乌兰巴托—锡林浩特样带草地植被特征与水热因子的关系
2015-02-10胡云锋巴图娜存毕力格吉夫刘纪远
胡云锋, 巴图娜存, 毕力格吉夫, 刘纪远, 甄 霖
1 中国科学院地理科学与资源研究所, 北京 100101 2 中国人民大学,环境学院, 北京 100872 3 内蒙古自治区草原勘察规划院, 呼和浩特 010051
乌兰巴托—锡林浩特样带草地植被特征与水热因子的关系
胡云锋1,*, 巴图娜存1,2, 毕力格吉夫3, 刘纪远1, 甄 霖1
1 中国科学院地理科学与资源研究所, 北京 100101 2 中国人民大学,环境学院, 北京 100872 3 内蒙古自治区草原勘察规划院, 呼和浩特 010051
2012年夏季,研究人员对蒙古高原长约1100km的乌兰巴托—锡林浩特草地样带开展考察,获取了46个样地的物种数量、地上生物量等数据;基于全球GHCN(全球历史气象网络)数据集,提取了样带夏季(6—8月)月均温度和降水总量;继而根据自然地理和行政区边界,将草地样带大致分成北部(蒙古国乌兰巴托—蒙古国艾日格)、中部(蒙古国艾日格—中国苏尼特左旗)和南部(中国苏尼特左旗—中国锡林浩特),开展了分析。研究表明:(1)样带夏季平均温度的空间分布形态呈现明显的倒“U” 型分布,南北两端温度较低,中部温度较高;夏季降水量在空间上的分布形态则与之相反,呈现南北两端降水量较高,中部降水量较低的正“U”型分布;(2)样带上植物物种数量、地上生物量的空间分布形态均呈现正“U” 型分布,即在生态景观类型为典型温性草原的样带南部和北部地区,其生物多样性、地上生物量明显好于呈现为温性荒漠草原、温性荒漠景观的样带中部地区。(3)相关分析体现了大尺度(高原样带尺度)上植被特征与水热环境因子间的关系:植物物种数量、地上生物量与夏季月均温度均呈现负相关,而与夏季降水总量则呈现正相关关系。(4)偏相关分析反映了局地小尺度上植被特征与水热环境因子间的关系:温度和降水要素对于植物物种数量、地上生物量均呈现正相关。
蒙古高原; 草地样带; 植被特征; 环境因子; 相关分析
草原植物的种属、数量及生物量等指标是衡量草原生物多样性、生态系统生产力和承载能力的重要参数,这些指标与环境要素间的关系是生态学家长期关注的问题。在全球气候变化背景下,上述草原植被特征对于环境变化的区域响应,更是研究的热点问题之一[1]。蒙古高原是欧亚大陆上相对独立的地理生态单元,总体上为典型温性草原生态系统,也是我国北方重要的生态屏障。就蒙古高原草原植被特征及其与环境要素关系开展研究,对于探索蒙古高原区域生态系统分布格局及演化态势具有重要理论意义,对于评估蒙古高原草地生态系统生态屏障作用具有重要实践价值[2]。然而,由于地域广大、国界分割、语言文字差异、野外工作环境艰苦等诸多原因,当前国内外研究人员对蒙古高原资源环境的研究并不十分透彻。
一方面,大多数研究主要集中在中国内蒙古高原地区,对于蒙古国境内的研究相对较少。大部分研究主要集中在大气环流形势、历史气候变化、沙尘天气及一些特定植物物种区系分布、植物生理微形态学、种属分支系统演化等[3- 7],此外还有一些关于高原植被生长和健康状况分布格局、长时间序列植被参数对气候变化响应、局地土壤风蚀速率测量、区域风蚀危险度评估等研究[8- 10]。总的来看,植物学家的分析常缺乏对蒙古高原总体分异规律的刻画,而地理学家的大尺度分析则在第一手数据支持方面略显不足。
其次,在目前大多数草原植被特征的研究中,由于生态系统的层次性和复杂性,特别是由于尺度推移效应,研究人员在不同地区、不同尺度上关于草原植物物种数量、生物量与环境因子间关系的认识并不统一[11- 15]。例如:白永飞的研究表明水、热因子是影响内蒙古锡林河流域草原群落植物多样性和生产力及主要因素[16- 17];胡云锋、艳燕等的研究也表明:内蒙古东北—西南草地样带上,生物多样性随着水热变化表现出明显的梯度变化规律[18]。蔡学彩等根据内蒙古锡林河南岸大针茅草原样地上的多年观测数据分析,认为生物量与降水量的变化不存在显著相关性[19];马文红等依据内蒙古高原113个样地数据分析,认为降水是导致内蒙古温带草地生物量空间变异的主要因子[20];陈效逑等针对典型草原样地开展研究,认为地上生物量对干燥度变化的响应非常敏感[21]。很多深入的研究还表明,环境因子中的降水因子在时间上的分布(季节分配、频次、强度、间隔等)对于草地生产力有着重要影响[22- 28]。因此,在蒙古高原尺度上,草原植被特征与温度、降水、干燥度等是否存在相关性?这有待于研究人员开展系统的采样设计和分析研究。
