祁连圆柏群落物种多样性的海拔梯度效应研究
2015-02-22占玉芳滕玉风田晓萍鲁延芳
占玉芳,马 力,滕玉风,田晓萍,鲁延芳
(张掖市林业科学研究院,甘肃 张掖 734000)
祁连圆柏群落物种多样性的海拔梯度效应研究
占玉芳,马 力,滕玉风,田晓萍,鲁延芳
(张掖市林业科学研究院,甘肃 张掖 734000)
祁连圆柏;物种多样性;海拔梯度效应;祁连山
为研究珍稀树种祁连圆柏群落物种多样性的垂直变化效应,在对祁连山北坡中段2 800~3 200 m间祁连圆柏群落进行调查的基础上,分析了不同海拔梯度祁连圆柏群落物种的重要值和物种多样性。结果表明,不同海拔梯度下的祁连圆柏群落在结构组成上有所不同,物种的重要值也有差异;不同生活型植物群落物种多样性对海拔梯度变化的敏感程度不同,草本层植物最敏感,其次是灌木群落,物种多样性与海拔梯度之间呈单峰关系。祁连圆柏群落物种多样性的垂直分布规律在植物的不同层次存在差异,草本层物种多样性较高且海拔梯度效应明显,即在海拔3 200 m的群落中,物种多样性和丰富度达到最高;灌木层多样性和均匀度在海拔2 800 m地带最高,但变化趋势不明显。在同一海拔梯度范围内,草本层的物种多样性和丰富度均明显高于灌木层。总的来说,祁连圆柏群落物种多样性海拔梯度变化规律明显,物种多样性高,种类组成复杂,群落乔木层、灌木层、草本层各层层次明显。
祁连圆柏(Sabinaprzewalskii)为常绿乔木,一般高达12 m,原产于青海、甘肃河西走廊及南部、四川北部,是中国特有的常绿针叶树种,以它为建群种所形成的天然林广泛分布于中国西北部海拔2 600~4 000 m的亚高山及高山地区[1],为祁连山林区亚高山暗针叶林带中唯一分布在阳坡或半阳坡上的森林类型[2],具较强的抗旱能力。其作为祁连山重要的水源涵养林之一,在水源涵养、调节径流、水土保持、净化水质、调洪蓄水与保持生态平衡等方面有重要的作用。
物种多样性反映了生物群落和生态系统的结构和功能复杂性,揭示了物种组织水平的生态学基础[3]。植物物种多样性随环境梯度的变化规律一直是生态学家十分关注的问题[4]。物种的空间分布格局受许多生态梯度的影响,而海拔梯度包含了土壤因子、气候因子、植被群落间的相互影响等多种环境因子,被认为是影响物种多样性格局的决定性因素之一[5]。因此,海拔梯度上的植物物种多样性格局成为植物生态学家的重要研究内容。祁连山北坡具有较大的海拔梯度,突出表现为海拔梯度上水热因子的剧烈变化,如年均温度从6 ℃(海拔1 500 m) 变为-5.4 ℃(海拔3 800 m),年降水量从129.7 mm(海拔1 500 m)变为690.26 mm(海拔3 800 m)[6],因此祁连山也是山地植被垂直多样性分布格局研究的典型山体之一。但有关祁连山北坡植物物种多样性垂直梯度方面的研究报道较少。本研究拟通过沿不同海拔梯度设置样地,研究祁连圆柏群落物种多样性的垂直变化效应,以丰富祁连圆柏群落生态学的内容,并为祁连圆柏群落生物多样性保护及其资源的利用、祁连圆柏濒危机制的探讨提供理论依据。
1 研究区概况
研究区位于甘肃省张掖市肃南县寺大隆林区的天涝池流域,地理坐标为99°31′~100°15′E、38°14′~38°44′N,海拔2 500~3 600 m。该区年平均气温0.7 ℃,1月平均气温-13.3 ℃,7月平均气温11.8 ℃;年降水量440.58 mm,其中5—9月降雨量占全年降水总量的89.2%,年蒸发量1 061.8 mm,年均相对湿度60%,属温带高寒半干旱气候。土壤以山地森林灰褐土为主。土壤和植被随山地地形和气候的差异而形成明显的垂直分布带。受生长环境条件的限制,乔木层树种结构单一,其他伴生树种很少。灌木层常见的植物有鬼箭锦鸡儿(Caraganajubata)、短叶锦鸡儿(Caraganabrevifolia)、金露梅(Potentillafruticosa)、高山绣线菊(Spiraeaaipina)、刚毛忍冬(Lonicerahispida)等,植被覆盖度约为40%。草本层以禾本科植物为主,主要有短颖鹅观草(Roegneriabreviglumis)、垂穗披碱草(Elymusnutans)、红棕薹草(Carexdigyne)、披针薹草(Carexlanceolata)、矮生嵩草(Kobresiahumilis)、北方嵩草(Kobresiabellardii)、绢茸火绒草(Leontopodiumsmithianum)、宽苞棘豆(Oxytropislatibracteata)、高山唐松草(Thalictrumalpinum)等。
