鄂西利川地区二叠-三叠系界线附近地层的碳酸盐微相类型和早三叠世火山活动的证据
2015-02-25姚华舟赵小明杨振强
余 聪,姚华舟,赵小明,杨振强
YU Cong1,2,3,YAO Hua-Zhou2,ZHAO Xiao-Ming2,YANG Zhen-Qiang2
(1.中国地质科学院,北京100037;2.中国地质调查局武汉地质调查中心,武汉430205;3.中国地质调查局古生物与生命—环境协调演化重点实验室,武汉430205)
(1.Chinese Academy of Geological Sciences,Beijing 100037,China; 2.Wuhan Center of China Geological Survey,Wuhan 430205,Hubei,China; 3.Key Laboratory of Paleontology and coevolution of life,CGS,Wuhan 430205,Hubei,China)
鄂西利川地区二叠-三叠系界线附近地层的碳酸盐微相类型和早三叠世火山活动的证据
余 聪1,2,3,姚华舟2,赵小明2,杨振强2
YU Cong1,2,3,YAO Hua-Zhou2,ZHAO Xiao-Ming2,YANG Zhen-Qiang2
(1.中国地质科学院,北京100037;2.中国地质调查局武汉地质调查中心,武汉430205;3.中国地质调查局古生物与生命—环境协调演化重点实验室,武汉430205)
(1.Chinese Academy of Geological Sciences,Beijing 100037,China; 2.Wuhan Center of China Geological Survey,Wuhan 430205,Hubei,China; 3.Key Laboratory of Paleontology and coevolution of life,CGS,Wuhan 430205,Hubei,China)
本文对湖北利川地区两个剖面的二叠-三叠系界线层序进行了沉积微相分析,划分出6种微相类型,讨论了每种微相的成因和沉积环境,揭示了微相环境的演变规律。并且对早三叠世的火山活动进行了讨论,以此探讨P/T之交生物礁和微生物岩的古环境意义以及生物大灭绝的诱因。
碳酸盐岩微相;微生物岩;P/T界线;生物大灭绝
1 引言
二叠纪末期曾发生过地质历史上影响最大的第三次生物绝灭事件,导致97%的海洋生物与70%的陆地生物在这次事件中永远消失[1-3]。在此之后的早三叠世古气候、古生物和古环境发生了巨大的变化,生态系长期处于大萧条状态。用来解释灭绝事件的理论有许多,但是在灭绝的模式和诱因方面至今仍存在较大争议。
二叠纪生物大灭绝时间之后,微生物重新开始大量繁盛[4-5]。Kershaw等[6,7]首次在川东华蓥山地区长兴期生物礁之上识别出微生物岩。Ezaki等[8]在其中发现了微生物活动的重要证据,并获得重要的生物地层学资料,确定了微生物岩的发育时代。Lehrmann等人[9-11]对扬子地台南部地区二叠-三叠系界线、早三叠世微生物岩、浅水碳酸盐岩向上变浅沉积旋回以及稳定碳同位素变化进行了深入的研究。Adachi等[12]对早三叠世微生物岩中的似球粒(peilods)进行了描述和成因讨论。微生物岩富集事件与生物演化时间以及环境变迁之间有着密切的关系。现有研究表明,微生物岩主要产出于礁顶或极浅水碳酸盐岩台地之上,向深水区很快尖灭[13],而且已发现的微生物岩中的主要生物化石均与非微生物岩相区的生物有明显的差别。因此,微生物岩中所保存的化石具有明显的地理区限,也就是说具有很好的原地性。这为重建大灭绝后礁相或极浅水碳酸盐岩台地相的生态系提供了难得的研究载体。