针对上述科学问题,本文研究人员于2012年夏对蒙古高原乌兰巴托—锡林浩特一线开展了野外综合科学考察。考察采用样带调查方式,自西北向东南贯穿了整个蒙古高原。考察采用等间距采样,对样带各点的土地利用与土地覆被、草地植物群落及农牧民生计等问题开展了综合调研。本文即利用样带草地样方调查成果,主要采用相关分析方法,对草地样带植被特征与水热等环境因子的关系进行定量分析和建模。
1 研究区、数据与方法
1.1 研究区
蒙古高原以祁连山、贺兰山和阴山山脉为南界,以唐努山、肯特山为北界,以阿尔泰山为西界,以大兴安岭山脉为东界;广义的蒙古高原还包括阴山、黄河以南的鄂尔多斯高原。从行政区划上看,蒙古高原大致包括蒙古国全部、中国内蒙古自治区、甘肃省北部、新疆自治区东部及俄罗斯东西伯利亚南部。由于四面远离海洋,周围为高、中山所环绕,蒙古高原呈现为山地与高原为主体的地貌格局,是亚欧大陆上一个相对封闭的内陆生态地理单元。
蒙古高原平均海拔1580 m,地势整体自西向东、自南向北逐渐降低;除东部、南部少数地区外,其它地区年降雨量一般少于400 mm,是典型的大陆性干旱、半干旱区。本区土地荒漠化严重,沙尘天气频发,是影响我国华北乃至整个东亚的主要沙尘源区之一[29- 30];高原主要植被类型为草原,具体可分为草甸草原、典型草原、荒漠草原和戈壁等几个次级类型,并以典型草原分布面积最大。
1.2 样带设置
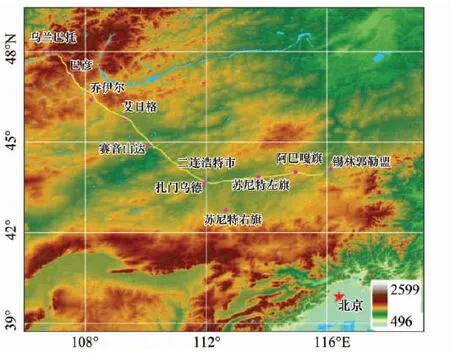
图1 研究区及乌兰巴托—锡林浩特草地样带示意图
2012年8月20日至30日,研究组对蒙古高原乌兰巴托、锡林浩特一线开展了科学考察。考察组成员主要来自中国科学院地理科学与资源研究所、内蒙古草原勘察设计研究院、蒙古国科学院植物研究所、中国人民大学、内蒙古自治区锡林郭勒盟草原监测站等单位。
野外考察起点为蒙古国乌兰巴托市(Ulaanbaatar),终点为中国内蒙古自治区锡林浩特市。在蒙古国境内,考察路线是沿着乌兰巴托—二连浩特—北京国际铁路沿线展开,在中国境内则是沿省道S101向东南方向展开。考察路线穿越了蒙古国中央省(Tov)、中戈壁省(Dundgovi)、戈壁苏木贝尔省(Govi-Sumber)、东戈壁省(Dornogovi)以及中国内蒙古自治区锡林郭勒盟,经过的主要城镇包括:乌兰巴托(Ulaanbaatar)、巴彦(Bayan)、乔伊尔(Choir)、艾日格(Airag)、赛音山达(Sainshand)、扎门乌德(Zanym-Uud),二连浩特、苏尼特右旗、苏尼特左旗、阿巴嘎旗和锡林浩特。路线全长约1100 km,海拔高程最低为887 m(蒙古国南戈壁省奥尔贡),最高为1650 m(蒙古国中央省巴彦)。
鉴于上述考察路线反映了蒙古高原气温、降水以及其它生态地理要素梯度变化的典型方向,因此该考察路线被定名为“蒙古高原乌兰巴托—锡林浩特草地样带”(图1)。进一步,根据自然地理和行政区分界,大致可以将上述草地样带分成北部(蒙古国乌兰巴托—蒙古国艾日格)、中部(蒙古国艾日格—中国苏尼特左旗)和南部(中国苏尼特左旗—中国锡林浩特)。
1.3 数据获取和预处理
针对蒙古高原地形坦荡、生态地理单元尺度较大、生态景观单一且连续过渡特点,考察采用等距离采样方法,即沿着考察路线、每隔20—25 km开展一次踏勘采样。植物样方调查基本流程是:首先使用GPS测定经纬度和高程,使用数码相机拍摄四至景观及正下方草地样方照片;而后在每个样地做3个植物详细样方,记录草地类型、草群高度、物种名称、高度、盖度、生物量等信息;最后在上述3个样方周围,随机选取10个物种频度样方。经统计,共有46个样地、近500个样方,采集了约136份草地生物量样品、2800余张相片。
