西藏草地植物功能性状与多项生态系统服务关系
2015-01-20余成群土艳丽罗黎鸣苗彦军武俊喜
潘 影, 余成群,2, 土艳丽, 孙 维,2, 罗黎鸣, 苗彦军, 武俊喜,2,*
1 中国科学院地理科学与资源研究所, 北京 100101 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 3 西藏自治区高原生物研究所, 拉萨 850001 4 西藏大学农牧学院, 林芝 860000
西藏草地植物功能性状与多项生态系统服务关系
潘 影1, 余成群1,2, 土艳丽3, 孙 维1,2, 罗黎鸣2,4, 苗彦军4, 武俊喜1,2,*
1 中国科学院地理科学与资源研究所, 北京 100101 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 3 西藏自治区高原生物研究所, 拉萨 850001 4 西藏大学农牧学院, 林芝 860000
针对植被功能性状与生态系统服务功能之间的相互关系,构建了西藏草地株高和可食性两种功能性状的9项指标,并基于土壤和植物采样,分析了9项植物功能性状指标和5项生态系统服务指标间的相关性,探讨了4种机制(Mass ratio, Selection, Niche complementarity及Insurance)在西藏草地的适用性。结果表明,9项功能性状指标中,株高Rao和可食种与所有种株高CWM比分别与土壤有机碳、土壤全氮和土壤含水率3项生态系统服务指标呈显著负相关及显著正相关。说明群落植被对光能竞争的互补性及可食性状植株在群落中的光能资源相对竞争力,与土壤固碳、肥力供给及水源涵养有显著相关关系。而群落可食种、优势种、优势种与次优势种对光能资源竞争力水平,可食植株多样性、可食植株在群落中的优势度及其光能资源竞争力均值,对草地生态系统服务无显著影响。西藏草地植物功能性状对多项生态系统服务的影响机制从光能资源竞争角度更符合Niche complementarity和Insurance理论,而从可食功能性状角度更符合Mass ratio和Selection理论。
西藏; 植物功能性状; 生态系统服务; 机理
植物与环境相互作用的漫长进化过程,形成了植物多样性;这些多样性改变了生态系统的过程与功能,影响着生态系统服务[1-4]。研究表明,生物多样性对生态系统功能与服务的影响多由植物性状的变化引起;这些植物性状也被称为植物功能性状[5-6]。植物功能性状对生态系统功能与服务影响的基础研究在土地利用、气候变化对生态系统功能与服务影响的模拟[7-9]以及生态规划及区域评价中都有所应用[10-13]。