鄂西地区二叠-三叠纪界线地层发育完整且出露好,是研究二叠-三叠纪界线地层的理想地区,沉积类型丰富[14],在沉积相环境方面也取得了一定的研究进展[14-17],但仍有部分典型剖面研究程度较低。
本文选择鄂西利川瑞坪、夏家槽两剖面为主要研究对象,主要目的是在沉积微相分析的基础上,详细研究该区二叠-三叠纪之交的沉积环境变迁、海平面变化和火山事件,以丰富二叠纪末生物大灭绝的环境背景及二叠-三叠纪之交海平面变化的研究。
2 古地理背景及地质概况
2.1 沉积特征及古地理背景
P/T之交钙质微生物岩在全世界广泛分布,主要存在于低纬度浅水海洋环境中,具有较好的等时性[18]。华南板块位于古特提斯洋东部的赤道附近,是微生物岩最主要的分布地区。中扬子区晚二叠世主要发育碳酸盐岩开阔台地、陆棚及盆地沉积,吴家坪期以浅海相沉积为主,深水盆地范围较小,长兴期发育台地相及盆地相,深水盆地范围变大[19]。
三叠纪早期构造运动加剧,古地理环境变化大,沉积相类型多变,深水碳酸盐沉积和浅水碳酸盐交替出现。三叠系底部普遍见有代表火山(或地外撞击)事件的凝灰质泥岩。台地边缘出现许多碳酸盐重力流、风暴岩和远源浊积岩。台地上广泛发育了生物建隆、藻礁、树枝状微生物岩,以及潮间带的藻鲕粒、藻层纹石、叠层石、隐藻团粒和隐藻凝块的微生物岩沉积。海水盐度增高,局部出现石膏假晶。海水表面温度上升。
2.2 剖面简介
利川市位于鄂西边陲,四川盆地以东,地理坐标为:N 30°00'-30°20',E 108°30'-109°00'[20]。二叠-三叠系剖面发育齐全,代表了浅水台地相区。利川著名的见天坝生物礁剖面为碳酸盐岩台地边缘相,而本研究的瑞坪、夏家槽两剖面位于该生物礁之南的碳酸盐岩台地相区(图1)。
瑞坪剖面(图2)位于马鞍山向斜北东端,交通便利,地层沿新开挖的小公路展布,出露良好,层序清楚,剖面产状平缓。下部为长兴组的含燧石结核灰岩、泥灰岩;其上为钙质微生物岩,以树枝状微生物岩为主,中间夹有灰岩以及灰质泥岩;上部为中层状泥岩与灰黑色叶片状片岩所构成的韵律层。
夏家槽剖面(图3)位于北北东向齐岳山背斜北西翼,有乡间小公路相连,交通便利,地层沿小公路出露良好,层序清楚。下部为长兴组块状生物碎屑灰岩;灰色钙质斑状微生物岩整合于其上,可分为叠层石状微生物岩、花斑状微生物岩、树枝状微生物岩三种类型,总厚度约3.11m;微生物岩上部为大冶组深灰色中厚层状生物屑泥粉晶灰岩,以及硅化灰岩夹薄层泥岩;其上为粘土岩。
3 材料和方法

图1 中国南方晚二叠世-早三叠世古地理及典型剖面分布点[21]Fig.1 Late Permian-Early Triassic palaeogeography of South China and typical profile points location

图2 瑞坪剖面柱状图Fig.2 Ruiping section column
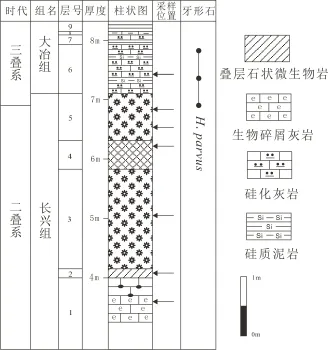