气象数据来源于美国NOAA网站所提供的GHCN(The Global Historical Climatology Network,全球历史气象网络)数据集(http://www.ncdc.noaa.gov/ghcnm/)。温度和降水数据的时间序列均为1950—2000年,其中降水数据来自全球47554个站点,温度数据来自全球24542个站点;对上述观测数据开展空间插值,最终形成了空间分辨率为1km、时间分辨率为1月的空间数据集。基于上述GHCN数据集,在ArcGIS环境中提取46个样地所在格点的数据,得到各个样地1950—2000年期间夏季(JJA:6—8月)逐月的温度、降水数据。
1.4 分析方法
草地样带生物多样性、生物量与水热因子的关系主要通过事实描述和统计分析(简单相关、偏相关和多元回归)等方法进行研究。事实描述主要是基于气象观测数据、野外调查结果,通过简单计算和制图,在草地样带延伸方向上对各个样地的物种数量、生物量以及气候因子进行数据平滑、曲线拟合,说明其空间变化趋势。统计分析是本文重点,研究中具体使用了简单相关分析、偏相关分析以及多元回归分析等3种统计方法。这些统计分析方法中所涉及的指标定义本文不再赘述,具体的统计分析工作是在SPSS(V17)的支持下完成的。
2 结果分析
2.1 气候因子变化
夏季月均温度在样带呈倒置“U” 型分布,而夏季降水总量在样带呈正的“U” 型分布,夏季平均温度和降水总量在空间上的分布态势正好相反(图2)。

图2 夏季平均温度、降水总量的空间变化
样带北部,即蒙古国乌兰巴托和蒙古国中央省地区,夏季平均温度最低,为(16.3±1.5) ℃,样地间的变异系数为9%;样带中部,即蒙古国东戈壁省赛音山达地区附近,夏季平均温度最高,为(20.7±0.8) ℃,变异系数为4%;样带南部,也即中国锡林郭勒盟地区,夏季平均温度较低,为(19.0±0.6) ℃,变异系数为3%。
与夏季平均温度空间分布态势变化正好相反,样带上夏季降水的空间分布则呈正的“U”型。即:样带北端夏季降水总量较高,为(166.8±27.0) mm,变异系数16%,样带中部夏季降水最低,仅为(94.9±12.0) mm,变异系数12.7%,样带南部夏季降水总量最高,为(167.7±17.0) mm,变异系数10%。
2.2 物种数量变化
样带上草原植物物种数在空间上大致呈现为南北高、中部低的“U”形分布(图3)。
样带北部,即蒙古国乌兰巴托和蒙古国中央省地区,平均每个样地内有植物(21±4)种,样地间的变异系数为18%;样带中部,即蒙古国东戈壁省赛音山达地区附近,平均每个样地中出现的植物(13±2)种,其变异数为11%;样带南部,即中国锡林郭勒盟地区,平均每个样地内有植物(20±3)种,其变异系数为14%。
上述事实表明,主要为典型温性草原的样带南部、样带北部,其物种数显著地高于呈现为温性荒漠草原、温性荒漠景观的样带中部地区,样带北部和南部的生态系统更具生物多样性。但是,从物种数量的局地空间差异(即样地间的变异系数)上看,样带中部的荒漠草原和荒漠区的物种数量维持情况要比样带南北两端的温性典型草原区更稳定。
2.3 地上生物量变化
样带草地地上生物量在空间上的部分明显呈现为南北高、中部低的“U”形(图4)。
在样带北部,即蒙古国乌兰巴托和蒙古国中央省地区,生物量较高,平均为(85±29) g/m2,样地间的变异系数为34%;样带中部,即蒙古国东戈壁省赛音山达地区,生物量较低,平均为(62±31) g/m2,变异系数为50%;样带南部,即中国锡林郭勒盟地区,其生物量最高,平均约为(208±69) g/m2,变异系数为33%。