国际上针对植物功能性状对生态系统功能与服务的影响提出了几种理论,包括Mass ratio, Selection, Niche complementarity及Insurance[6]。Mass ratio理论最早针对功能性状及多样性对生态系统初级生产力影响的研究[14],后扩展到生态系统功能[6]。Mass ratio理论认为在某个特定时间断面,生态系统功能主要由其优势种功能性状决定,而与次优势种和一些生物量较小物种无关[6,14]。Selection理论与Mass ratio大体一致,表示生态系统功能主要由群落中生物量最大的优势种的功能性状决定,与其他生物量较小物种无关[15]。
而Niche complementarity理论是指群落中不同生态位植物能够增强群落的资源利用率,从而提高群落生物量,影响生态系统功能[15]。而Insurance理论表示,在群落中,一些非优势种,甚至稀有种,能够在群落的生态系统功能供给方面提供“缓冲”作用,缓冲降雨、温度、病原体等生物、环境因子变化对群落生态功能的影响[16]。
4种理论在不同区域、不同生态系统有不同适用性,对这种适用性的研究有助于理解当地气候环境因子-植物群落-生态系统功能服务的相互联系和影响,指导当地生态系统恢复及生态系统功能与服务提升[17-19]。本文拟利用西藏草地土壤、植被采样数据,研究西藏草地植物典型功能性状对草地多功能的影响及四种理论的适用性。
1 研究地区和研究方法
1.1 研究区概况
新仓村(29°29′—30°02′N、91°15′—91°41′E)隶属于西藏自治区拉萨市达孜县,位于拉萨河流域的藏南谷地下游南岸。研究区域平均海拔4500 m,年均温7.5 ℃,年均日照时数3065 h,年均降水450 mm,降水多集中在植物生长季6—9月。采样区域土壤类型包括亚高山草甸草原土、亚高山灌丛草甸土、高山草甸土,植被类型包括亚高山草甸草原、亚高山灌丛草甸、高山草甸。
1.2 研究方法
本文选取株高及可食性两种功能性状,分析其功能性状差异性和功能性状均值[6]。在此基础上构建了9项指标(表1),其中群落株高功能性状差异性、可食性功能性状差异性参数反映植株功能性状在群落生态位的多样性,用以验证niche complementarity及insurance理论在西藏草地的适用性。所有种、优势及次优势种株高功能性状均值及可食性功能性状优势度用于分析优势种、次优势种等与多功能的相关性,用以验证mass ratio和selection理论在西藏草地的适用性。本文选取了地上可食生物量、土壤有机碳、土壤全氮、土壤含水率、植被覆盖度5项指标定量生态系统服务,研究不同功能性状指标与生态系统服务之间的关系。具体参数见表1。