2 研究方法
2.1 调查方法
2012年7月,选择在祁连山寺大隆林区祁连圆柏分布的集中区域进行野外调查。采用样地法对祁连圆柏群落进行调查,分别在海拔2 800、3 000、3 200 m处各建立20 m×20 m的调查样地3个,每个样地取4 m×4 m灌木样方4个,并在每个灌木样方中取1 m×1 m草本样方1个。记录指标有:乔木和灌木的物种名称、株数、高度、胸径、地径、冠幅、乔木层盖度、灌木层盖度,草本的名称、株数、高度等。同时,记录样地的群落总盖度、地理位置、经纬度、坡度、坡向、海拔、土壤类型,以及人类活动的干扰情况。采用R.P.McIntosh和J.T.Curtis等提出的计算重要值的方法[7]来计算草本层、灌木层物种重要值。
2.2 物种多样性测定方法
采用丰富度指数、Shannon-Wiener指数、Simpson指数、Pielou均匀度指数、种间相遇几率5个多样性指标[8]进行测定。
2.2.1 丰富度指数
物种丰富度指数R用群落中物种的总数S进行计算,公式为
式中:N为样方中观察到的所有物种的个体总数;S为群落中物种的总数。
2.2.2 物种多样性
物种多样性采用目前应用最为广泛的Shannon-Wiener指数H和Simpson优势度指数D表示,公式为
式中:N、S意义同上;ni为第i个物种的个体数;Pi为第i个物种的个体数占总个体数N的比例,Pi=ni/N。
2.2.3 物种均匀度
均匀度是指样地中各个物种多度的均匀程度,即每个物种个体数间的差异。基于Shannon-Wiener指数的Pielou均匀度指数J的计算公式为
式中:S意义同上;H为Shannon-Wiener指数。
2.2.4 种间相遇几率
种间相遇几率PIE,或称群落组织水平相关关系指数,计算公式为
式中:N、i、S意义同上。
3 结果与分析
物种多样性是指一定区域中物种各类的丰富程度和各物种分布的均匀程度。由于群落的物种组成受制于环境因子的影响,因而也对群落的物种多样性有着很大的影响[9]。祁连圆柏是祁连山林区亚高山暗针叶林带中唯一分布在阳坡或半阳坡上的森林类型[2],因所处的环境条件较差,故呈稀疏分布,森林覆盖率较低,且一般生长发育不良,低矮扭曲。但是它作为分布区内干旱地区森林群落阳坡的主要建群种,是祁连山重要的水源涵养林。特殊的环境条件使祁连圆柏群落形成了自己独特的物种多样性,其结构较为简单,乔木层只有祁连圆柏1种,盖度小,无层间植物,所以只对灌木层和草本层的重要值、物种多样性进行计算。
3.1 不同海拔梯度祁连圆柏群落物种重要值分析
重要值是反映植物种类在群落中地位重要程度的综合指标之一[10],其大小也是确定建群种和优势种的重要依据,故对不同海拔梯度祁连圆柏群落不同层次物种重要值进行分析具有重要意义。根据不同海拔梯度祁连圆柏群落物种调查结果,可以把祁连圆柏群落划分为3个层次,即乔木层、灌木层、草本层。乔木层只有祁连圆柏1种,重要值为1,灌木层和草本层主要物种的重要值见表1(草本层只列出了重要值大于0.2的物种)。
由表1可知,不同海拔梯度下的祁连圆柏群落在结构组成上有所不同,物种的重要值也有差异。灌木层以金露梅、银露梅为主:海拔2 800 m群落中,以银露梅为主,其重要值达到了0.927 7;海拔3 000 m群落中,金露梅、银露梅的重要值相差不大,金露梅为0.439 4、银露梅为0.560 6;海拔3 200 m群落中,金露梅、银露梅的重要值相差不大,金露梅为0.462 4、银露梅为0.537 6。由此可以看出,海拔3 000~3 200 m的群落中,灌木层重要值随海拔梯度变化效应不明显,这是由于灌木层树种对不同海拔的适应能力相差不大造成的。在草本层中,海拔3 200 m群落优势种重要值最大的是圆穗蓼(0.489 0),海拔3 000 m群落优势种重要值最大的是矮生嵩草(0.844 1),海拔2 800 m群落优势种重要值最大的是多茎委陵菜(0.353 9)。由此可知,草本层重要值的海拔梯度效应较为明显,如:多茎委陵菜在海拔2 800 m的重要值为0.353 9,在海拔3 000 m的重要值为0.223 9,在海拔3 200 m的重要值为0.250 6;同时矮生嵩草在海拔3 000 m占有绝对优势,圆穗蓼在海拔3 200 m占有绝对优势。这说明,草本层同一物种的重要值在不同海拔梯度也不同,即不同海拔梯度的环境条件造成了群落优势种的不同。