上述事实表明,主要为典型温性草原的样带南部、样带北部,其地上生物量明显高于呈现为温性荒漠草原、温性荒漠景观的样带中部地区,样带南部和北部生态系统具有更高的地上部分生产力。并且,从地上生物量的局地空间差异(即样地间的变异系数)上看,样带南部和北部地区的草原地上生物量稳定性要较样带中部更稳定;并且在同一区域上,生物量的空间变异程度明显要比物种数量的空间变异程度要大得多。
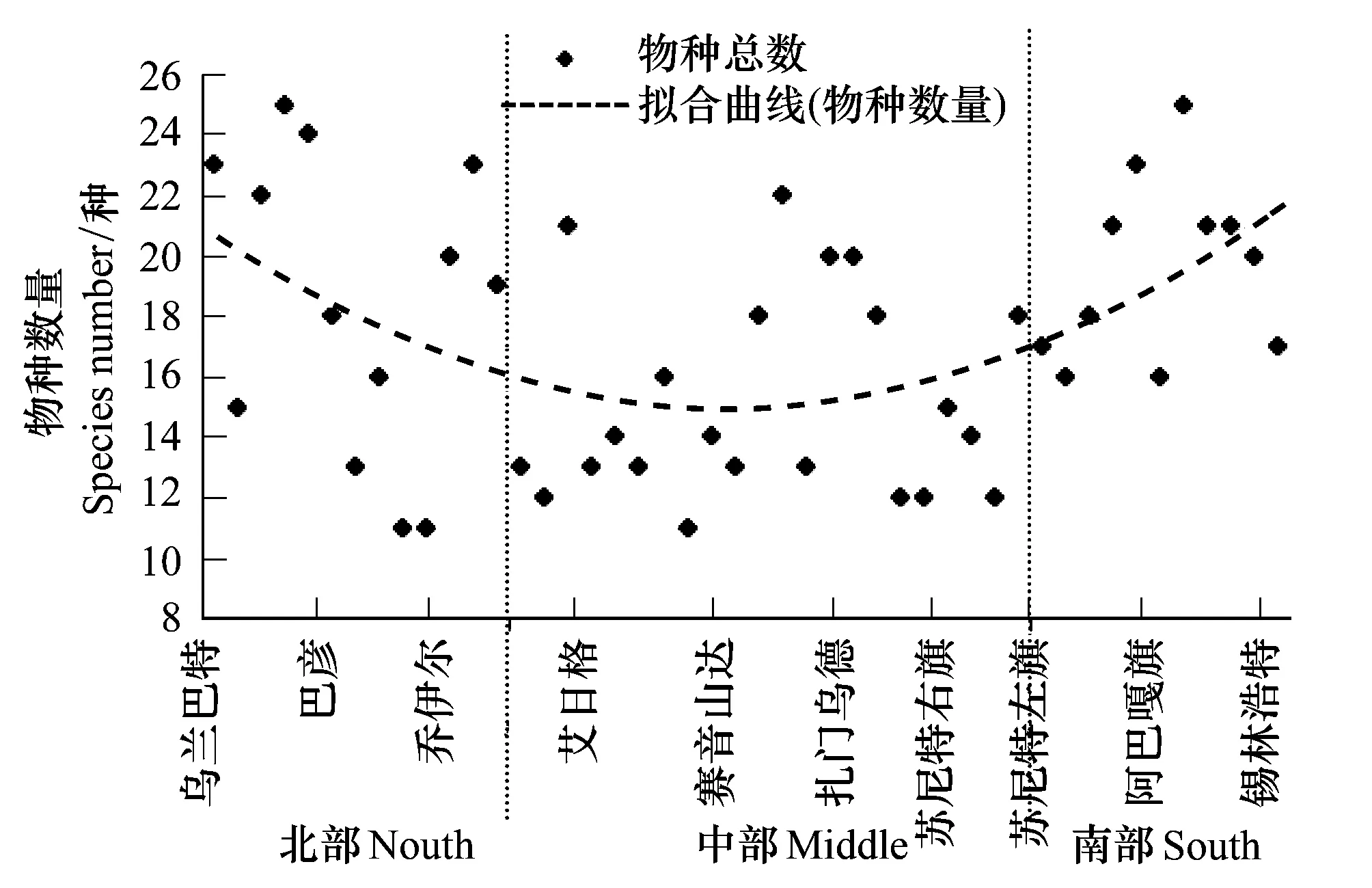
图3 样带物种数量的空间变化
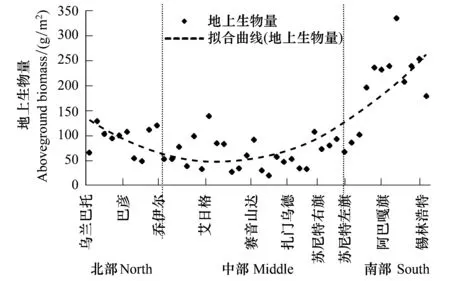
图4 样带地上生物量的空间变化
2.4 地上生物量与物种数量的关系
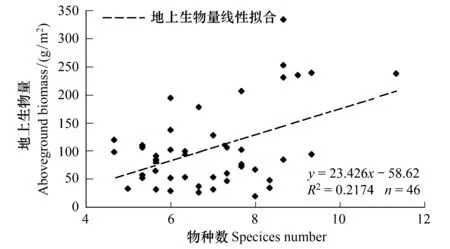
图5 地上生物量与物种数量间的关系
据图3和图4,草地样带上地上生物量与物种数量遵循大致相似的变化趋势,在空间上呈现南北高、中部低的“U”形分布。可以初步判断,两者之间可能存在相关关系。针对样地详细样方上的生物量—物种数量关系进行制图(图5),也可以清楚发现两者之间存在正相关。但相关程度高低及是否能据此建立生物量—物种数量经验模型,还需要进一步分析。
相关分析表明,在乌兰巴托—锡林浩特草地样带尺度上,样地地上生物量与物种数量程序显著正相关(r=0.467,P=0.01),即随着草地植物物种数量的增加,样带上草地地上生物量也会相应升高。但是,线性回归分析得到的决定系数(R2=0.2174)并不大,即仅有22%的地上生物量变化可以通过物种数量变化来解释,回归模型的拟合程度相对较低。这也表明,决定样地地上生物量的决定性因子可能并不是物种数量,而是具体的草地植物物种类型等其他因素。
2.5 物种数量与温度、降水的关系
针对样地上的物种数量及相应点位的温度、降水因子开展简单相关分析(表1)发现:样带上物种数和夏季月均温呈显著负相关关系(r=-0.363,P=0.013),即样带上物种数量会随着夏季月均温度的升高而减少。物种数与夏季降水量偏呈显著正相关(r=0.462,P<0.01),即样带上物种数量随着降雨量的增加而增多。
但是,对样带上物种数与温度、降水做偏相关分析(表1)则发现上述关系发生了变化。样带上物种数量和夏季月均温之间关系逆转,呈现极微弱的正相关关系(r=0.057),但是这种正相关关系没有通过显著性检验(P=0.709);而物种数量与夏季降水量偏相关系数依然为正相关,但相关程度略有降低(r=0.313,P=0.037)。
显然,简单相关分析结果符合图 6、图3和图4所展示的表象规律,也是大多数前人草原生物多样性研究的一般性总结,即干旱少雨不利于草地物种数量的提高[17]。偏相关分析从本质上揭示了物种分布及数量与温度、降水等因子之间的机理。即在温度条件相同的情况下,降水增加毫无疑问可以增加物种数量;但是在降水条件相同情况下,温度提升对于物种数量的提高仍然存在有限的作用(弱正相关,且未通过检验)。事实上,在青藏高原等高寒地区,温度就是影响植物物种分布的重要因子[31];但在蒙古高原这种典型温性草原区,影响物种分布的关键因子为水分,温度的提升对于典型温性草原物种数量的影响非常有限。

表1 物种数量与夏季平均温度、降水总量的相关性Table 1 Correlation between the average temperature, precipitation and the total number of species in summer
*在 0.05 水平(双侧)上显著相关; ** 在0.01 水平(双侧)上显著相关
2.6 地上生物量与温度、降水的关系
针对样地上的草地地上生物量及相应点位的温度、降水因子开展简单相关分析(表2)发现:样带地上生物量和夏季月均温呈负相关关系(r=-0.199),但未通过统计检验(P=0.185)。地上生物量与夏季降水量偏呈显著正相关关系(r=0.560,P<0.01),即表明样带上地上生物量随着降雨量的增加而增多。

表2 地上生物量与夏季平均温度、降水总量的相关性Table 2 Correlation between the average temperature, precipitation and the aboveground biomass in summer
