油松不同种群2年生苗木光合特性的比较*
2015-01-17周丽杨学娇陈诗徐杨蔡年辉许玉兰段安安李悦
周丽,杨学娇,陈诗,徐杨,蔡年辉,许玉兰,段安安,李悦
(1.西南林业大学西南山地森林保育与利用省部共建教育部重点实验室,云南 昆明650224;
2.云南省保山市森林资源管理总站,云南 保山678000;3.北京林业大学生物科学与技术学院,北京100083)
油松(Pinus tabulaeformis)是松科(Pinaceae)松属(Pinus)的常绿乔木。生于海拔100~2 700 m地带,多组成纯林。极喜光,耐贫瘠,适于干冷气候,不耐水涝及盐碱[1]。在砂地、微酸性土壤、中性土壤及钙质黄土上均可生长,根系发达,生长迅速,材质优良,具有良好的生态、社会和经济效益,为西北地区荒山造林的先锋树种[2]。油松自然分布区范围广,分布于辽宁、吉林、内蒙古、山西、陕西、四川北部等14个省 (市、区)。随着分布区内地理变化,不同种群的油松在生理、生态、抗性等方面有着显著的差异,形成了不同种源和地理小种,其生长环境和水分生态系统日渐恶化[3~5]。近年来,关于油松和云南松 (P.yunnanensis)参与高山松 (P.densata)的杂交起源研究报道较多[6~11],结果表明不同群体杂交起源的多样性,油松不同种群在参与高山松杂交起源中遗传组成不一样[6~7,10]。光合作用、呼吸作用和蒸腾作用是绿色植物生存和生长的重要生理过程,其中光合作用又是植物体内最重要的化学反应[12~13],是对环境条件变化比较敏感的生理过程,是植物环境适应性研究的重要方面。不同地理种源油松移栽到与其分布区具有不同光照强度的云南松分布中心区后,其对光照的适应及光合生理特性等尚未见相关报道。本研究在前期研究工作的基础上,以油松分布区范围内的8个代表性种群为对象,研究其在云南松分布中心区生境下苗期的光合生理指标变化及油松种群间光合特性的变异规律,以揭示油松不同种群在云南松中心分布区生境下生长的生理响应及其差异,为其在云南松分布区引种提供科学依据。
1 材料与方法
1.1 试验地自然概况
试验地设在西南林业大学苗圃,位于东经102°45'41″、北纬 25°04'00″,海拔 1 945 m,属北亚热带半湿润高原季风气候,年平均气温15℃左右,年降水量700~1 100 mm,全年明显地分为干、湿两季,5-10月为雨季,降水量占全年的85%左右;11月至次年4月为旱季,降水量仅占全年的15%左右,年平均相对湿度68.2%,全年无霜期近年均在240天以上,全年晴天较多,日照数年均2 445.6 h,日照率56%。终年太阳投射角度大,年均总辐射量达129.78 kcal/cm2。土壤为红壤土,肥力中等。
1.2 试验材料与试验设计
参试的8个油松种群,均为实生苗,油松种子产地情况见表1。苗床设置5个重复,以完全随机区组试验排列。每个苗床平均分成24个小区,每个小区为1个种群并且平均分为3行,每行播20粒共60粒种子,于2011年4月17日播种,并进行正常的苗期管理,于发芽后,对每个种群随机选取5株进行标记,并且每月固定时间测量生长量。本次试验以8个2年生油松种群苗木为研究对象,于2012年10月中旬对其当年生针叶进行各项指标的测定。