表1 不同海拔梯度灌木层、草本层主要物种的重要值
3.2 海拔梯度与物种多样性分析
3.2.1 不同海拔梯度物种丰富度
不同海拔梯度的祁连圆柏群落的个体数、物种数、优势种等都有所不同(表2),其中个体数、物种数是最直观的多样性指标。在不同海拔的祁连圆柏群落中,海拔3 000 m群落的草本层物种数量最多,海拔3 200 m群落次之,海拔2 800 m群落最少。在不同海拔梯度群落中,乔木层、灌木层个体数量和物种数变化不明显。海拔3 000 m群落草本层个体数最多,灌木层、乔木层个体数最少;海拔2 800 m群落草本层个体数次之,灌木层个体数34个,乔木层个体数最多;海拔3 200 m群落草本层个体数最少,灌木层个体数最多,乔木层个体数41个。由此可知,金露梅、银露梅对此生态环境适应且地位稳定,为不同海拔梯度祁连圆柏群落灌木层中的常见种。

表2 祁连圆柏群落不同海拔梯度物种及个体数
3.2.2 灌木层物种多样性
由表2可知,祁连圆柏群落灌木层物种较少,每个海拔梯度只有2种。由表3可看出,随着海拔的增加,灌木层丰富度指数R、种间机遇率PIE呈先增大后减小的趋势,多样性指数H、均匀度指数J呈减小的趋势,优势度D呈增大趋势。在灌木层物种多样性海拔梯度效应中,海拔2 800 m群落丰富度指数R、多样性指数H、均匀度指J均大于海拔3 200 m群落,这说明在较低海拔2 800 m的群落中,灌木层物种能较好地适应生长环境,个体分布均匀且生长良好;随着海拔升高,适应生长的灌木层物种会减少,物种优势度明显。
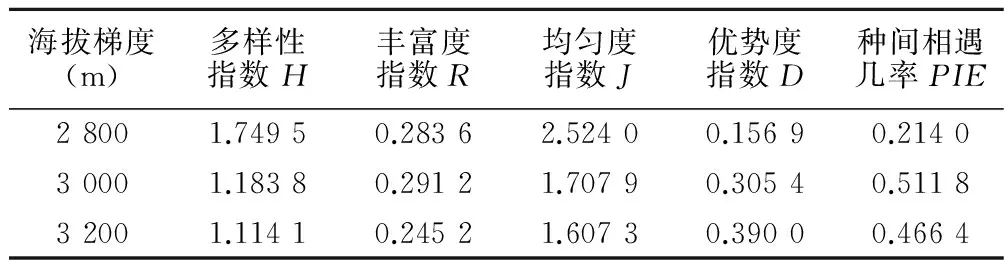
表3 灌木层物种多样性指数
祁连圆柏群落中灌木层物种多样性偏低,主要有两个方面的原因。一方面是由于祁连圆柏群落分布在阳坡,阳坡接受的辐射强度大,水分蒸发快,通常比较干燥,容易造成植物生理干旱,影响灌木的生长,致使其他灌木物种很难入侵阳坡;且祁连圆柏是一种寒温带针叶树种,群落所分布的海拔较高,其灌木层的物种多样性低于温带和亚热带森林群落符合生物多样性的分布规律[9]。另一方面也与樵采、放牧等人为活动有关,因为亚高山林草交错区阳坡灌丛草甸往往是良好的冬牧场[11],牛羊的过度啃食与践踏抑制了灌木的更新与生长。
3.2.3 草本层物种多样性
草本层物种多样性指数见表4。在草本层中,海拔3 000 m群落的物种数最多,但海拔3 200 m的丰富度指数R、多样性指数H、种间机遇率PIE都最大,优势度指数D和均匀度指数J略低于海拔2 800 m。这说明在不同海拔梯度中,海拔3 200 m是向高山草甸过渡地带,地形较开阔,为草本植物生长提供了良好的生长环境,因而草本植物个体分布均匀,多样性较高。这与贺金生等[12]总结出的群落物种多样性随海拔梯度变化的5种模式之一是吻合的。植物群落物种多样性在中等海拔高度较低,如落基山脉国家公园Colorado Front Range山脉的东坡[13],属干旱大陆性气候,森林郁闭度较低,在中等海拔及山坡中部的森林中物种多样性较低,而在沟谷中的森林、向高山草甸过渡地带、地形较开阔的地方,物种多样性较高。

表4 草本层物种多样性指数
祁连圆柏群落草本层的多样性程度高,与乔木层和灌木层的稀疏有关,上层乔灌木稀疏使阳光可以直接照射到草本层,从而促进草本植物的生长。充足的阳光除了促进草本种子萌发外,还能促进植物体本身的生长,使其顺利完成正常的发育周期。海拔3 200、2 800 m的样地草本层均匀度指数大于0.7,这说明祁连圆柏群落草本层各物种都能比较好地适应林草交错区阳坡的特殊环境。
4 讨 论