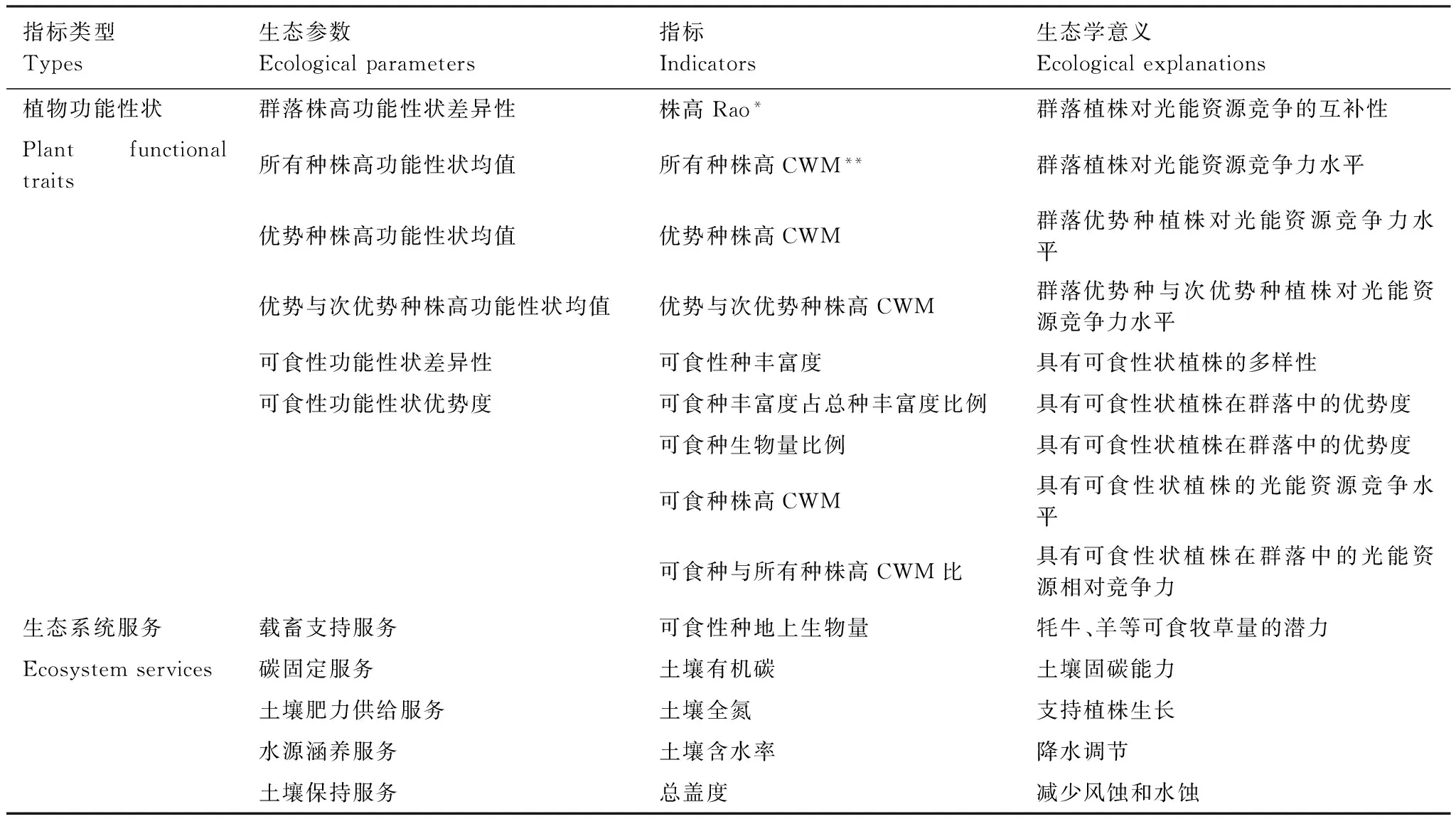
表1 植物功能性状对多项生态系统服务影响机制分析参数
功能性状差异性由Rao指数定量,计算公式如下:
(1)
式中,s为群落中物种数,pi、pj为第i、j个物种在群落种的相对盖度,dij为物种i和j之间某种性状的差额。
功能性状均值由CWM指数定量,计算公式如下:
(2)
式中,s为群落中物种数,pi为第i个物种在群落种的相对盖度,di为物种i功能性状值。
1.3 数据来源
本文数据样品于2012年7月采集于西藏拉萨市达孜县新仓村山坡草地,海拔梯度从3949 m至4940 m,沿坡底到坡顶共设15个5 m× 5m样地,每个样地中随机选取3个面积为0.5 m×0.5 m的样方重复。物种的株高、盖度和多度现场记录,地上生物量齐地面分种刈割,于实验室65 ℃烘干48 h称重[20-21]。并在刈割后的样方挖剖面环刀取土,测定含水量,土钻取土测定有机碳、全氮。本文的植株可食性是针对牦牛、山羊、绵羊及马,假设一种或一种以上牲畜采食该种草,则该草可食。为了减少牲畜啃食对植株高度的影响,在测量植株高度时,仅选取叶片完整的植株进行测量。
2 研究结果
2.1 西藏草地植物功能性状的相互联系
在株高与可食性两个功能性状的9项指标中,有6项指数相互呈显著相关(图1)。其中所有种、优势种、优势种与次优势种株高CWM指标两两之间呈显著正相关;说明某群落优势种和次优势种对光能资源竞争力越强,整个群落光能资源竞争水平越高。

图1 草地植物功能性状指数皮尔逊相关性Fig.1 Pearson correlation of the pairs of indicators of the plant functional traits点图为散点图,数字为皮尔逊相关系数,其中**代表在0.01水平显著相关
株高Rao指数和可食种与所有种株高CWM比指标呈显著负相关;说明可食性物种在群落中对光能资源竞争力越高,群落植物对光能资源竞争互补性越低,或者说群落株高功能性状趋同。
可食性种CWM分别与所有种、优势种、优势种与次优势种株高CWM呈显著正相关关系;主要由于新仓村草地群落大部分由可食性种建群。
可食性种的丰富度与株高相关指标无显著相关关系,说明群落可食性功能性状的差异性与群落植株对光能资源竞争力无明显关系。
2.2 西藏草地植物功能性状与多项生态系统服务的相互影响关系