表1 油松不同种群的地理分布Tab.1 Geographical distribution of different populations of Pinus tabulaeformis
1.3 测定方法
1.3.1 光合作用参数的测定
选取试验小区3个苗床,于3个苗床中部各选择植株和针叶长势中等,具有代表性的1株共3株进行挂牌和测定。试验选择10月中旬晴朗无云天气进行,在自然条件下,采用美国LI-COR公司生产的Li-6400光合测定系统,测定当年生中部成熟健康的功能叶片的净光合速率〔Pn,μmol/(m2·s)〕、胞间 CO2浓度 (Ci,μmol/mol)、蒸腾速率 〔Tr,mmol/(m2·s)〕、气孔导度〔Gs,mmol/(m2·s)〕,光合有效辐射〔PAR,μmol/(m2·s)〕、样品室CO2浓度 (CO2S,μmol/mol)、样品室相对湿度(RH_S)、水汽压亏缺 (VpdL)等光合参数。并根据这些光合参数计算下面几个指标,水分利用率(WUE,mmol/mol)=Pn/Tr;光能利用率 (LUE,gC/MJ)=Pn/PAR;羧化效率〔CE,mol/(m2·s)〕=Pn/Ci;气孔限制值 (LS)=1-Ci/CO2S。
1.3.2 光梯度曲线的测定
选择标记的植株,利用6400-02B LED红蓝光源,使用 Li-6400自动“light-curve”曲线测定功能,测定8个油松种群在梯度下Pn的变化情况,在测定过程中,光合测定仪的样品室流速均设定为300 μmol/s,CO2浓度设定为 400 μmol/mol,叶室温度(Ti)为20℃,设定的光合有效辐射通量密度(PPFD)梯度为0 μmol/(m2·s)、20 μmol/(m2·s)、50 μmol/(m2·s)、100 μmol/(m2·s)、150 μmol/(m2·s)、200 μmol/(m2·s)、400 μmol/(m2·s)、600 μmol/(m2·s)、800 μmol/(m2·s)、1 000 μmol/(m2·s)、1 200 μmol/(m2· s)、1 400 μmol/(m2· s)、1 600 μmol/(m2· s)、1 800 μmol/(m2·s)、2 000 μmol/(m2·s),共15 个水平。
以光合有效辐射通量密度PPFD≤200 μmol/(m2·s)时的Pn进行PPFD-Pn线性拟合,可以得出PPFD与Pn符合线性回归,方程拟合效果较好,R2均在0.949 1以上。直线回归方程为Pn=a×PPFD-b,其中:a,b均为常数,且a>0,b>0,根据该方程设Pn=0时,可以求得函数图像与横轴的交点的 PPFD值即为光补偿点〔LCP,μmol/(m2·s)〕,此外,还可以得出表观量子效率 (Q)、暗呼吸速率 (Rd)等光合作用参数[14]。

根据非直角双曲线[15]进行光响应曲线的拟合(图1),其模型的理论公式为:Pn=〔Q×PAR+,式中:Pn为净光合速率;Q为表观量子效率;PAR为光合有效辐射〔μmol/(m2·s)〕;P max为最大净光合速率〔μmol/(m2·s)〕;k为光响应曲线曲角;Rd为暗呼吸速率〔μmol/(m2·s)〕。