图3 夏家槽剖面柱状图Fig.3 Xiajiacao section column
碳酸盐岩沉积微相分析是碳酸盐岩沉积学研究的重要方法和手段,对于了解地史时期的沉积环境模式和岩相古地理具有重大意义。二叠纪-三叠纪之交生物大绝灭、残存和复苏已成为研究热点之一,众多学者对此提出了种种解释,但随着研究的深入,人们逐渐认识到只有在详细客观论证地层沉积特征的基础之上,生物大灭绝和海平面变化的关系以及生物灭绝的机制才能迎刃而解。
在瑞坪和夏家槽P/T界线附近地层共取得20个微相分析样品。在镜下主要观察薄片中的生物类型、磨蚀程度、岩石组分、颗粒粒度等,结合野外工作收集的岩石宏观和微观特征标志,参照标准微相类型[22]及其沉积环境模式,对剖面碳酸盐岩进行微相分析,总结出微相类型。碳酸盐岩微相类型是沉积环境的反映,与海平面变化密切相关,所以微相类型是研究海平面变化的一个重要参数。
4 二叠-三叠纪之交的微相类型
本研究的两个地层剖面位于灰岩型沉积相区[14]内。通过对采集薄片的镜下鉴定,结合已有的样品资料以及野外工作收集的岩石宏观和微观特征标志,参照威尔逊[22]标准微相类型及其沉积环境模式,对碳酸盐岩微相进行了初步研究,归纳总结出了6种沉积微相类型。
4.1 有孔虫微晶灰岩【MF-1】
产于长兴组,瑞坪剖面的第1层,宏观上呈深灰色厚层含燧石结核灰岩,镜下为微晶灰岩,主要生物为有孔虫和薄壳的介形虫壳体,有极少量的双壳类,生物个体非常小(图版Ⅰ-a),说明当时的水体可能相对较深。相当于深水陆棚相带中的生物碎屑泥晶灰岩至泥晶生物碎屑灰岩[23]。
另外,在刘家场型的深水陆棚相区,还可见到含放射虫黑色页岩(根据姚华舟采集标本的薄片样品)。该泥质页岩呈黑色,有机质丰富,半透明,其中含有10%~15%放射虫(图版Ⅰ-b)。放射虫为硅质成分,直径为0.08~0.24 mm左右。见于利川见天坝海绵生物礁剖面以北的硫黄厂和硫厂坪剖面一带,属于炭质灰岩-泥岩型沉积相区[14]。这是一种典型的局限性的缺氧的深水陆棚-盆地或盆地边缘相类型,与碳酸盐台地交替分布,相当于威尔逊(1975)[22]标准微相类型SMF-1。
4.2 藻-有孔虫生物碎屑灰岩【MF-2】
出现于夏家槽剖面第1层。该类微相生物碎屑含量主要介于30%~60%之间,以蜓类、非蜓有孔虫、介形虫、钙藻类为主,双壳类等生物碎屑较少。(图版Ⅰ-c)
该微相类型基质主要是以微晶为主,含少量的亮晶,可能反映了一种相对较弱动荡的浅水环境;蜓类属浅海底栖生物,生活于水深20~100 m的热带或亚热带,多数化石颗粒完整,但有部分显示有弱泥晶化现象,表明少数蜓在水底短暂停留或徘徊,而大多数为原地埋藏;有孔虫、藻类等化石颗粒保存较好,棘皮类等颗粒破碎可能说明它们是由高能区被搬运到低能的泻湖环境中来;较多粗枝藻的出现反映该灰岩形成于潮汐面之下的温暖浅水环境。据研究,在这种微相类型中,结壳状的异常丰富,它为二叠纪常见的生物,常常被认为是藻类,并成集合体形式构成生物丘,或者在生物建隆中起着粘结作用。但是一些研究者认为是一种钙质海绵幼体集合体[23]。可见,微相类型MF-2反映一种台地边缘后的海峡或者海湾环境的沉积环境,即为一种开阔海台地相,相当于威尔逊(1975)[22]相带7及8的标准微相类型SMF-18有孔虫藻粒灰岩。
4.3 叠层石状微生物岩【MF-3】