9项植物功能性状指标中,株高Rao、可食种与所有种株高CWM比两项指标与土壤有机碳、土壤全氮和土壤含水率3项生态系统服务指标显著相关(图2,表2)。其中株高Rao与土壤有机碳、土壤全氮、土壤含水率呈显著负相关,说明群落植被对光能竞争互补性越强,则对土壤中养分、水分等资源有更多利用,导致土壤有机碳、全氮和含水率的降低。

图2 植物功能性状指标与生态系统服务指标散点图Fig.2 Scatter diagrams of the plant functional traits and ecosystem services
可食种与所有种株高CWM比和土壤有机碳、土壤全氮、土壤含水率呈显著正相关,说明具有可食性状植株在群落中的光能资源相对竞争力,与土壤有机碳、全氮和含水量的保持有显著正相关关系。
所有种、优势种、优势种与次优势种、可食种株高CWM,可食种丰富度、可食种占总种比例、可食生物量比例7项指标与生态系统服务的5项指标皆无显著相关关系。说明群落植株包括可食种、优势种、优势种与次优势种对光能资源竞争力水平,具有可食性状植株的多样性、具有可食性状植株在群落中的优势度和其光能资源竞争力均值,对草地生态系统服务无显著影响。
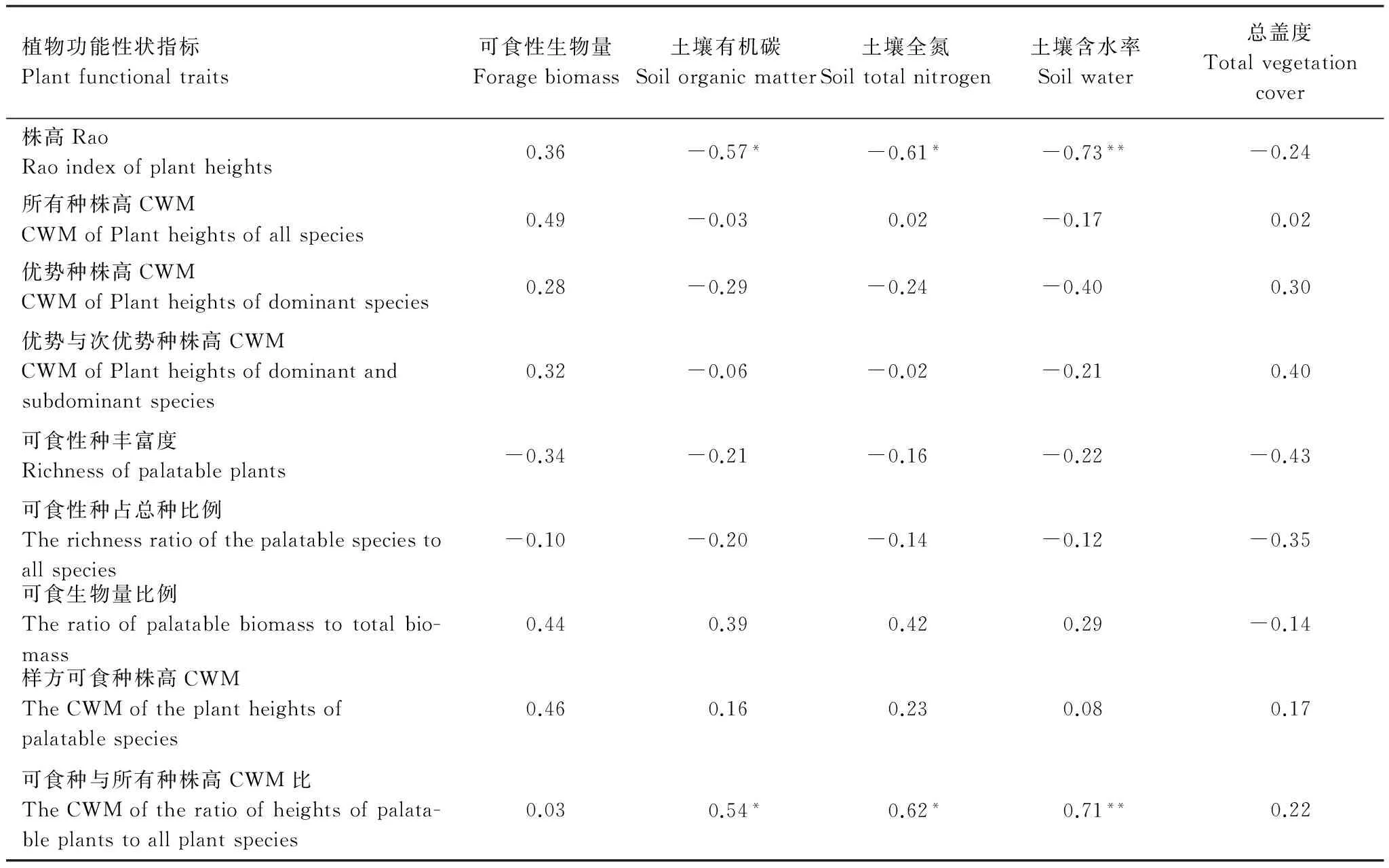
表2 植物功能性状指标与生态系统服务指标皮尔逊相关性
3 讨论
3.1 植物功能性状指标和生态系统服务指标选取
本文选取了株高和可食性两项植物功能性状,株高是反映植株对光能资源的竞争力较成熟的指标[6-7,12-13],同样比叶面积和叶片氮含量也能反映植株光能资源竞争力[5,7]。由于研究区域和问题的不同,可食性性状在很多国际研究中被忽视[5,22-23],但由于西藏当地牧民的生产生活和其生态安全屏障功能同样重要[24],可食性功能性状在西藏草地研究中尤为值得关注。除了株高和可食性两种功能性状外,还有很多功能性状,如叶片磷含量、叶片光合速率、植株生殖高度、根密度、凋落物碳氮含量等,可以反映群落的健康水平、繁殖能力、碳氮等营养物质循环[5,25-26]。这些功能性状都与生态系统服务相关[27]。生态系统水源涵养功能与水循环过程有关,其中地表植被、枯落物层及土壤的最大持水力、土壤的降雨入渗率、地表蒸散率等都是重要参数。草地土壤持水占草地生态系统持水的大部分,而小尺度上,本研究选取的土壤含水率可以反映一定时间断面水循环的状态,从一定程度上区别不同生态系统水源涵养功能。
3.2 西藏草地植物功能性状对多项生态系统服务的影响机制理论
Mass ratio, Selection理论和Niche complementarity及Insurance理论的较大分歧点是在群落中的一些非优势种和稀有种对生态系统功能是否有影响。
