长江口水域夏季鱼卵和仔稚鱼年间变化
2015-01-20刘守海王金辉刘材材秦玉涛刘志国邓邦平
刘守海,王金辉,刘材材,*,秦玉涛,刘志国,邓邦平
1 国家海洋局东海环境监测中心,上海 200137 2 国家海洋局海洋赤潮灾害立体监测技术与应用重点实验室,上海 200137
长江口水域夏季鱼卵和仔稚鱼年间变化
刘守海1,2,王金辉1,2,刘材材1,2,*,秦玉涛1,2,刘志国1,2,邓邦平1,2
1 国家海洋局东海环境监测中心,上海 200137 2 国家海洋局海洋赤潮灾害立体监测技术与应用重点实验室,上海 200137
基于2005 年、2008 年、2009 年和2011 年8 月(夏季)在长江口水域(30°30′—31°45′ N,121°15′—123°10′ E) 4 个航次的浮游生物拖网资料,分析了长江口水域鱼卵和仔稚鱼的种类组成、数量分布特征及其年间变化。结果表明:4 个航次采集的鱼卵和仔稚鱼鉴定到种的种类有17 种,隶属于8 目13 科,以鲈形目种类最多,11 种,其次是鲱形目,5 种,其他各目种类均小于5 种;种类数存在明显年间差异,2005 年种类数最多(鱼卵3 种,仔稚鱼8 种),其次是2009 年和2011 年,2008 年种类数最少(鱼卵1种,仔稚鱼5 种)。优势种年间更替明显,长蛇鲻(Sauridaelongata)、虾虎鱼(Gobiidae spp.)和中华小公鱼(Stolephoruschinensis)在2005 年是优势种,2008 年优势种为鳀鱼(Engraulisjaponicus),2009 年优势种为鳀鱼、寡鳞飘鱼(Pseudolaubucaengraulis)、虾虎鱼等,2011 年虾虎鱼和小公鱼(包括小公鱼属未定种Stolephorusspp.和中华小公鱼Stolephoruschinensis)成为优势种。2005 年鱼卵和仔稚鱼数量分布的密集区在嵊泗列岛附近水域,2008 年鱼卵和仔稚鱼出现较少,未出现明显的数量密集区;2009 年鱼卵数量较少,仔稚鱼数量较多,密集区主要分布在在长江口以外123° E附近水域;2011 年鱼卵主要分布在在长江北支口门外附近水域,仔稚鱼在调查区内分布相对均匀。
长江口; 鱼卵; 仔稚鱼; 年间变化
长江口水域由于长江径流带来了大量的营养物质,孕育了丰富的饵料资源,使之成为多种经济鱼类的产卵、索饵和育幼的重要场所,也是我国重要的渔场[1]。三峡水库2006 年夏季基本完成蓄水,2006 年9 月水位提高到156 m,开始运行[2]。三峡水库的建成运行改变了长江径流的季节变化规律,使得夏季长江径流量减少,改变了长江口水域的生态环境,并将进一步影响该水域渔业资源的补充[3]。