(1)用重要值来研究祁连圆柏群落不同物种优势度,用多样性指数来测定其群落在不同海拔梯度下的物种多样性,通过比较分析可以说明该群落的结构、组织水平,以及演替与生境等方面关系的生态学特征,具有重要的生态学价值及实践意义。
(2)从本研究结果来看,祁连山北坡寺大隆林区珍稀树种祁连圆柏群落物种多样性的垂直分布规律在植物不同层次存在差异。草本层物种多样性较高且海拔梯度效应明显,即在海拔3 200 m的群落中,物种多样性和丰富度达到最高;灌木层多样性和均匀度在海拔2 800 m地带最高,但变化趋势不明显。说明在同一区域植物群落的物种多样性对海拔梯度的响应也存在差异,位于温带干旱区的祁连圆柏群落物种多样性在中等海拔处较低。同时,在同一海拔梯度范围内,草本层的物种多样性和丰富度均明显高于灌木层,这与温带森林群落不同层次物种多样性的测度结果一致[14]。这也说明了祁连圆柏群落不同海拔梯度不同层次物种多样性格局是对区域气候特征的直接响应。
(3)祁连圆柏群落物种多样性海拔梯度效应研究结果表明:不同生活型植物群落物种多样性对海拔梯度变化的敏感程度不同,草本层植物最敏感,其次是灌木群落,且物种多样性与海拔梯度之间呈单峰关系。就资源的可利用性而言,物种多样性的海拔梯度效应反映了多样性和资源生产力之间的关系。因此,研究祁连圆柏群落物种多样性海拔梯度效应,对保护祁连圆柏及其群落物种多样性具有一定的指导意义,可为保护好该地区的珍稀树种、发展林业生产、改善人类生存的自然环境服务。
[1] 文陇英,陈拓,张满效,等.不同生境下祁连圆柏叶片色素和稳定碳同位素组成的变化[J].冰川冻土,2010,32(4):823-828.
[2] 张涛,安黎哲,陈拓,等.不同海拔青海云杉与祁连圆柏叶片抗氧化系统[J].植物生态学报,2009,33(4):802-811.
[3] 赵洋毅,王玉杰,王云琦,等.渝北不同模式水源涵养林植物多样性及其与土壤特征的关系[J].生态环境学报,2009,18(6):2260-2266.
[4] 冯建孟,王襄平,徐东成,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110-116.
[5] 李俞萍.昆仑山北坡中段植被生态及营养特征的垂直梯度研究[D].北京:北京师范大学,2010.
[6] 王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多样性,2002,10(1):7-14.
[7] 杨梅娇.甜槠林群落结构及物种多样性特征研究[J].福建林业科技,2006,33(4):36-40.
[8] 赵志模,郭依泉.群落生态学原理与方法[M].重庆:科学技术文献出版社重庆分社,1990.
[9] 陈文年,吴宁,罗鹏,等.岷江上游林草交错带祁连山圆柏群落的物种多样性及乔木种群的分布格局[J].应用与环境生物学报,2003,9(3):221-225.
[10] 丁圣彦.浙江天童常绿阔叶林演替系列栲树和木荷成为优势种的原因[J].河南大学学报:自然科学版,2001,31(1):79-83.
[11] 吴宁,刘照光.青藏高原东部亚高山森林草甸植被地理格局的成因探讨[J].应用与环境生物学报,1998,4(3):290-297.
[12] 贺金生,陈伟烈.陆地植物群落物种多样性的梯度变化特征[J].生态学报,1997,17(1):91-99.
[13] Peet R K.Forest vegetation of the Colorado Front Range:Patterns of species diversity[J].Vegetatio,1978,37(2):65-78.
[14] 黄建辉,高贤明,马克平,等.地带性森林群落物种多样性的比较研究[J].生态学报,1997,17(6):611-618.
(责任编辑 徐素霞)
中央预算内林业基本建设投资计划项目(林规发〔2010〕240号)
S791.44;Q948.114
A
1000-0941(2015)08-0052-04
占玉芳(1970—),女,甘肃民勤县人,副研究员,学士,主要从事林业科研、林木良种繁育及推广工作;通信作者马力(1964—),男,甘肃武威市人,研究员,硕士,主要从事森林生态及林业科研、林木良种繁育及推广工作。
2014-09-20