本研究结果表明,株高功能性状差异性对多项生态系统服务有显著影响,而无论是优势种、次优势种或全部种的株高性状均值对生态系统服务皆无显著影响。这意味着,西藏草地生态系统服务与植株株高差异程度有关,而与优势种或次优势种等的株高功能性状相对水平无关。本研究结果表明,西藏草地从光能资源竞争角度,其植物功能性状对多项生态系统服务的影响更加符合Niche complementarity和Insurance理论。
而西藏草地可食性功能性状对多项生态系统服务的影响机制与株高性状不同。可食性功能性状的指标更多程度上反映的是群落中可食种与不可食种的种间竞争和对环境的适应性。本研究结果表明,在反映可食性功能性状优势度与生态系统服务联系方面,可食种与所有种株高CWM较丰富度比例、生物量比例更加适合本研究。可食性种的优势度,或更进一步描述,是可食性种在群落中对光能的竞争力与草地生态系统服务显著相关。而可食性种间的多样性或株高差异性对生态系统服务并无显著影响。西藏草地可食功能性状对多项生态系统服务的影响更加符合Mass ratio和Selection理论。
4 结论
本文利用株高、可食性两个植物功能性状的9项指标与5项生态系统服务指标的统计分析,能够较好反映西藏草地植物功能性状与多项生态系统服务的相关性;同样也初步印证了Mass ratio, Selection, Niche complementarity及Insurance理论在西藏草地生态系统的适用性。对于可食性功能性状,前两种理论更加适用,而对于株高功能性状后两种理论更加适用。结果表明,西藏植株光能竞争的生态位差异及可食性种优势度与草地生态系统服务供给显著相关,而群落优势种、次优势种等的光能竞争水平及可食性种多样性与草地生态系统服务供给无关。
[1] Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning:current knowledge and future challenges. Science, 2001, 294(5543):804-808.
[2] Balvanera P, Pfisterer A B, Buchmann N, He J S, Nakashizuka T, Raffaelli D, Schmid B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters, 2006, 9(10):1146-1156.
[3] Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature, 2007, 448(7150):188-190.