国外学者针对上游水库建设对河口鱼卵和仔稚鱼的影响开展了大量研究:如Faria等[4]研究发现,欧洲最大的阿尔克瓦水坝的使用引起瓜迪亚纳河口及其邻近水域盐度的变化,从而成为影响鱼卵、仔稚鱼丰度和分布的重要因素。Rowell等[5]研究发现,科罗拉多河(Colorado River)大坝的修建对河口生活的石首鱼科鱼类麦氏托头石首鱼(Totoabamacdonaldi)种群生长有明显的变化,大坝修建前的麦氏托头石首鱼幼鱼生长速率是修建后的两倍。Doornbos[6]对赫雷弗灵恩(Grevelingen)河口上游大坝截水前后的研究也发现鱼类群落发生了变化。
三峡工程建设后,我国学者针对该水域也开展了广泛的研究,王金辉等[7]研究发现,由于三峡工程建设,长江口门以内区域浮游生物种类明显下降,浮游生物群落结构发生变化,硅藻比重减少,甲藻增加。丁月旻等[8]研究结果表明,2007 年和2009 年秋季长江口鱼卵和仔稚鱼群落种类、丰度和多样性均有显著提高,并且群落结构发生显著变异。三峡工程对河口鱼卵和仔稚鱼群落变化的研究也有报道,例如刘淑德等[9]、丁月旻等[8]分别对春季和秋季的长江口鱼卵和仔稚鱼的群落结构进行了研究,但是对径流量显著减少的夏季却鲜见报道。另外,还有长江口水域鱼卵、仔稚鱼分布的相关研究[3,8- 16],针对鱼卵、仔稚鱼年间变化的研究仅见于刘淑德等[11]和丁月旻等[8]。
本文利用三峡工程高位蓄水完成前2005 年(156 m高位蓄水前)和2008 年、2009 年、2011 年(运行后)在长江口水域的鱼卵、仔稚鱼定点调查资料,通过长江口水域鱼卵、仔稚鱼的种类组成、数量分布及其年间变化的分析,研究水库工程等人类活动对河口鱼卵和仔稚鱼群落特征的影响,为河口生态环境保护提供基础数据。
1 材料与方法
数据来源于2005 年、2008 年、2009 年以及2011 年在长江口水域(30°30′—31°45′ N,121°15′—123°10′ E)夏季(8 月)的鱼卵和仔稚鱼调查资料,调查共设20 个站位(图1)。样品采集用浅水Ⅰ型浮游生物网(口径50 cm、筛绢CQ14、孔径0.505 mm),由底至表进行垂直拖网。样品采集和处理均按照《海洋调查规范——海洋生物调查》(GB12763.6-91)进行。采集的鱼卵和仔稚鱼样品加体积分数为5%的福尔马林溶液固定保存。样品鉴定及个体计数参考《中国近海鱼卵和仔鱼》[17]进行。对于无法鉴定到种但是能够鉴定到科的物种,在记录时用科名代替;对于无法鉴定的种类用未定种命名。
2 结果
2.1 种类组成及年间变化
2005 年,2008 年,2009 年和2011 年的8 月采集的鱼卵和仔稚鱼隶属于8 目13 科,其中鉴定到种的有17 种,鉴定到属的有7 个,鉴定到科的有5 个(表1)。其中鲈形目种类数最多。其中,作为长江口重要经济种类的凤鲚(Coiliamystus)在2005 年和2011 年出现,而2008 年和2009 年都没有出现。小公鱼(Stolephorusspp.)仅出现在2005 年和2011 年,2008 年和2009 年没有出现。