1.3.3 数据处理
完成数据收集后,用Excel 2003和SPSS 17.0进行分析处理。
2 结果与分析
2.1 油松光响应曲线的拟合
油松光响应曲线的拟合结果见图1。

图1 油松不同种群的光响应曲线Fig.1 Light response curve of different populations of Pinus tabulaeformis
图1曲线拟合结果表明,8个油松种群光响应曲线拟合的决定系数R2除土默特 (R2=0.951)、灵空山 (R2=0.969)以外,其他的都在0.978以上,拟合效果较好。由图1来看,8个油松种群光响应曲线规律基本一致,光照强度在0~500 μmol/(m2·s)之间,Pn迅速上升,光合速率随光照强度的升高而加快;当光照强度在500~1 500 μmol/(m2·s)之间时,Pn上升的速率减慢;PAR>1 500 μmol/(m2·s)时,Pn逐步稳定。但各种群Pn上升的速率和上升的趋势不同,呼市土默特 (图1-a)和青海互助 (图1-b)的光响应曲线在PAR>500 μmol/(m2·s)后,明显趋于平缓,达到完全饱和状态;而山西灵空山 (图1-d)的光响应曲线在PAR为0~2 000 μmol/(m2·s)时,并没有达到完全饱和,一直处于上升趋势,说明山西灵空山的油松具有较高的耐高强辐射能力。在相同的光照强度下,北京松山 (图1-f)油松种群的Pn明显高于其余7个种群。
由表2可知,油松不同种群的Pn在8.293~23.338 μmol/(m2·s)之间波动,其中,最大的是北京松山种群,最小的是呼市土默特种群。而陕西宁陕种群具有最小的 LCP〔3.15 μmol/(m2·s)〕,具有较强的利用弱光的能力,四川九寨沟种群次之〔12.292 μmol/(m2·s)〕,而山西方山油松种群对弱光的利用能力最低,LCP 为37.69 μmol/(m2·s)。Pn没有一个明显的饱和点,从不饱和到饱和是一个渐变的过程,因此对于光饱和点(LSP,μmol/(m2·s)不能给出一个确定的值,只能给出一个大致的范围[16]。光响应曲线估计值表明,北京松山种群具有最大的利用强光的能力,其LSP>2 000 μmol/(m2·s),山西灵空山种群次之,呼市土默特和青海互助种群的LSP值最小,约为500 μmol/(m2·s),利用强光的能力最小。
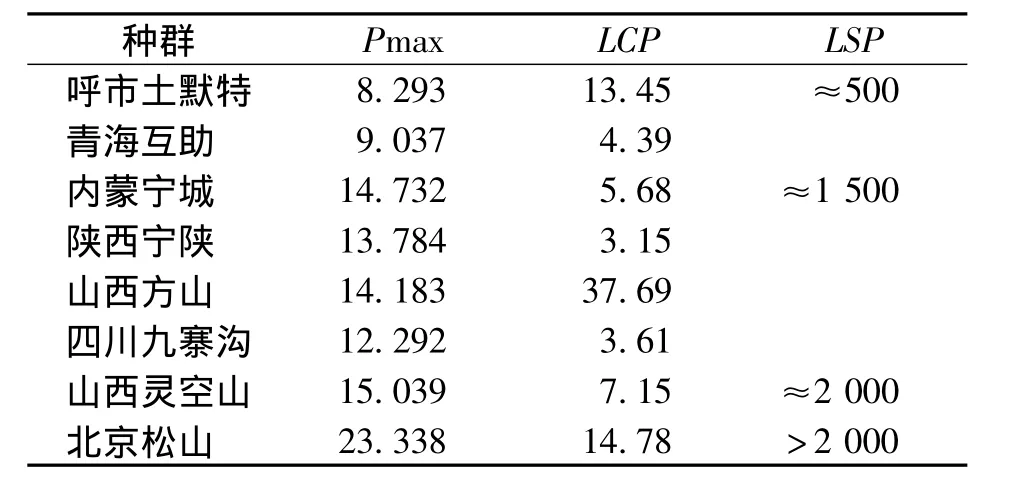
表2 油松不同种群光合响应曲线参数Tab.2 Photosynthetic response curve parameters of different populations of Pinus tabulaeformis μmol·m-2·s-1
2.2 光合生理指标比较
对8个油松种群的 Pn、Tr、Gs、Ci、WUE、LUE、CE、LS进行方差分析,结果见表3。
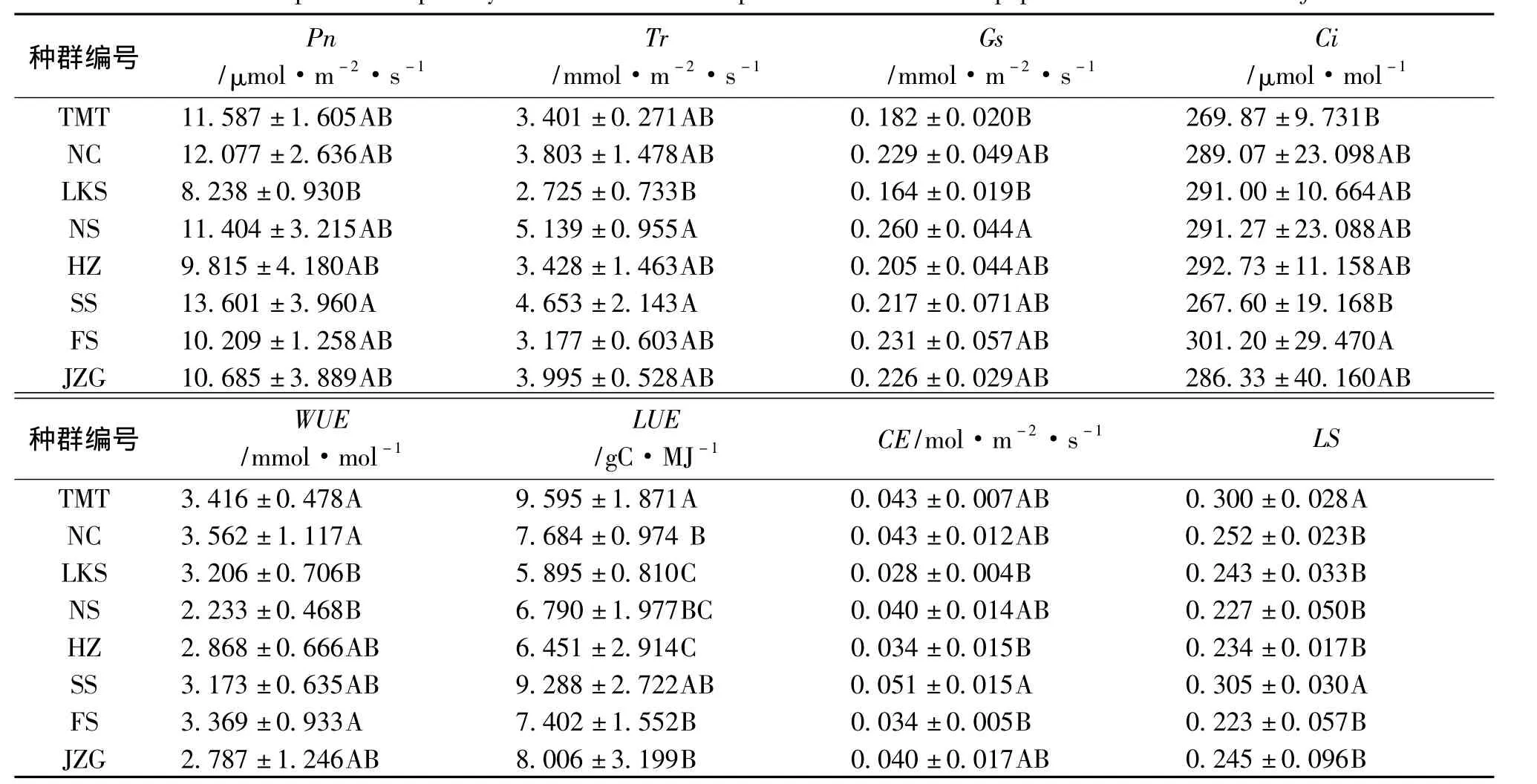