[4] Mace G M, Norris K, Fitter A H. Biodiversity and ecosystem services:a multilayered relationship. Trends in Ecology and Evolution, 2012, 27(1):19-26.
[5] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985):821-827.
[6] Diaz S, Lavorel S, de Bello F, Quetier F, Grigulis K, Robson T M. Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(52):20684-20689.
[7] Quetier F, Thebault A, Lavorel S. Plant traits in a state and transition framework as markers of ecosystem response to land-use change. Ecological Monographs, 2007, 77(1):33-52.
[8] Soudzilovskaia N A, Elumeeva T G, Onipchenko V G, Shidakov I I, Salpagarova F S, Khubiev A B, Tekeev D K, Cornelissen J H C. Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(45):18180-18184.
[9] Geijzendorffer I R, Roche P K. Can biodiversity monitoring schemes provide indicators for ecosystem services? Ecological Indicators, 2013, 33:148-157.
[10] Naidoo R, Balmford A, Costanza R, Fisher B, Green R E, Lehner B, Malcolm T R, Ricketts T H. Global mapping of ecosystem services and conservation priorities. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(28):9495-9500.
[11] Anderson B J, Armsworth P R, Eigenbrod F, Thomas C D, Gillings S, Heinemeyer A, Roy D B, Gaston K J. Spatial covariance between biodiversity and other ecosystem service priorities. Journal of Applied Ecology, 2009, 46(4):888-896.
[12] Lavorel S, Grigulis K, Lamarque P, Colace M P, Garden D, Girel J, Pellet G, Douzet R. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. Journal of Ecology, 2010, 99(1):135-147.
[13] Lavorel S, Grigulis K. How fundamental plant functional trait relationships scale-up to trade-offs and synergies in ecosystem services. Journal of Ecology, 2012, 100(1):128-140.
[14] Grime J P. Benefits of plant diversity to ecosystems:immediate, filter and founder effects. Journal of Ecology, 1998, 86(6):902-910.
[15] Cardinale B J, Wright J P, Cadotte M W, Carroll I T, Hector A, Srivastava D S, Loreau M, Weis J J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(46):18123-18128.
[16] Diaz S, Fargione J, Chapin F S III, Tilman D. Biodiversity loss threatens human well-being. PLoS Biology, 2006, 4(8):e277.
[17] 王平, 盛连喜, 燕红, 周道玮, 宋彦涛. 植物功能性状与湿地生态系统土壤碳汇功能. 生态学报, 2010, 30(24):6990-7000.
[18] 龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011, 31(20):6088-6097.
[19] 胡耀升, 么旭阳, 刘艳红. 长白山不同演替阶段森林植物功能性状及其与地形因子间的关系. 生态学报, 2014, 34(20): 5915-5924
[20] Cornelissen J H C, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4):335-380.
[21] Perez-Harguindeguy N, Diaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quetier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, van der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3):167-234.
[22] Grigulis K, Lavorel S, Krainer U, Legay N, Baxendale C, Dumont M, Kastl E, Arnoldi C, Bardgett R D, Poly F, Pommier T, Schloter M, Tappeiner U, Bahn M, Clément J C. Relative contributions of plant traits and soil microbial properties to mountain grassland ecosystem services. Journal of Ecology, 2013, 101(1):47-57.
[23] Maestre F T, Quero J L, Gotelli, N J, Escudero A, Ochoa V, Delgado-Baquerizo M, García-Gómez M, Bowker M A, Soliveres S, Escolar C, García-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A., Gaitán J, Gatica M G, Ghiloufi W, Gómez-González S, Gutiérrez J R, Hernández R M, Huang X, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramírez-Collantes D A, Romão R, Tighe M, Torres-Díaz C, Val J, Veiga J P, Wang D, Zaady E. Plant species richness and ecosystem multifunctionality in Global Drylands. Science, 2012, 335(6065):214-218.
[24] Yu C, Zhang Y, Claus H, Zeng R, Zhang X, Wang J. Ecological and environmental issues faced by a developing Tibet. Environmental Science and Technology, 2012, 46(4):1979-1980.