2005 年鉴定到种的鱼卵有3 种,仔稚鱼有8 种;2008 年鉴定到种的鱼卵只有1 种,有5 种仔稚鱼鉴定到种;2009 年鉴定到种的鱼卵有3 种,仔稚鱼有2 种;2011 年鉴定到种的鱼卵有3 种,仔稚鱼有5 种。鱼卵种类数以2005 年和2011 年最多,仔稚鱼种类数以2005 年最多,2009 年最少(图2)。

图1 采样站位Fig.1 Location of sampling stations
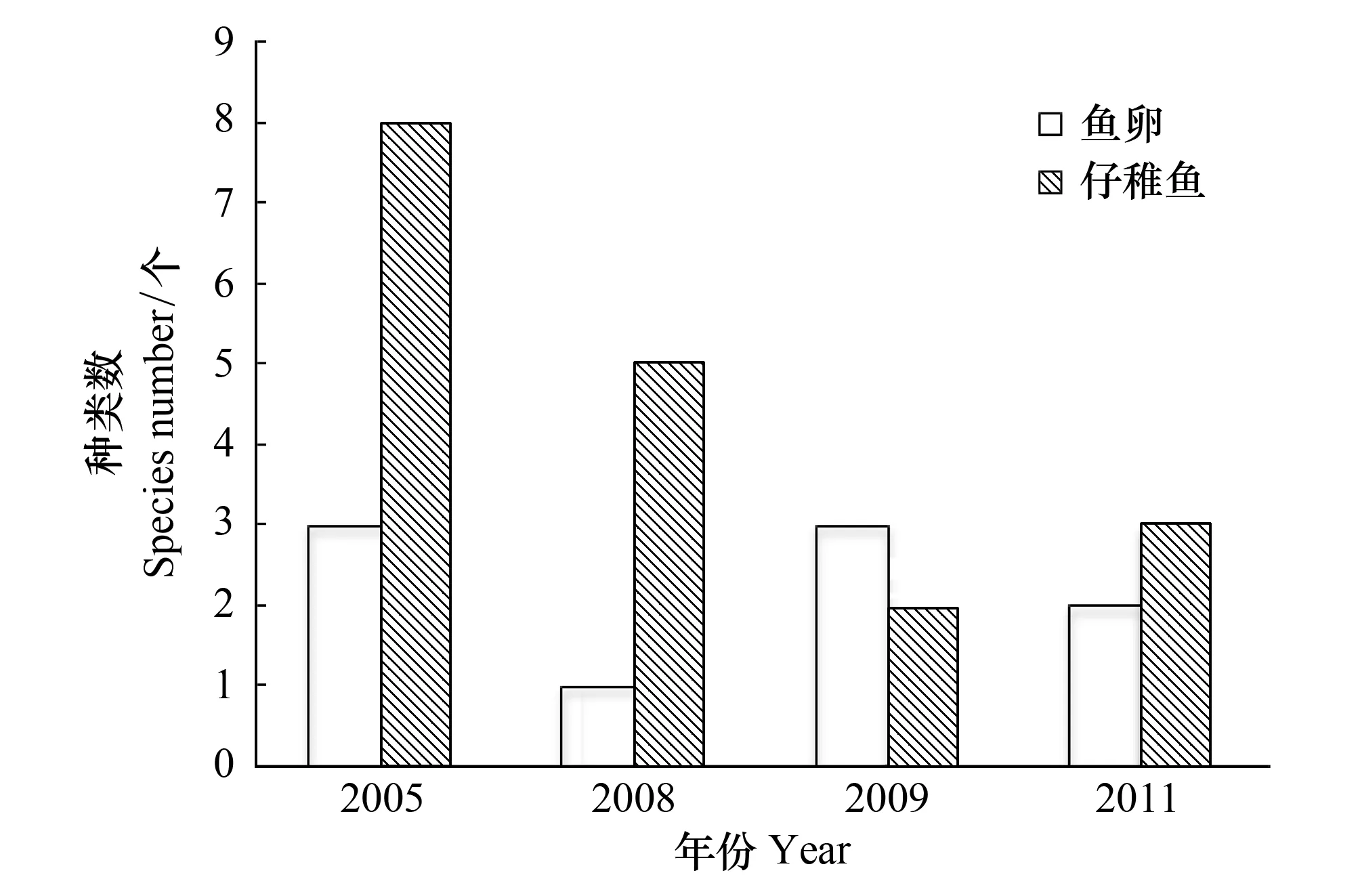
图2 鱼卵和仔稚鱼种数的年际变化 Fig.2 Inter-annual variation in species number of fish eggs, larvae and juveniles

表1 长江口水域鱼卵和仔稚鱼名录Table 1 The categories of fish eggs, larvae and juveniles in Yangtze River Estuary
2.2 鱼卵、仔稚鱼优势种类的年际变化
以个体总数比例≥10%的种类为优势种[18]。2005 年,长蛇鲻(Sauridaelongata)是鱼卵优势种,中华小公鱼(Stolephoruschinensis)和虾虎鱼(Gobiidae spp.)是仔稚鱼优势种;2008 年鱼卵和仔稚鱼的优势种均为鳀鱼(Engraulisjaponicus);2009 年鳀鱼是鱼卵优势种,仔稚鱼优势种有寡鳞飘鱼(Pseudolaubucaengraulis)、虾虎鱼等;小公鱼是2011 年鱼卵的优势种类,小公鱼和虾虎鱼是仔稚鱼的优势种(表2)。

表2 长江口水域鱼卵和仔稚鱼优势种总量比例的年际变化Table 2 Inter-annual variation in the percent of dominant species to total abundance of fish eggs, larvae and juveniles in Yangtze River Estuary
2.3 数量分布及年际变化
长江口水域鱼卵和仔稚鱼数量及其分布格局存在明显年间变化(图3,图4)。2005年有10 个站采到鱼卵,高值区出现在嵊泗列岛附近水域,口内站位也有鱼卵出现;2008 年鱼卵数量较少,20 个站位中只有2 个站采到,分别是位于长江北支口门处和嵊泗列岛附近水域;2009 年有10 个站采到鱼卵,无高值区出现;2011 年有10 个站采到鱼卵,高值出现在长江北支口门外和嵊泗列岛附近水域(图3)。比较各站位4 年鱼卵数量分布,显示嵊泗列岛和长江北支口门附近水域鱼卵数量最高。而长江口南支河口水域鱼卵数量较低,仅在2005 年出现鱼卵(图3)。

图3 长江口水域鱼卵数量分布(ind /m3)Fig.3 Distribution and abundance of fish eggs in Yangtze River Estuary