表3 油松不同种群的光合作用特征参数比较Tab.3 Comparison on photosynthesis characteristic parameters of different populations of Pinus tabulaeformis
油松不同种群之间光合特征指标参数Pn(p=0.000)、Tr(p=0.000)、Gs(p=0.000)、Ci(p=0.001)、WUE(p=0.001)、LUE(p=0.000)、CE(p=0.000)、LS(p=0.000)在 99% 置信度水平下差异都极显著,变异范围较广,除去Ci(CV=0.09),变异系数均≥0.22,变异最大的是Tr(CV=0.36),表明不同油松种群间的光合生理指标具有较大的变异。
对不同种群间的光合参数分析 (表3)显示,8个油松种群之间的各个光合参数之间存在着极显著的差异,从平均值可以看出,北京松山 (SS)种群的 Pn 〔13.601 ±3.960 μmol/(m2·s)〕、CE〔0.051 ± 0.015 mol/(m2·s)〕和 LS(0.305 ±0.030)均最大,其次为采自内蒙宁城 (NC)的种群 〔Pn=12.077 ± 2.636 μmol/(m2·s)、CE=0.043 ± 0.012 mol/(m2· s)、LS=0.252 ±0.023〕,该种群具有最大的 WUE(3.562 ± 1.117 mmol/mol);而山西灵空山(LKS)油松种群,其Pn、Tr、Gs、LUE 分别为 8.238 ±0.930 μmol/(m2·s)、2.725 ± 0.733 mmol/(m2· s)、0.164 ± 0.019 mmol/(m2·s)、5.895 ±0.810 gC/MJ,在8 个种群中均是最小的;陕西宁陕(NS)的油松种群则具有最大的 Tr〔5.139 ± 0.955 mmol/(m2·s)〕、Gs〔0.260 ± 0.044 mmol/(m2·s)〕和 最 小 的 WUE〔2.233 ±0.468 mmol/(m2·s)〕。Tr的波动范围最大,为2.725 ~5.139 mmol/(m2·s),Ci的波动范围最小,为 267.60 ~301.20 μmol/mol。从标准差可以看出,同一种群之间的个体在各参数之下也存在着显著的差异。波动最大的是Ci,为9.731~40.160 μmol/mol,Gs波动的范围最小,为 0.019 ~0.071 mmol/(m2·s)。
2.3 油松种群的各光合参数关系
植物的Pn、Tr除了受到外界环境因子的影响,还受到内在生理因子的调节。通过对8个种群各光合生理参数进行简单相关性分析 (表4)可知,大部分光合参数与Pn和Tr有着显著 (p<0.05)和极显著 (p<0.01)的相关关系。油松的Gs与Pn和Tr之间有着极显著的正相关关系 (r=0.574**和0.726**),PAR 与 Pn、Tr的相关系数分别为0.260**、0.404**;Pn与 LS间存在极显著正相关关系 (r=0.558**),与VpdL间存在显著正相关关系 (r=0.227*),与Ci和CO2S间存在极显著负相关关系,相关系数分别为 -0.572**、 -0.255**。VpdL、CO2S、RH_S均与Tr有着极显著的相关性,其中与CO2S和RH_S间呈负相关 (r=-0.315**和-0.472**)。Ci与 Ls存在极显著的负相关关系,相关系数为-0.926。