[25] Jiang J, Li Y, Wang M, Zhou C, Cao G, Shi P, Song M. Litter species traits, but not richness, contribute to carbon and nitrogen dynamics in an alpine meadow on the Tibetan Plateau. Plant and Soil, 2013, 373(1/2):931-941.
[26] Butterfield B J, Suding K N. Single-trait functional indices outperform multi-trait indices in linking environmental gradients and ecosystem services in a complex landscape. Journal of Ecology, 2013, 101(1):9-17.
[27] 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1):150-165.
The relationship between plant functional traits and multiple ecosystem services in a Tibetan grassland ecosystem
PAN Ying1, YU Chengqun1,2, TU Yanli3, SUN Wei1,2, LUO Limimg2,4, MIAO Yanjun4, WU Junxi1,2,*
1InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,China2KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences.Beijing100101,China3TibetPlateauInstituteofBiology,Lhasa850001,China4AgriculturalandAnimalHusbandryCollegeofTibetUniversity,Linzhi860000,China
We studied the relationships among plant functional traits and multiple ecosystem services. Nine indicators of plant functional traits of plant heights and palatability were established, which were Rao index of plant heights, the community weighted mean value (CWM) of plant heights of all species, the CWM of plant heights of dominant species, the CWM of plant heights of dominant and subdominant species, the richness of palatable plants, the richness ratio of palatable species to all species, the ratio of palatable biomass to total biomass, the CWM of plant heights of palatable species, and the CWM of the ratio of heights of palatable plants to all plant species. The ecosystem services we analyzed included forage supply, soil carbon stocks, soil fertilization supply, water regulation, and soil retention. Further, we attempted to test four candidate mechanisms by which plant functional traits influence ecosystem services. These were:1) mass ratio, 2) selection, 3) niche complementarity, and 4) insurance. In 2012, we collected soil and vegetation samples from Xincang village, Lhasa, Tibet. Ecosystem services and plant functional traits were quantified based on these samplings. Pearson correlations were calculated among the nine functional traits, as well as between the functional traits and ecosystem services. The results showed that among the functional traits, the Rao index of plant heights was significant negatively correlated with soil organic carbon, soil total nitrogen, and soil water content. The ratio of the CWM of the heights of palatable plants to all plant species was significant positively correlated with soil organic carbon, soil total nitrogen, and soil water content. However, the CWM of the plant heights of all species, palatable species, dominant species, dominant and subdominant species, as well as the richness of palatable species, the ratio of the palatable specie richness to all species, and the ratio of palatable biomass to all species, were not significantly correlated with any of the ecosystem services. Our results demonstrated that the niche complementarity of the light captures of the plant community has negative impacts on ecosystem soil carbon stock, fertilization provisioning, and water regulating services. However, the relative niche competitiveness of light captures of palatable plants in the community has positive impacts on ecosystem soil carbon stock, fertilization provisioning, and water regulating services. The results imply, from the perspective of resource competitiveness, the concepts of niche complementarity and insurance best characterize the mechanisms by which plant functional traits determine ecosystem services in grassland. However, from the perspective of palatability, the concepts of mass ratio and selection are more reasonable.
Tibet; plant functional traits; ecosystem services; mechanisms
国家自然科学青年基金(31200364); 国家科技支撑计划项目(2011BAD17B05); “西部之光”人才培养计划项目(20100309); 西藏生态专项(Z2012C07G03)
2014-04-16; < class="emphasis_bold">网络出版日期:
日期:2014-12-18
10.5846/stxb201404160738
*通讯作者Corresponding author.E-mail: wujx@igsnrr.ac.cn
潘影, 余成群, 土艳丽, 孙维, 罗黎鸣, 苗彦军, 武俊喜.西藏草地植物功能性状与多项生态系统服务关系.生态学报,2015,35(20):6821-6828.
Pan Y, Yu C Q, Tu Y L, Sun W, Luo L M, Miao Y J, Wu J X.The relationship between plant functional traits and multiple ecosystem services in a Tibetan grassland ecosystem.Acta Ecologica Sinica,2015,35(20):6821-6828.