*在 0.05 水平(双侧)上显著相关; ** 在0.01 水平(双侧)上显著相关
对样带地上生物量与温度、降水做偏相关分析(表2)后发现上述关系发生了变化。样带上地上生物量和夏季平均温度之间的关系逆转,呈现为显著正相关、且相关系数很高(r=0.615,P<0.01)。地上生物量和夏季降水总量之间则也呈现相关系数更高的显著正相关关系(r=0.745,P<0.01)。
显然,简单相关分析结果符合图 7、图3和图4所展示的表象,验证了大多数前人对草原地上生物量研究的一般性总结,即干旱少雨不利于草地地上生物量的增加[17, 32]。偏相关分析则从本质上揭示了地上生物量与温度、降水等因子之间的机制。即在温度条件相同的情况下,降水增加毫无疑问可以增加地上生物量;而在水分条件相同的情况下,温度的提升也依然有利于促进植物生长,有利于地上生物量的增加,这种规律突出地反映在青藏高原等高寒草原地区[31]。
2.7 多元回归模型的建立
相关分析已经表明了蒙古高原乌兰巴托—锡林郭勒草地样带上草地物种数量(SC)、草地地上生物量(AB)等2个因子与夏季温度(T)、夏季降水(P)等环境因子有着显著的相关关系。进一步考虑到温度和降水因子之间交互作用,即湿润度水平(可表达为P/T)的影响,可以在SPSS软件支持下,对征草地植被特征(物种数量和地上生物量)进行非线性回归建模,最终建立基于温度和降水特征参数的回归方程。
方差分析表明,在P=0.005的显著性水平上,基于夏季平均温度(T)、夏季降水量(P)因子建立草地物种数量的回归方程有意义。具体的回归方程为:
SC=-15.294 + 1.226×T-0.29×P+ 1.798×P/T
该回归模型的决定系数(R2)仅为0.233,这表明仅有22.3%的物种数量变化可以由温度、降水和湿润度因子变化来解释,模型总体拟合程度较低。
分析还表明,在P=0.001的显著性水平上,基于夏季平均温度(T)、夏季降水量(P)因子建立草地地上生物量的回归方程有意义。具体的回归方程为:
AB =-339.625 + 8.521×T+ 4.447×P-42.337×P/T
该回归模型的决定系数(R2)达到0.605,这表明有60.5%的地上生物量变化可以由温度、降水和湿润度因子变化来解释,模型拟合程度相对较好。
3 讨论
3.1 研究尺度所致的不确定性
由于研究区域位置、研究尺度不同,研究人员对于草原植被特征(生物多样性、植被生产力)与环境因子(温度、降水)之间的关系的研究结论多有不同[17, 31, 33- 35]。总的来看,在局地小尺度(站点、小流域尺度)环境中,温度与水分与上述植被特征呈正相关,或者关系过于复杂而呈现为没有规律;在水分制约前提下(典型者如蒙古温性草原区),大尺度(长样带、高原尺度)环境中温度因子与上述植被特征呈负相关,而水分因子仍然与植被特征呈正相关;而在温度制约环境下(典型者如青藏高寒区),大尺度(长样带、高原尺度)环境中温度、水分与上述植被特征均呈正相关。这充分体现了地学研究中的尺度推移效应,即研究区域、研究尺度的变化导致了不确定性[36]。
在本文研究中,由于目标样带跨度长,包含了多个生态地理亚区(草地类型区),同时采样间隔较短、每个生态地理亚区内均有足够数量的样地,这对于以综合的视角将局地小尺度研究规律和区域大尺度研究规律联系起来,具有一定优势。在研究方法上,简单相关分析展现了样带尺度上的生态地理变化规律,而偏相关分析则主要揭示小尺度环境中(特别是水分制约环境中)草原植物生长的环境驱动机制。这个研究有助于认识研究尺度变化所致的不确定性。
3.2 分析要素所致的不确定性
本文使用了草原植物物种数量绝对值作为衡量生物多样性的指标、地上生物量绝对值作为衡量生物生产力的指标,使用了夏季月均温度和降水总量作为驱动生态系统特征参数变化的气候环境指标。显然,尚有许多重要生态特征参数和地理环境参数还未纳入评估范围。事实上,分析要素数量的多少和具体要素的选择差异,也将会给有关研究结果带来不确定性[35, 37]。