图4 长江口水域仔稚鱼数量分布(ind /m3)Fig.4 Distribution and abundance of fish larvae and juveniles in Yangtze River Estuary
2005 年仔稚鱼高值区主要分布在长江口咸淡交错水域及嵊泗列岛附近水域,主要是中华小公鱼、白姑鱼(Argyrosomusargentatus)和舌鳎属未定种(Cynoglossusspp.)等构成;2008 年只有3 个站采到仔稚鱼且丰度均较低;2009 年仔稚鱼共有12 个站采到,高值区主要分布在长江口以外123° E附近水域,主要由舌鳎属未定种组成,此外,在长江口口内水域采到寡鳞飘鱼的仔稚鱼;2011 年仔稚鱼分布较均匀(图4)。比较4 年各站位仔稚鱼数量显示,2005 年仔稚鱼数量最多,主要分布在嵊泗列岛附近水域(图4)。
3 讨论
鱼卵和仔稚鱼是鱼类生命周期中的初级阶段,也是最为脆弱的时期。海洋环境因素的细微变化将对其发育、生长直至种群的补充产生强烈影响[19]。鱼卵和仔稚鱼对环境变化敏感,对环境的响应是非线性的,且复杂。影响河口鱼卵仔稚鱼的环境因素众多,例如径流量[15]、过度捕捞[20]、环境污染[21]、生境破碎化[22]等等。研究表明,径流量是影响长江口鱼卵仔稚鱼物种组成和优势种年间变异的关键因子[20,23]。
3.1 种类及优势种的年间变化
长江流域水库工程的建成运行,改变了长江下泄径流量的变化规律[24]、造成长江来水中含沙量[25]、淡水资源[26]、叶绿素[27]、长江口基础生物资源等生态变化[7,28],对长江口生态环境,尤其是盐度环境带来显著影响。使得盐度成为决定河口鱼类空间分布的主要因子之一[29]。蒋玫等[3]研究了长江口河口及邻近水域仔稚鱼群落结构与环境因子的关系,结果表明夏季长江口仔稚鱼淡水群落的丰度与径流量和盐度的相关系数较高,长江口仔稚鱼咸水群落的丰度与盐度的相关系数较高。认为盐度可能是造成长江河口仔稚鱼群落结构时空差异性的主要原因。
2005年鱼卵和仔稚鱼的种类数均大于运行后(2008 年、2009 年和2011 年)的种类数(表1和图1)。蒋玫等[15]认为对于径流量相对较小的年份,鱼卵数量明显地减少,分布范围内移缩小。洪峰减少,径流量变化幅度趋缓,引起的海洋水系分布及消长变化,进一步影响鱼卵、仔稚鱼的种类数变化[28]。在三峡工程运行后的2008 年和2009 年,由于夏季冲淡水势力的减弱,外海水团对河口的入侵,造成咸淡水区域的西移,外海种随着外海水团西移,导致调查海域出现的种类多为外海种,例如2008 年和2009 年的鱼卵主要优势种为鳀鱼,比例均超过50%(表2)。河口沿岸优势种中华小公鱼在三峡工程建成运行前的2005 年数量较多,运行后的2008 年和2009 年则没有出现,到2011 年中华小公鱼数量有所上升,小公鱼属未定种重新成为优势种类(表2)。2005—2011年,鱼卵仔稚鱼优势种类发生了变化。