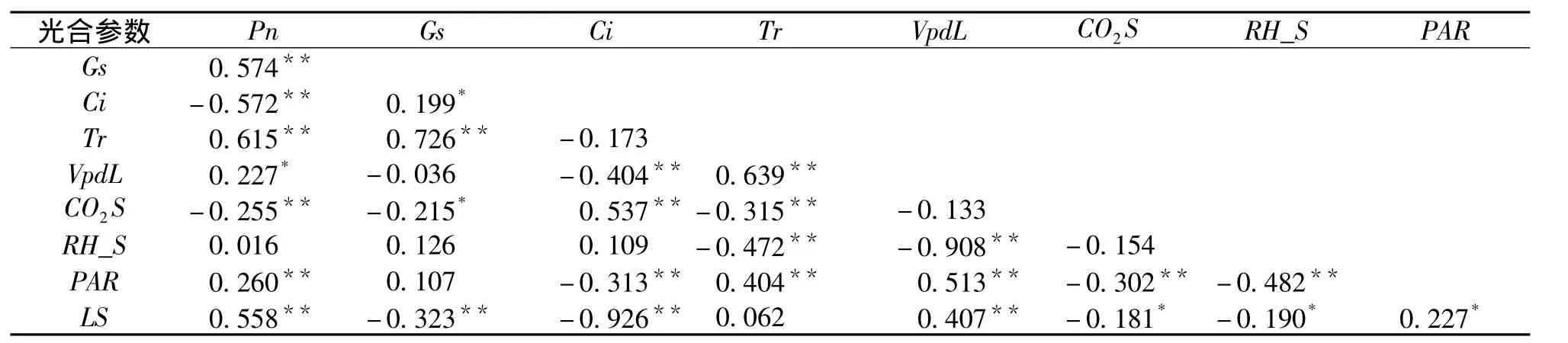
表4 光合参数之间的相关性分析Tab.4 Correlation analysis between photosynthetic parameters
3 结论与讨论
光响应曲线是植物Pn随着PAR的变化而改变的反应曲线,①式可以较好地模拟光合速率随光强的变化,通过光响应曲线可以直观地看出植物光合能力的强弱。植物的光合能力在一定程度上是由其遗传特性决定的,LSP、LCP的大小反映了植物对强光、弱光的利用能力,P max则反映了植物光合能力的强弱。LCP越小,表明其利用弱光的能力越强,高LCP低LSP的植物对光环境的适应较弱,反之则较强。由表2可知,呼市土默特种群具有较高的LCP,而LSP和P max最低,说明其对光照的适应性最弱;而北京松山种群则具有最大的LSP和P max,但其LCP并不是最小的,说明其对云南松分布区光照环境的适应能力较强。综合比较8个油松种群,本次试验结果与高琼等[17]的研究结果存在一定差异,可能是由于云南与北京的地理环境和生态环境存在较大差异引起,是油松适应云南松生境的结果。
衡量光合作用最重要的指标是Pn,植物进行光合作用,是一个积累有机物的重要过程,在相同的环境下,Pn的高低,可以说明植物能否很好地生存。Tr反映了植物在一定时间内单位面积蒸腾的水量,在同一环境里,Tr越高,植物的抗旱能力越弱,反之,植物抗旱能力越强。Gs反映了气孔张开的程度,影响着光合作用、呼吸作用和蒸腾作用,对蒸腾作用有着直接的影响[18]。WUE是植物光合作用、蒸腾作用的综合反映[19]。本研究结果表明,北京松山油松种群在试验生境下,Pn最大,光合能力最强,Ci最小,表明它能利用最低的CO2来获得最高的光合速率,但其Tr较高,仅次于陕西宁陕种群,表现出较弱的抗旱能力,WUE、LUE值较大,具有较强的水分和光能利用能力,表明其生理基础较强。山西灵空山油松种群,其Pn、Tr、Gs和LUE均最小,光能利用能力最弱,光合作用较弱,蒸腾作用最慢,但是其抗旱能力最强,其生理基础相对较弱,研究结果与高琼等[17]的研究结论一致。
相关分析表明,大部分光合参数与Pn和Tr有着显著 (p<0.05)和极显著 (p<0.01)的相关关系。其中Gs与Pn和Tr间的相关性极显著,可能是影响油松不同种群Pn、Tr的最大内部因子;PAR 与 Pn、Tr的相关系数分别为 0.260、0.404,也存在极显著的相关关系,推测PAR可能是影响Pn、Tr最大的外部环境因素,其结果与徐佳佳等[18]的研究结果一致。
不同种群油松苗木的光合特性除了受到遗传特性、生理特性的影响,还由其生长的地理位置、环境条件和气候条件决定。本次试验所得结论是针对2年生油松苗木,于生长当年生长基本停止时所测定的,在不同季节或不同年龄阶段的变异规律还有待进一步研究。
[1]张志翔.树木学2版[M].北京:中国林业出版社,2005.