该微相类型产于夏家槽剖面第1层顶部至第2层底,野外岩性呈灰黑色中厚层块状灰岩,在宏观上表现出包菜状的藻叠层石。藻叠层石呈高5 cm和宽6 cm左右的集合体(图版Ⅱ-a、b),单圈层厚度约为2~3 mm,分布较密,间距约1~3 cm。微观上为微晶-细晶的碳酸盐矿物,偶呈带状相间排列分布。基质中还含有少量藻类,几乎不含其他生物碎屑。野外采集标本表面可见密集的微球状构造(图版Ⅱ-c),微观上表现为藻包壳和藻包粒,大的藻包粒个体呈圆形,直径约为0.6 mm以上,小的圆形藻凝块石直径为0.2~0.3 mm,核心为重结晶的镁方解石或白云石所充填,并被藻泥晶包壳所包围(图版Ⅰ-d)。藻包粒的外圈之间为微亮晶方解石胶结的基质。
4.4 花斑状钙质微生物岩【MF-4】
该微相类型产于瑞坪剖面第4层以及夏家槽剖面第3、5层。宏观上表现为花斑状的构造(图版Ⅱ-d、e),镜下显示为细晶-微晶组成的藻斑点、斑块、凝块石,形状不规则。花斑状的构造是一种最为普遍的沉积构造,在华南地区分布很广。具有花斑状构造的微生物岩通常被称为凝块石[18]。
微生物岩通常被认为形成于受限制的局限或半局限性潮下低能的沉积环境中。在树枝状或花斑状钙质的微生物岩中,常常保存有较完整的介形虫、腹足类等化石。亦可反映开放-受限的潮下生物建隆的沉积环境[14]。吴亚生等[24]指出花斑状构造微生物岩为局限台地相沉积。现代巴哈马和鲨鱼湾地区微生物岩研究表明:凝块石多出现在水体较动荡、沉积速率较快且相对开阔的浅潮下带环境,并且生物扰动对凝块石结构的形成有较大影响[25-26]。
4.5 树枝状钙质微生物岩【MF-5】
该微相类型产于瑞坪剖面第6、8层,夏家槽剖面第4层。宏观上呈灰色至灰黑色中-厚层块状。该微相类型在宏观上表现出树枝状构造(图版Ⅱ-e),此种构造主要是由微生物岩中的微晶和中粗晶组分呈近垂直层面的树枝状排列引起的。而微观上表现出由较均一的微细晶基质和方解石颗粒组成的特征。生物面貌以低丰度、低分异度为特征,仅含极少量有孔虫、介形虫、腹足类、腕足类等(图版Ⅰ-e)。这是一种类似生物建隆环境中直立的树枝状或似叶状体的生物捕捉或藻粘结作用造礁环境,可称为生物障积岩,相当于威尔逊(1975)[22]相带5的标准微相类型SMF-7生物障积岩。
4.6 含黄铁矿的泥晶灰岩【MF-6】
产于夏家槽剖面第5层顶。野外露头呈薄层夹中厚层灰岩。生物量极低,含少量或极少量黄铁矿。大部分为泥晶,有些已重结晶或白云石化,白云石化并成团块状堆积。极少有微裂缝或微方解石脉,偶尔见发育有一些缝合线。反映出由于沉积速率较慢,沉积作用受到影响,岩层发生凝缩减薄,为潮下较深水的陆棚环境(图版Ⅰ-f)。
姚华舟等将晚二叠世长兴期时长江三峡地区浅水碳酸盐台地与深水盆地结合带的沉积相分为五种沉积类型[14]。根据姚华舟提供的采集标本的薄片样品,通过镜下观察,认为除了上述描述的微相类型以外,至少可识别以下3种碳酸盐岩微相类型:
4.7 薄壳微生物泥晶灰岩【MF-7】
泥晶灰岩中以薄壳的介形虫,有孔虫为特征(图版Ⅰ-g),表明水介质环境不利于厚壳钙质生物的发育,水盐度加大或水温升高引起对生物的抑制。这是一种典型的盐度不正常的受限制的浅海-陆棚碳酸盐台地的潮下沉积。
4.8 含有孔虫细晶灰岩和白云岩【MF-8】