有学者认为长江口仔稚鱼某些优势种的变化甚至消失,可能与三峡大坝建设期径流量和泥沙量的调控变动有关[3]。三峡水库蓄水后,影响径流分布,从而改变了长江口及其邻近海域的理化和生物因子的原因,由于河口环境条件波动较大,鱼卵和仔稚鱼的种类、数量、优势种的结构也会相应地发生变动,并且部分鱼类的生殖时间上也出现了相应的改变[13]。长江径流量改变,造成了河口区域鱼卵和仔稚鱼优势种类的变化。综上所述,由于三峡工程对来水资源的调蓄作用,导致长江径流量的减少,造成外海水团对河口的入侵,对河口渔场环境造成了明显影响,表现为8 月河口种沿岸种的减少,外海种数量增加。
3.2 数量分布的年间变化
长江流域上的筑库建坝、截流引水,改变了径流和泥沙输运原有的季节性与年际变化的格局,引起生物群落的变化[30]。例如,有研究表明,与三峡工程蓄水前对比,悬浮体高值区的界限往西移动了近半个经度,同季节的叶绿素含量的平均值降低[27]。另有研究表明径流量的输入对长江口浮游植物丰度分布尤其是近岸低盐性的中肋骨条藻(Skeletonemacostatum)分布有着决定性的影响[31-32]。鱼类产卵中心位置的变化随长江入海流量多少和影响的盐度、透明度变化而趋近岸或离岸移动[1]。
本研究中,2008 年鱼卵和仔稚鱼的数量较2005 年有显著降低。2005 年鱼卵和仔稚鱼数量的高值区出现在嵊泗列岛附近水域;2008 年鱼卵和仔稚鱼出现较少,未出现明显的高值区;2009 年鱼卵数量较少,而仔稚鱼出现较多,高值区在长江口以外123° E附近水域;2011 年鱼卵高值出现在长江北支口门外附近水域,仔稚鱼多分布较均匀(图3和图4)。鱼卵和仔稚鱼丰度的变化与环境变化密切相关。Faria等[4]对瓜迪亚纳河口研究,认为径流变化将对河口鱼卵和仔稚鱼产生影响,径流减少,河口的营养物质不能被运输到河岸,河岸边的仔稚鱼数量也减少。
三峡大坝建成后,由于8 月长江径流量的减少,也使得入海泥沙量同步减少,引起了河口水域海水透明度的增加,可能使浮游植物高生产力区向海岸扩展,许多鱼类的产卵索饵场位置均会有相应的变化[33]。三峡水库蓄水后的2004 年长江口春季鱼卵和仔稚鱼丰度迅速下降,仅为1999 年的13.9%和2001 年的4.3%,2007 年略有回升;鱼卵和仔稚鱼群落结构年度间差异显著,2004 年鱼卵和仔稚鱼群落结构与1999 年和2001 年相异性最高,群落相似性以2004 和2007 年最高,达71.17%。与蓄水前相比,蓄水后的长江口鱼卵和仔稚鱼群落多样性也显著下降[34]。本研究中,2009 年和2011 年鱼卵和仔稚鱼的数量、分布区域均略有回升,且2011 年鱼卵的高值出现在长江北支口门外附近水域,较2005 年的高值区有西移动的趋势,可能与水库蓄水,径流量减少,外海水团接近河口有关,河口水域海水透明度的增加。
[1] 杨东莱, 吴光宗, 孙继仁. 长江口及其邻近海区的浮性鱼卵和仔稚鱼的生态研究. 海洋与湖沼, 1990, 21(4): 346- 355.