一方面,环境要素的选择对于评估模型的具体形态及模拟精度会有重要影响。在蒙古高原这一水分制约环境中,针对物种数量、地上生物量这2个草地植被特征参数,如进一步开展包括水热条件在内的更多的环境因子,如将海拔高度、局地地形地貌、土壤类型、土壤养分和水分、风场强度和风向、太阳辐照等要素纳入进来;或者是在考虑总量基础上,进一步考虑时间动态分布特征(如降雨的时间分布);或者是进一步考虑各要素之间的协同、耦合作用(如显性地分析温度-水分耦合要素,即湿度/干燥度因子)。毫无疑问,这些深入的分析工作,将可得到更加复杂,但相关性更好、决定系数更大、可信度更高评估模型。
另一方面,草地植被特征参数的选择对于评估模型的构建也会有重要影响。例如,研究已经表明,不同地区草地群落地上地下生物量的比例有很大不同,在一些地区地上生物量和地下生物量呈正相关,而另一些地区地上生物量和地下生物量没有明显相关性,甚至是负相关[37- 38]。因此,如果单独考虑地下生物量、或者将地上和地下部分生物量同时考虑,蒙古高原样带草原生产力与气候因子的关系可能会发生很大变化。此外,对于其它重要生态系统特征参数,如均匀度指数、植被盖度、植被营养元素和指示同位素(如δ13C)等,这些生态系统特征要素与环境因子的关系会更加复杂,需要更加细致和深入研究。
4 结论
以长约1100 km的蒙古高原乌兰巴托—锡林浩特草地样带为研究区,本文描述了夏季温度、降水、物种数量、地上生物量等要素的空间变化规律,分析了物种数量、地上生物量等草地样带植被特征参数与水热因子之间的相关关系,构建了相应的回归模型,并对研究区域、研究尺度和分析要素变化所致的不确定性进行了讨论。主要结论有:
(1)蒙古高原乌兰巴托—锡林浩特草地样带上的夏季降水总量、物种数和地上生物量,呈现明显的“U” 型分布,夏季月均度则呈现明显倒“U” 型分布。生态景观类型为典型温性草原的样带南部和北部地区,其生物多样性、地上生物量明显好于呈现为温性荒漠草原、温性荒漠景观的样带中部地区。物种数量的局地空间变异性,是样带南部、北部劣于样带中部;地上生物量的局地空间变异性,则是样带南部、北部好于样带中部。
(2)应用简单相关分析方法,可以得到蒙古高原乌兰巴托—锡林浩特样带尺度上植被特征与水热环境因子间的关系:植物物种数量、地上生物量与夏季月均温度均呈现负相关,而与夏季降水总量则呈现正相关关系。应用偏相关分析,可以得到小尺度上植被特征与水热环境因子间的关系:温度和降水要素对于植物物种数量、地上生物量均呈现正效应,温度和降水的增加从本质上有利于植物物种数量、地上生物量的增加。
(3)由于尺度推移效应,在不同地理区域、不同研究尺度上针对植被特征与环境因子间关系的研究会有差异。本研究通过较长的样带考察、较密集的采样计划,将局地小尺度研究规律和区域大尺度研究规律联系起来,有助于认识研究尺度变化所致的不确定性。此外本研究还指出,分析要素数量的多少和具体要素的选择差异,也将会给有关研究结果带来不确定性。
[1] 陈灵芝, 钱迎倩. 生物多样性科学前沿. 生态学报, 1997, 17(6): 565- 572.
[2] 叶笃正, 丑纪范, 刘纪远, 张增祥, 王一谋, 周自江, 鞠洪波, 黄签. 关于我国华北沙尘天气的成因与治理对策. 地理学报, 2000, 55(5): 513- 521.
[3] 刘桂香, 赵一之, 徐杰. 蒙古高原鸦葱属植物分类和地理分布研究. 中国草地, 2001, 23(2): 12- 18.
[4] 王铁娟, 赵一之. 蒙古高原绣线菊属植物区系地理成分及其生态地理分布规律的研究. 植物研究, 2001, 21(2): 245- 251.
[5] 燕玲, 宛涛, 乌云. 蒙古高原葱属植物种皮微形态研究. 内蒙古农业大学学报, 2000, 21(1): 91- 95.
[6] 徐杰, 赵一之, 田桂泉. 蒙古高原天门冬属植物分支系统演化的研究. 内蒙古大学学报: 自然科学版, 2003, 24(3): 325- 329.