细晶灰岩和白云岩中见有密集的细小几丁质盘状、棒状、管状和短柱状有孔虫和微生物,生物内部结构发生简单化的变异(图版Ⅰ-h)。这是一种典型的高盐度介质条件下的受限制的局限或半局限性碳酸盐台地沉积。
4.9 砾屑和火山晶屑泥岩【MF-9】
泥岩中见有火山成因的砾状泥岩屑(火山砾屑或火山弹?)(图版Ⅱ-g),直径可达0.2~0.3 cm,还见有大量粘土化的玻屑和火山晶屑[14]。晶屑的粒径为0.15~0.6 mm,呈尖棱角状、钩状、港湾状和碎裂状(图版Ⅱ-h)。这是一种典型的盆地边缘相类型。
二叠-三叠系界线层的微相类型是受限制的浅海-陆棚碳酸盐台地的潮下沉积,和缺氧的深水陆棚-盆地硅质沉积,以及与火山作用有关的砾屑、火山晶屑凝灰质泥岩,代表了局限台地环境恶化,海水咸化、缺氧以及受火山作用的控制强烈。可见三峡地区的过渡层的相类型向局限或半局限性环境发展,推测可能是由于火山作用加剧的原因,导致水介质盐度增高,生物个体变小,钙壳质变薄(变为几丁质)。
5 早三叠世早期火山活动的证据
华南二叠-三叠系界线附近普遍发育有多层薄层的粘土岩层。它们的主要特征是:成层薄,一般厚度不超过10 cm,分布广泛,有明显的等时性,与上下岩层接触面平整,含海相化石。界线粘土岩层是重要的研究对象,因为从这些界线附近的粘土岩层中,可能找到地质历史时期曾经发生过的火山喷发或者外星撞击等事件所遗留下来的蛛丝马迹,从而找到导致二叠、三叠系之交生物大绝灭的真正原因[27]。
前人对二叠-三叠系界线地层进行了大量研究,发现石英晶屑在二叠-三叠系界线地层中十分常见,并且主要存在于粘土岩中。这些粘土岩层大多具有成层薄、分布广、具有等时性;粘土矿物成分以伊/蒙混层粘土矿物为主;碎屑物质中具有晶形完整的、未受磨圆的六方双锥石英、锆石、磷灰石、火山灰、微球粒等与火山活动有关的物质,这些特征明显地反映了粘土岩的形成与火山活动密切相关[27]。
鄂西地区二叠-三叠系界线附近地层一般发育有3-5层粘土岩,最多发育7层,主要表现为灰黄色、黄棕色、浅黄色,或者带点蓝色的黄白色伊利石、蒙脱石或者伊蒙混层粘土岩。而这些特征大体上与华南其它地区类似[28]。在利川瑞坪剖面和夏家槽剖面界线地层处各发育有两层火山凝灰岩,其中瑞坪剖面火山凝灰岩层厚度分别为11 cm和12 cm;夏家槽剖面火山凝灰岩层厚度分别为17 cm和19 cm。说明在三叠纪早期利川地区发生过火山事件。
在三峡地区界线层的薄片样品中发现了大量石英晶屑(图版Ⅱ-h,图版Ⅲ),剖面中可见厚约4 cm的伊利石粘土岩,也指示了该地区在早三叠时期的火山活动。
6 结论
本文以鄂西利川的二叠-三叠系地层为研究对象,通过实地观察和测量、采集样品和室内显微镜观察,分析了类型,还原了二三叠之交环境的变化。结合邻区的相关资料,所得结论如下:
在利川地区瑞坪、夏家槽两个剖面共划分出六种微相类型,包括有孔虫微晶灰岩、藻-有孔虫生物碎屑灰岩、叠层石状钙质微生物岩、花斑状钙质微生物岩、树枝状钙质微生物岩、含黄铁矿的泥晶灰岩。三峡地区的过渡层的微相类型具有环境恶化的特征。
瑞坪、夏家槽剖面界线层之上均发育有火山凝灰岩,指示了本区早三叠世可能发生火山事件,另外在三峡地区剖面的薄片中发现有大量石英晶屑,也是该区早三叠世火山活动的有力证明。
[1]Erwin D H.The Great Paleozoic Crisis [M].New York: Columbia University Press,1993:1-327.