[2] 梁福庆. 三峡库区生态环境建设成效与保护对策. 人民长江, 2010, 41(13): 28- 32.
[3] 蒋玫, 李磊, 沈新强, 全为民. 2010-2011年长江河口及邻近水域春夏季仔稚鱼群落结构及其多样性的研究. 海洋学报, 2014, 36(6): 131- 137.
[4] Faria A, Morais P, Chícharo M A. Ichthyoplankton dynamics in the Guadiana estuary and adjacent coastal area, South-East Portugal. Estuarine, Coastal and Shelf Science, 2006, 70(1/2): 85- 97.
[5] Rowell K, Flessaa K W, Dettmana D L, Román M J, Gerber L R, Findley L T. Diverting the Colorado River leads to a dramatic life history shift in an endangered marine fish. Biological Conservation, 2008, 141(4): 1138- 1148.
[6] Doornbos G. Changes in the fish fauna of the former Grevelingen Estuary, before and after the closure in 1971. Hydro Biological Bulletin, 1992(16): 279- 283.
[7] 王金辉, 徐韧, 秦玉涛, 孙亚伟, 刘材材. 长江口基础生物资源现状及年际变化趋势分析. 中国海洋大学学报, 2006, 36(5): 821- 828.
[8] 丁月旻, 线薇微. 秋季长江口鱼类浮游生物群落时空结构. 中国海洋大学学报, 2011, 41(10): 67- 74.
[9] 刘淑德, 线薇微, 刘栋. 春季长江口及其邻近海域鱼类浮游生物群落特征. 应用生态学报, 2008, 19(10): 2284- 2292.
[10] 徐兆礼, 袁骐, 蒋玫, 臧增加. 长江口鱼卵和仔、稚鱼的初步调查. 中国水产科学, 1999, 6(5): 63- 64.
[11] 朱鑫华, 刘栋, 沙学绅. 长江口春季鱼类浮游生物群落结构与环境因子的关系. 海洋科学集刊, 2002, 44: 169- 178.
[12] 李建生, 胡芬, 程家骅. 长江口水域春季鱼卵仔鱼分布及其与温度盐度的关系. 海洋科学, 2007, 31(4): 16- 21.
[13] 单秀娟, 线薇薇, 武云飞. 三峡工程蓄水前后秋季长江口鱼类浮游生物群落结构的动态变化初探. 中国海洋大学学报, 2005, 35(6): 936- 940.
[14] 王金辉, 孙亚伟, 刘材材, 秦玉涛, 程祥圣, 徐韧. 长江口鱼卵仔鱼资源现状的调查与分析. 海洋学研究, 2007, 25(4): 40- 50.
[15] 蒋玫, 沈新强, 陈莲芳. 长江口及邻近水域春季鱼卵仔鱼分布与环境因子的关系. 海洋环境科学, 2006, 25(2): 37- 44.
[16] 蒋玫, 沈新强. 长江口及邻近水域夏季鱼卵、仔鱼数量分布特征. 海洋科学, 2006, 30(6): 92- 97.
[17] 张仁斋, 陆穗芬, 赵传絪, 陈莲芳, 臧增嘉, 姜言伟. 中国近海鱼卵与仔鱼. 上海: 上海科学技术出版社, 1985.