[2]陈晓阳,沈熙环.林木育种学[M].北京:中国林业出版社,2007.
[3]Li Cui,Chai Baofeng,Wang Mengben.Population genetic structure of Pinus tabulaeformis in Shanxi Plateau,China[J].Russian Journal of Ecology,2008,39(1):34-40.
[4]郑淑霞,上官周平.黄土高原油松和刺槐叶片光合生理适应性比较[J].应用生态学报,2007,18(1):16-22.
[5]狄晓燕,池喜梅,陈建文,等.5个油松种源光合特性的比较研究[J].植物研究,2012,32(2):165-170.
[6]Song BH,Wang XQ,Wang XR,et al.Maternal lineages of Pinus densata,a diploid hybrid[J].Molecular Ecology,2002,11(6):1057-1063.
[7]Song BH,Wang XQ,Wang XR,et al.Cytoplasmic composition in Pinus densata and population establishment of the diploid hybrid pine[J].Molecular Ecology,2003,12(11):2995-3001.
[8]Mao JF,Wang XR.Distinct niche shift characterizes the homoploid hybrid speciation of Pinus densata on the Tibetan Plateau[J].The American Naturalist,2011,177(4):424-439.
[9]Mao JF,Li Y,Wang XR.Empirical assessment of the reproductive fitness components of the hybrid pine Pinus densata on the Tibetan Plateau[J].Evolutionary Ecology,2009,23(3):447-462.
[10]Wang BS,Mao JF,Gao J,et al.Colonization of the Tibetan Plateau by the homoploid hybrid pine Pinus densata[J].Molecular Ecology,2011,20(18):3796-3811.
[11]Xing FQ,Mao JF,Meng JX,et al.Needle morphological evidence of the homoploid hybrid origin of Pinus densata based on analysis of artificial hybrids and the putative parents,Pinus tabuliformis and Pinus yunnanensis[J].Ecology Evolution,2014,4(10):1890-1902.
[12]沈允钢,施教耐.动态光合作用[M].北京:科学出版社,1998.
[13]王建林,于贵瑞,房全孝,等.不同植物叶片水分利用效率对光合CO2的响应与模拟[J].生态学报,2008,28(2):525-533.
[14]梁军生,陈晓鸣,杨子祥,等.云南松与华山松人工混交林针叶光合速率对光及CO2浓度的响应特征[J].林业科学研究,2009,22(1):21-25.
[15]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].农业基础科学,2005,21(8):76-79.
[16]钱莲文,张新时,杨智杰,等.几种光合作用光响应典型模型的比较研究[J].武汉植物学研究,2009,27(2):197-203.
[17]高琼,王维有,梁冬,等.8个种源油松生长和光合生理指标的比较研究[J].北京林业大学学报,2014,36(2):87-93.
[18]徐佳佳,张建军,王清玉,等.油松和侧柏的光合蒸腾特性及其与环境因子的关系[J].东北林业大学学报,2011,39(7):15-18.
[19]张卫强,贺康宁,王正宁,等.光辐射强度对侧柏油松幼苗光合特性与水分利用效率的影响[J].中国水土保持科学,2006,4(2):108-113.