[2]Erwin D H,Bowring S A,Jin Y.End-Permian mass extinctions:A review[M]//In:Koeberl C and MacLeod K G(eds). Catastrophic Events and Mass Extinctions:Impacts and Beyond.Geological Society of America Special Paper,2002, 356:363-383.
[3]Alroy J.Dynamics of origination and extinction in the marine fossil record[J].Proceedings of the National Academy of Sciences,2008,105:11536-11542.
[4]Baud A,Richoz S,Pruss S.The lower Triassic anachronistic carbonate facies in space and time [J].Global&Planetary Change,2007,55(1-3):81-89.
[5]Kershaw S,Li Y,Crasquin-Soleau S,Feng Q L,Mu X A, Collin P Y,Reynolds A,Guo L.Earliest Triassic microbialites in the South China block and other areas:controls on their growth and distribution[J].Facies,2007,53(3):409-425.
[6]Kershaw S,Zhang T S,Lan G Z.A?Microbialite Carbonate Crust At The Permian-Triassic Boundary In South China, And Its Palaeoenvironmental Significance[J].Palaeog eography Palaeoclimatology Palaeoecology,1999,146(1):1-18.
[7]Kershaw S,Guo L,Swift A,Fan J S.?Microbialites in the Permian-Triassic boundary interval in central China:age and distribution[J].Facies,2002,47(1):83-90.
[8]Ezaki Y,Liu J,Adachi N.Earliest Triassic Microbialite Micro-to Megastructures in the Huaying Area of Sichuan Province,South China:Implications for the Nature of Oceanic Conditions after the End-Permian Extinction [J].Palaios, 2003,18(4):388-402.
[9]Lehrmann D J.Early Triassic calcimicrobial mounds and biostromes of the Nanpanjiang basin,south China[J].Geology, 1999,27:359-362.
[10]Lehrmann D J,Wan Y,Wei J Y,Yu Y Y,Xiao J F.Lower Triassic peritidal cyclic limestone: an example of anachronistic carbonate facies from the Great Bank of Guizhou,Nanpanjiang Basin,Guizhou province,South China[J].Palaeogeography Palaeoclimatology Palaeoecology, 2001,173(3-4):103-123.
[11]Lehrmann D J,Payne J L,Felix S V,Dillett P M,Wang H M, Yu Y Y,Wei J Y.Permian-Triassic boundary sections from shallow-marine carbonate platforms of the Nanpanjiang basin,South China:Implications for oceanic conditions associated with the end-Permian extinction and its aftermath.Palaios,2003,18(2):138-152.
[12]Adachi N,Ezaki Y,Liu J.2004.The fabrics and origins of peroids immediately after the end-Permian extinction, Guizhou Province,South China[J].Sedimentary Geology,16 (1):161-178.
[13]王永标,童金南,王家生,周修高.华南二叠纪末大绝灭后的钙质微生物岩及古环境意义[J].科学通报,2005,50 (6):552-558.
[14]姚华舟,张仁杰.长江三峡地区晚二叠世晚期-早三叠世早期沉积特征[J].华南地质与矿产,1996,(4):63-68.
[15]杨振强,林甲兴.从鄂西二叠纪碳酸盐岩微相探讨有孔虫的生态环境[J].沉积学报,1983,(4).