[18] 蒋玫, 王云龙, 林钦, 徐兆礼, 沈新强, 袁骐. 洋山深水港水域鱼卵仔鱼分布特征. 海洋环境科学, 2008,27(1): 43- 46.
[19] 万瑞景, 孙珊. 黄、东海生态系统中鱼卵、仔稚幼鱼种类组成与数量分布. 动物学报, 2006, 52(1): 28- 44.
[20] 单秀娟, 线薇薇, 武云飞. 长江河口生态系统鱼类浮游生物生态学研究进展. 海洋湖沼通报, 2004, (4): 87- 93.
[21] 姚野梅. 长江口石油类污染状况调查. 上海水产大学学报, 1995, 4(3): 225- 230.
[22] 叶属峰, 吕吉斌, 丁德文, 黄秀清. 长江口大型工程对河口生境破碎化影响的初步研究. 海洋工程, 2004, 22(3): 41- 47.
[23] Morais P, Chícharo M A, Chícharo L. Changes in a temperate estuary during the filling of the biggest European dam. Science of the Total Environment, 2009, 407(7): 2245- 2259.
[24] 史赟荣. 长江口鱼类群落多样性及基于多元排序方法群落动态的研究[D]. 上海: 上海海洋大学, 2012.
[25] 翟世奎, 张怀静, 范德江, 杨荣民, 曹立华. 长江口及其邻近海域悬浮物浓度和浊度的对应关系. 环境科学学报, 2005, 25 (5): 693- 699.
[26] 曹勇, 陈吉余, 张二凤, 陈沈良, 曹卫承. 三峡水库初期蓄水对长江口淡水资源的影响. 水科学进展, 2006, 17(4): 554- 558.
[27] 邵和宾, 范德江, 张晶, 王亮, 张喜林, 褚忠信. 三峡大坝启用后长江口及邻近海域秋季悬浮物、叶绿素分布特征及影响因素. 中国海洋大学学报, 2012, 42(5): 94- 104.
[28] 蒋玫, 沈新强, 王云龙, 袁骐, 陈莲芳. 长江口及其邻近水域鱼卵、仔鱼的种类组成与分布特征. 海洋学报, 2006, 28(2): 171- 174.
[29] Krumme U. Patterns in tidal migration of fish in a Brazilian mangrove channel as revealed by a split-beam echosounder. Fisheries Research, 2004, 70(1): 1- 15.
[30] 刘淑德. 长江口及其邻近海域鱼类浮游生物群落结构特征研究[D]. 青岛: 中国科学院研究生院, 2009.
[31] 王俊, 陈瑞盛, 左涛. 三峡工程截流后长江口邻近海域的网采浮游植物群落结构特征. 水生态学杂志, 2009, 2(2): 80- 87.
[32] 唐峰华, 伍玉梅, 樊伟, 沈新强, 王云龙. 长江口浮游植物分布情况与径流关系的初步探讨. 生态环境学报, 2010, 19(12): 2934- 2940.
[33] 庄平, 王幼槐, 李圣法, 邓思明, 李长松, 倪勇, 章龙珍, 张涛, 冯广朋, 凌建忠. 长江口鱼类. 上海: 上海科学技术出版社, 2006.
[34] 刘淑德, 线薇薇. 三峡水库蓄水前后春季长江口鱼类浮游生物群落结构特征. 长江科学院院报, 2010, 27(10): 82- 87.