[7] 王菱, 甄霖, 刘雪林, Batkhishig O, 王勤学. 蒙古高原中部气候变化及影响因素比较研究. 地理研究, 2008, 27(1): 171- 180.
[8] 张雪艳, 胡云锋, 庄大方, 齐永清. 蒙古高原NDVI的空间格局及空间分异. 地理研究, 2009, 28(1):10- 18.
[9] Zhang X Y, Gong S L, Zhao T L, Arimoto R, Wang Y Q, Zhou Z J. Sources of Asian dust and role of climate change versus desertification in Asian dust emission. Geophysical Research Letters, 2003, 30(24): 2272- 2272.
[10] 刘纪远, 齐永青, 师华定, 庄大方, 胡云锋. 蒙古高原塔里亚特-锡林郭勒样带土壤风蚀速率的137Cs 示踪分析. 科学通报, 2007, 52(23): 2785- 2791.
[11] Waide R B, Willig M R, Steiner C F, Mittelbach G, Gough L, Dodson S I, Juday G P, Parmenter R. The relationship between productivity and species richness. Annual Review of Ecology and Systematics, 1999, 30: 257- 300.
[12] Mittelbach G G, Steiner C F, Scheiner S M, Gross K L, Reynolds H L, Waide R B, Willig M L, Dodson S I, Gough L. What is the observed relationship between species richness and productivity? Ecology, 2001, 82(9): 2381- 2396.
[13] 刘先华, 李凌浩, 陈佐忠. 内蒙古锡林河流域植被多样性特点及其与气候因子的关系. 植物生态学报, 1998, 22(5): 466- 472.
[14] 王国杰, 汪诗平, 郝彦宾, 蔡学彩. 水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响. 生态学报, 2005, 25(7): 1649- 1656.
[15] 郑晓翾, 靳甜甜, 木丽芬, 刘国华. 呼伦贝尔草原物种多样性与生物量、环境因子的关系. 中国草地学报, 2008, 30(6): 74- 81.
[16] 白永飞, 张丽霞, 张焱, 陈佐忠. 内蒙古锡林河流域草原群落植物功能群组成沿水热梯度变化的样带研究. 植物生态学报, 2002, 26(3): 308- 316.
[17] 白永飞, 李凌浩, 王其兵, 张丽霞, 张焱, 陈佐忠. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667- 667.
[18] 胡云锋, 燕艳, 阿拉腾图雅, 毕力格吉夫. 内蒙古东北-西南草地样带植物多样性变化. 资源科学, 2012, 34(6): 1024- 1031.
[19] 蔡学彩, 李镇清, 陈佐忠, 王义凤, 汪诗平, 王艳芬. 内蒙古草原大针茅群落地上生物量与降水量的关系. 生态学报, 2005, 25(7): 1657- 1662.
[20] 马文红, 杨元合, 贺金生, 曾辉, 方精云. 内蒙古温带草地生物量及其与环境因子的关系. 中国科学C辑: 生命科学, 2008, 38(1): 84- 92.
[21] 陈效逑, 郑婷. 内蒙古典型草原地上生物量的空间格局及其气候成因分析. 地理科学, 2008, 28(3): 369- 374.
[22] Fay P A, Carlisle J D, Knapp A K, Blair J M, Collins S L. Productivity responses to altered rainfall patterns in a C-4-dominated grassland. Oecologia, 2003, 137(2): 245- 251.
[23] Fay P A. Precipitation variability and primary productivity in water-limited ecosystems: how plants ‘leverage’ precipitation to ‘finance’ growth. New Phytologist, 2009, 181(1): 5- 8.
[24] Heisler-White J L, Knapp A K, Kelly E F. Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia, 2008, 158(1): 129- 140.
[25] Zhang B C, Cao J J, Bai Y F, Zhou X H, Ning Z G, Yang S J, Hu L. Effects of rainfall amount and frequency on vegetation growth in a Tibetan alpine meadow. Climatic Change, 2013, 118(2): 197- 212.
[26] 白永飞. 降水量季节分配对克氏针茅草原群落初级生产力的影响. 植物生态学报, 1999, 23(2): 155- 160.
[27] 王玉辉, 周广胜. 内蒙古羊草草原植物群落地上初级生产力时间动态对降水变化的响应. 生态学报, 2004, 24(6): 1140- 1145.
[28] 周双喜, 吴冬秀, 张琳, 施慧秋. 降雨格局变化对内蒙古典型草原优势种大针茅幼苗的影响. 植物生态学报, 2010, 34(10): 1155- 1164.
[29] Uno I, Wang Z, Chiba M, Chun Y S, Gong S L, Hara Y, Jung E, Lee S S, Liu M, Mikami M, Music S, Nickovic S, Satake S, Shao Y, Song Z, Sugimoto N, Tanaka T, Westphal D L. Dust model intercomparison (DMIP) study over Asia: Overview. Journal of Geophysical Research: Atmospheres, 2006, 111(D12), doi: 10.1029/2005JD006575.