[16]王延奇,胡明毅,刘富艳,王辉,胡治华.鄂西利川见天坝长兴组海绵礁岩石类型及礁体演化阶段[J].岩性油气藏,2008,20(3):44-48.
[17]赵小明,童金南.浙江煤山钻孔二叠-三叠系界线剖面遗迹化石的两幕式变化[J].中国科学:地球科学,2010,40 (9):1241-1249.
[18]何磊,王永标,杨浩,廖卫,翁泽婷.华南二叠纪-三叠纪之交微生物岩的古地理背景及沉积微相特征[J].古地理 學報,2010,12:151-163.
[19]万秋,李双应,孔为伦,王松.中扬子晚二叠世沉积特征及古地理演化[J].地质科学,2011,46(2):336-349.
[20]王令占,涂兵,赵小明,田洋,谢国刚,吴俊,童金南,安志辉,曾波夫,李珉.湖北利川地区1∶5万区域地质调查成果与主要进展 [J].华南地质与矿产,2013,29(3): 169-176.
[21]刘丽静,姜红霞,吴亚生,蔡春芳.中国南方晚二叠世-早三叠世礁区生物群落演替序列与古环境变化——以四川盆地东北部盘龙洞剖面为例[J].中国科学:地球科学, 2014,44(4):617-633.
[22]威尔逊J L.冯增昭等译.地质历史中的碳酸盐相 [M].北京:地质出版社,1981:1-365.
[23]杨振强,林甲兴.从鄂西二叠纪碳酸盐岩微相探讨有孔虫的生态环境[J].沉积学报,1983,1(4):92-106.
[24]吴亚生,Yang W,姜红霞,范嘉松.江西修水二叠纪-三叠纪界线地层海平面下降的岩石学证据[J].岩石学报,2006,22(12):3039-3046.
[25]Turner E C,James N P,Narbonne G M.Taphonomic Control on Microstructure in Early Neoproterozoic Reefal Stromatolites and Thrombolites[J].Palaios,2009,15(2):87-111.
[26]Planavsky N,Ginsburg R N.Taphonomy of Modern Marine Bahamian Microbialites[J].Palaios,2009,24:5-17.
[26]殷鸿福,黄思骥.华南二叠纪-三叠纪之交的火山活动及其对生物绝灭的影响[J].地质学报,1989,(2):169-180.
[27]张素新,冯庆来,曾毅.广西柳桥深水相二叠系-三叠系界线附近粘土岩特征 [J].电子显微学报,2003,(6): 637-638.
[28]Yao H Z,Zhang R J.Sedimentary Facies Types and Volcanics near the Permo-Triassic Boundary in the Yangtze Gorge Area[J].Acta Geoscientica Sinica,1996:199-204.
Yu C,Yao H Z,Zhao X M and Yang Z Q.Carbonate microfacies in strata near the Permian-Triassic boundary and the volcanic activity evidence in the early Triassic in Lichuan area,Western Hubei Province.2015,31(2)∶115-124.
∶Carbonate microfacies sequence analysis is carried out in two sections of the Permian-Triassic boundary in Lichuan area,Hubei province,which can be divided six microfacies types,and genesis and sedimentary environment of each kind microfacies are discussed to reveal the revolution of micro environment. The volcanic activities in the Early Triassic in this area are also discussed.The combined evidence is used to investigate the palaeoenvironment significance of the reef and microbialite,and the cause of the biomass extinction in the Permian-Triassic boundary.
∶carbonate microfacies;microbialite;Permian-Triassic boundary;biomass extinction

图版Ⅰ

图版Ⅱ

图版Ⅲ
P53
A
1007-3701(2015)02-115-10
10.3969/j.issn.1007-3701.2015.02.001
2015-04-15;
2015-05-31.
余聪(1990—),女,硕士在读,古生物与地层学专业,E-mail:shanhucong12@163.com.
