草的一个品种[28]。结合核型分析结果以及分子方面的研究,可推断巨菌草隶属狼尾草属,且与象草的亲缘关系较近。但与前人结果不同的是,此材料没有观察到随体,这可能与试验材料和染液有关。染色体上的着丝粒由于为人工设定,存在一定误差,故需要进一步将染色体C带分析、原位杂交等技术运用于染色体研究,以期获得更准确的细胞学信息。同时对巨菌草的研究应结合细胞学、形态学和分子标记等方面的研究方法,为鉴定此品种提供更加详尽和准确的生物学信息,以达到巨菌草的育种、推广以及种质资源保护的目的。
[1] 林占熺.菌草学[M].3版.北京:国家行政学院出版社,2013.
[2] 崔红标,杨家妮,周 静,等.磷灰石和石灰联合巨菌草对重金属污染土壤的改良修复[J].农业环境科学学报,2013,32(7):1334-1340.
[3] 肖 正,赵 超,刘 斌.巨菌草高温发酵产沼气的初步研究[J].安徽农学通报,2012(17):18-21.
[4] 张 辉.造纸新原料皇竹草[J].造纸信息,2005(5):28.
[5] 董晓娜,陈喜蓉,钟剑锋,等.巨菌草栽培灵芝试验初探[J].热带林业,2013,41(1):39-40.
[6] 林兴生,林占熺,林冬梅,等.5种菌草苗期抗盐性的评价[J].福建农林大学学报,2013,43(2):195-201.
[7] 林兴生,林占熺,林 辉,等.五种菌草苗期对碱胁迫的生理响应及抗碱性评价[J].植物生理学报,2013,49(2):167-174.
[8] 林兴生,林占熺,林冬梅,等.低温胁迫5种菌草的抗寒性评价[J].草业学报,2013,22(2):227-234.
[9] 黄水珍.8种狼尾草属牧草农艺性状和鲜草产量分析及相关性研究[J].养殖与饲料,2011(11):39-41.
[10] 陈志彤,何水林,黄毅斌.狼尾草属牧草研究进展[J].草地学报,2010(5):740-748.
[11] Il R R,Yoon J H,Hyung I L.Karyotype analysis using FISH(fluorescence in situ hybridization) in Fragaria[J].Scientia Horticulturae,2012,136:95-100.
[12] 周自玮,袁福锦,匡崇义,等.云南德宏地区逸生狼尾草的综合鉴定[J].中国草地学报,2008(3):110-114.
[13] 张 伦,康 念.长序狼尾草的核型分析[J].贵州科学,1992,10(2):44-47.
[14] 李懋学,张敩方.植物染色体研究技术[M].哈尔滨:东北林业大学出版社,1991(3):31-48.
[15] 李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985(4):297-302.
[16] Levan A,Fredga K,Sandberg A A.Nomenclature for centromeric position on chromosomes [J].Hereditas,1964,52:197-201.
[17] Kuo S R.Karyotype analysis of some Formosan gymnosperms[J].Taiwania,1972,17(1):66-80.
[18] Arano H.The karyotype and the speciations in subfamily Carduoideae of Japan[J].Japanese Journal of Botany,1965,19(3):31-67.
[19] Stebbins G L.Chromosomal Evolution in Higher Plan ts [M].London:Edward Arnold L TD,1971:87-89.
[20] 吴姝菊.2种寒地稗草农艺学性状与细胞学研究[J].草地学报,2010,18(2):280-285.
[21] 王志宁,季道藩,许复华.棉属四个栽培种的核型比较研究[J].浙江农业学报,1997(3):135-139.
[22] 李桂芬,梁国鲁,林顺权,等.枇杷属植物核型分析及其在系统分类中的应用[J].园艺学报,2013,40(8):1465-1474.
[23] 辛培尧,尚勋武,郭鸿彦,等.大麻染色体行为分析[J].西北植物学报,2008,28(11):2189-2193.
[24] 张义贤.胡枝子的核型研究[J].山西大学学报,1990,13(1):87-89.
[25] 李国泰,刘 斐.东方蓼染色体的核型分析[J].通化师范学院学报,2009(2):48-49.
[26] 阎贵兴,张素贞,云锦凤,等.33种禾本科饲用植物的染色体核型研究[J].中国草地,1991(5):1-13.
[27] 陈默君,贾槙修.中国饲用植物[M].北京:中国农业出版社,2002:236-247.
[28] 卞承荫,黄舒婷.五个菌草新品种rDNA ITS序列克隆与遗传多样性分析[EB/OL].北京:中国科技论文在线[2014-06-10].http://www.paper.edu.cn/releasepaper/content/201406-145.
(责任编辑: 刘忠丽)
Karyotype Analysis ofPennisetumpurpureumandPennisetumgiganteum
ZHU Dandan1, WANG Peidan1, LIN Xingsheng2, LIN Hui2, SU Dewei2, LIN Zhanxi2*
(1.CollegeofLifeScience,FujianAgricultureandForestryUniversity,Fuzhou,Fujian350002; 2.ChinaNationalEngineeringResearchCenterofJuncaoTechnology,Fuzhou,Fujian350002,China)
To provide certain cellular basis for germplasm, breeding and genetic kinship ofP.giganteum, The karyotype analysis and chromosome number ofP.purpureumandP.giganteumwere conducted by traditional squash method. The results showed that the karyotype morphology of materials were similar, consisted of metacentric chromosomes and sub-metacentric chromosomes mostly. In this study, the karyotype formulas ofP.purpureumandP.giganteumwere respectively 2 n=4 x=28=20 m+8 sm, 2 n=4 x=28=16 m+8 sm+4 st. Constitutions of relative length of two plants were 2 n=4 x=28=6 L+6 M2+10 M1+6 S, 2 n=4 L+8 M2+10 M1+6S. The karyotype ofP.purpureumwas the same as that ofP.giganteumin chromosome basic number and total number. And two varieties were classed as 2A. The asymmetry coefficient were in the range of 58% to 60%, which demonstrated basic symmetry karyotypes. Conclusion: Both of the two plants had no significant differences in chromosomes between them and the relationship between them was close from cellular level.
Pennisetumpurpureum;Pennisetumgiganteum; chromosome; karyotype
2015-01-26; 2015-04-29修回
福建省科技重大专项“水土流失初步治理区生态循环与产业提升技术研发与示范”(2012NZ0002);科技部项目“国家菌草工程技术研究中心组建”(2011FU125X12)
朱丹丹(1989-),女,在读硕士,研究方向:植物遗传学研究。E-mail:1042527543@qq.com
*通讯作者:林占熺(1943-),男,研究员,博士生导师,联合国国际生态安全科学院院士,从事菌草研究。E-mail:linjuncao@163.com
1001-3601(2015)05-0226-0014-05
Q949
A