[30] Zhang X Y, Gong S L, Zhao T L, Arimoto R, Wang Y Q, Zhou Z J. Sources of Asian dust and role of climate change versus desertification in Asian dust emission. Geophysical Research Letters, 2003, 30(24): 2272- 2272. (本条文献与第9条重复)
[31] 杨元合, 饶胜, 胡会峰, 陈安平, 吉成均, 朱彪, 左闻韵, 李轩然, 沈海花, 王志恒, 唐艳鸿, 方精云. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系. 生物多样性, 2004, 12(1): 200- 205.
[32] 韩彬, 樊江文, 钟华平. 内蒙古草地样带植物群落生物量的梯度研究. 植物生态学报, 2006, 30(4): 553- 562.
[33] 龙慧灵, 李晓兵, 王宏, 魏丹丹, 张程. 内蒙古草原区植被净初级生产力及其与气候的关系. 生态学报, 2010, 30(5): 1367- 1378.
[34] 马文红, 方精云, 杨元合, 安尼瓦尔·买买提. 中国北方草地生物量动态及其与气候因子的关系. 中国科学: 生命科学, 2010, 40(7): 632- 641.
[35] 郑晓翾, 赵家明, 张玉刚, 吴雅琼, 靳甜甜, 刘国华. 呼伦贝尔草原生物量变化及其与环境因子的关系. 生态学杂志, 2007, 26(4): 533- 538.
[36] 李明财, 朱教君, 孙一荣. 植物对气候变化生理生态响应的不确定性分析. 西北植物学报, 2009, 29(1): 207- 214.
[37] 鄢燕, 张建国, 张锦华, 范建容, 李辉霞. 西藏那曲地区高寒草地地下生物量. 生态学报, 2005, 25(11): 2818- 2823.
[38] 王长庭, 曹广民, 王启兰, 景增春, 丁路明, 龙瑞军. 青藏高原高寒草甸植物群落物种组成和生物量沿环境梯度的变化. 中国科学C辑: 生命科学, 2007, 37(5): 585- 592.
The relationship between vegetation characteristics and hydro-thermic factors along the Ulanbattar-Xilinhot Grassland Transect of the Mongolian Plateau
HU Yunfeng1,*, Batunacun1,2, Biligejifu3, LIU Jiyuan1, ZHEN Lin1
1InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China2RenMinUniversityofChina,Beijing100872,China3InnerMongoliaRangelandSurvey&DesignInstitute,Hohhot010051,China
During August of 2012, the research team carried out a field exploration of the Ulanbattar-Xilinhot Grassland Transect on the Mongolian Plateau. The transect was split into three parts according to the physical geographical conditions and practical administrative boundaries: the northern section (from Ulanbattar to Airag), the central section (from Airag to Sunitezuoqi), and the southern section (from Sunitezuoqi to Xilinhot). Along the 1100 km transect, 46 sample areas, 136 grass yield quadrats, and about 500 species-frequency plots were surveyed in detail and a series of species number data and above ground biomass data were then measured. Using GIS software and the GHCN datasets (The Global Historical Climatology Network), the historical meteorological data along the above transects were then extracted. The monthly average temperatures and total precipitation during the summer season (i.e., June, July and August) were calculated. Based on the vegetation characteristics data and hydro-thermic factor data, the correlation and partial correlation were analysed. The results show that: (1) the distribution of monthly average temperatures in summer are presented as an inverted "U" pattern along the transect, i.e., the northern and southern sections showing lower temperatures, while the central section maintained a high temperature level. Conversely, the distribution of total precipitation in summer is shown as an upstanding “U” pattern, i.e., high precipitation in the northern and southern sections, with low precipitation in the central section; (2) the distribution of numbers of plant species and above ground biomass are both depicted as an upstanding "U" shape along the transect. In the southern and northern regions, where the ecological type is typically warm steppe, both the bio-diversity and aboveground biomass are significantly better than those in the central region, where the ecological type is usually warm temperate desert grassland or temperate desert; (3) simple correlation analysis normally reflects the relationship between vegetation characteristics and hydro-thermic environmental factors over large scales (for example, plateau transect). In this study, the numbers of plant species and above ground biomass are negatively correlated with the summer monthly average temperature and are positively correlated with the summer precipitation; (4) the partial correlation analysis is inclined to reflect the relationship among different factors over a local scale (for example, small watershed). In this study, both the temperature and precipitation factors have positive effects on the number of plant species and the ground biomass.
Mongolian Plateau; grassland transect; vegetation characteristic; environmental factors; correlation analysis
中国科学院知识创新方向性项目(KZCX2-EW- 306); 国家自然科学基金项目(40971223); 科技部973计划(2010CB950904)
2013- 08- 09;
2014- 05- 30
10.5846/stxb201308092051
*通讯作者Corresponding author.E-mail: huyf@lreis.ac.cn
胡云锋, 巴图娜存, 毕力格吉夫, 刘纪远, 甄霖.乌兰巴托—锡林浩特样带草地植被特征与水热因子的关系.生态学报,2015,35(10):3258- 3266.
Hu Y F, Batunacun, Biligejifu, Liu J Y, Zhen L.The relationship between vegetation characteristics and hydro-thermic factors along the Ulanbattar-Xilinhot Grassland Transect of the Mongolian Plateau.Acta Ecologica Sinica,2015,35(10):3258- 3266.