Inter-annual variation in pelagic fish egg, larval, and juvenile assemblages during summer in the Yangtze River Estuary, China
LIU Shouhai1,2, WANG Jinhui1,2, LIU Caicai1,2,*, QIN Yutao1,2, LIU Zhiguo1,2, DENG Bangping1,2
1EastChinaSeaEnvironmentalMonitoringCenter,StateOceanicAdministration,Shanghai200137,China
2KeyLaboratoryofIntegratedMonitoringandAppliedTechnologyforMarineHarmfulAlgalBlooms,StateOceanicAdministration,Shanghai200137,China
Important fish life history and ecological information may be obtained from fish eggs, larvae, and juveniles; therefore, these developmental stages play an important role in environment impact assessments, fishery stock analyses, fish propagation, seeding release, and fish farming. Fish eggs, larvae, and juveniles are mainly found in near-surface waters. Growth and survival during the earliest life stages of fishes are important, because variability in survival during these early developmental stages may lead to great differences in the number of recruits that survive to catchable size. Physical factors, such as ocean circulation, frontal systems, and turbulence, represent major parameters that control the survival and growth of fish eggs, larvae and juveniles. The Yangtze River Estuary is an important spawning ground for many commercially important fishes. Furthermore, salinity and temperature have also been shown to be the most important parameters affecting the distribution and abundance of fish eggs, larvae, and juveniles. In recent decades, the Yangtze River Estuary has been subject to major human-induced environmental changes, particularly following the construction of the Three Gorges Dam in the upper reaches of the river. Negative impacts on fish egg, larval, and juvenile assemblages have been observed in other estuaries after dam construction. However, the potential impact of environmental changes on the fish egg, larval, and juvenile assemblages in the Yangtze River Estuary remain unclear. We investigated the fish egg, larval, and juvenile assemblages in the Yangtze River Estuary (30°30′—31°45′ N, 121°15′—123°10′ E) in the summers of 2005, 2008, 2009, and 2011. Species composition, distribution pattern, and annual variation in fish eggs, larvae, and juveniles were studied. The results showed that a total of 17 fish species were present in the river. These species belonged to 8 orders and 13 families. Perciformes was the most diverse order, with 11 species; the second group was Cluperformes with 5 species, while all other groups had less than 5 species. The highest number of species was recorded in the summer of 2005 (3 fish eggs, 8 fish larvae), while the lowest number of species was recorded in the summer of 2008 (1 fish egg, 5 fish larvae). Yearly succession of dominant species was observed. For example, sedentary estuarine or coastal species, includingSauridaelongate, Gobiidae spp., andStolephoruschinensis, were the numerically dominant species in 2005. In comparison, the marine migratory speciesEngraulisjaponicuswas the dominant species in both 2008 and 2009. Yet,Stolephorusspp. regained dominance in 2011. Fish eggs, larvae, and juveniles were mainly distributed in the waters adjacent to Shengsi Archipelago in 2005. The abundance of fish eggs, larvae, and juveniles was low in 2008, with no noticeable peak being documented in the study area. In 2009, although the abundance of fish eggs was low, the abundance of fish larvae and juveniles increased compared to 2008. Fish larvae and juveniles were mainly distributed in the region near 123°E, off the Yangtze River Estuary. The peak abundance of fish eggs in 2011 was near the mouth of the North branch of the river. In general, salinity may be the most important factor influencing the spatial distribution of fish eggs, larvae, and juveniles in the Yangtze River Estuary.
the Yangtze River Estuary; fish eggs; fish larvae and juveniles; inter-annual variation
海洋公益性行业科研专项(201305027); 海洋赤潮灾害立体监测技术与应用国家海洋局重点实验室资助课题(MATHAB201303, MATHAB201402)
2013- 10- 27;
日期:2015- 04- 14
10.5846/stxb201310272588
*通讯作者Corresponding author.E-mail: liucaicai@eastsea.gov.cn
刘守海,王金辉,刘材材,秦玉涛,刘志国,邓邦平.长江口水域夏季鱼卵和仔稚鱼年间变化.生态学报,2015,35(21):7190- 7197.
Liu S H, Wang J H, Liu C C, Qin Y T, Liu Z G, Deng B P.Inter-annual variation in pelagic fish egg, larval, and juvenile assemblages during summer in the Yangtze River Estuary, China.Acta Ecologica Sinica,2015,35(21):7190- 7197.
