黑河流域中游水陆交错带湿地土壤盐度对芦苇种群生长发育影响研究
2015-01-13李雯靖孟好军王立赵维俊牛赟雷军杨逍虎
李雯靖,孟好军,王立,赵维俊,牛赟,雷军,杨逍虎
(1.甘肃农业大学林学院,甘肃 兰州730070;2.甘肃省祁连山水源涵养林研究院,甘肃 张掖734000)
生态交错带是一个基本的景观单位[1],是生态系统管理的重要基础[2,3]。早在20世纪70年代,Clements就提出,生态交错带是指由气候决定的植物群丛交叠的应力区,主要包括3个类型:边缘(Local edges or margins)、树线(Treeline)和群落交错带(Biome ecotone)[4-6],黑河流域水陆交错带是生态交错带的景观带之一,分布的典型植物芦苇种群是景观带的重要标志。芦苇是我国分布最广、产量最大的禾本科植物之一[7],是适应性广,抗逆性强,种质资源十分丰富的世界性植物[8],也是黑河流域中游水陆交错带湿地区内挺水植物的优势种,因受水分与土壤条件的影响,在群落组成与分布上,呈现出随水环境规律性的分布特征。虽然水环境是影响植被生长的重要因素,但土壤条件也决定着植被生长繁殖。繁殖是植物繁衍后代、延续种族的最根本行为和过程,它不仅是植物种群形成、发展和进化的核心问题之一,也是植物群落和生态系统演替的基础[9]。盐度是影响湿地植物生存、分布、生长和繁殖的重要环境因子[10]。芦苇作为广生态幅的物种,喜生沼泽,无性繁殖力很强,天然种群以根茎繁殖补充更新,形成了单优群落或双优群落[11-13]。目前,黑河中游水陆交错带湿地生态系统植物生长环境面临过度开发、面积缩小、污染严重、水源涵养功能降低等诸多生态环境问题。本研究以交错带湿地区内挺水植物的优势种——芦苇种群为研究对象,研究其土壤盐度对芦苇种群生长发育的影响,以期为黑河流域湿地生态系统的恢复提供数据,为该流域湿地生态环境评价、生态环境修复和生态科学研究等提供理论依据。
1 研究区概况
研究区地理位置介于100°06′—100°54′E,38°32′—39°24′N,区内地势由南向北倾斜,农田、沼泽、内陆盐沼镶嵌分布,地下水位较高。气候属明显的大陆性荒漠气候,其显著特点是:降水稀少而集中,年降雨量仅129mm,在时间分布上,多集中在6—9月,约占全年总量的71.9%,春季降水仅占14%,年内降水分布很不均匀,年际变化较大;蒸发强烈,全区年平均蒸发量2 047mm,干旱指数高达10.3,大气干燥度为4.9;日照充足,温差大,太阳年辐射总量619.6kJ·m-2,年日照时数为3 085h;多年平均气温为7℃,历年最高气温为37.4℃,最低气温为-28℃;≥0℃积温3 388℃,持续时间240 d,≥10℃积温2 896.6℃,持续时间160.2d,无霜期153d;全年盛行西北风,年均风速2m·s-1,最大风速36m·s-1,年均大风日数14.9d,最多天数40d,最少3d,年均沙尘暴日数20.3d,最多33d,最少14d。灾害性天气有大风、沙尘暴、干热风、干旱、霜冻、初春低温等。该湿地区现在有降水、地表水、地下水等多种形式的水资源,降水是最基本的水资源,制约着区域内的水分条件和湿地的类型。表水资源合计3 185万m3,主要来自城市生活污水、工业废水和泉水溢出。城区生活污水进入湿地区域内的排放量850万m3·a-1,工业废水排放量395万m3,泉水溢出量1 940万m3;现有灌溉机电井50眼,年提水量375.71万 m3。
2 研究方法
2.1 植物和土壤样品的采集
2014年生长季(5—9月),在研究区选取12个典型湿地芦苇群落作为固定监测点,然后在每个固定监测点选择面积大小为1m×1m(测定芦苇地上部用)和1m×0.5m(测定芦苇不定根和根状茎生物量)的小样方各2个,总共24个小样方。取样时间为5月25日、6月25日、7月25日、8月25日、9月25日和10月5日,每月取1次芦苇植物样和生长芦苇的土壤样。植物样采集芦苇不同部位样品(根状茎、茎秆、叶),置于80℃烘箱中,烘至恒质量称量。
土壤样与芦苇植物样品的采集同时进行,根据芦苇地下根系分布特点与土壤自身的分层特点,分别采集不同层次土壤(A1:0~10cm,A2:10~20 cm,A3:20~40cm,A4:40~60cm),取样时将同一监测点采集的2个小样方质量大致相当的土样混合,样品带回置于实验室内通风处阴干后装袋保存,用于土壤pH和含盐量的测定。土壤pH采用比色法测定,含盐量利用电导法测定。
3 结果与分析
3.1 地表淹水深度季节变化
湿地地表淹水深度的季节变化,势必影响植物生长的水体及土壤中盐度的含量变化。研究区域的地表水位变化较大(图1),在芦苇的生长期5—9月的表现为:5月,芦苇萌发以及生长的初期,地表水位很浅,平均深度为6.5cm,其主要原因是这一时期内该区域水位随着春季返潮的回落,地下水位下降,地表水随之下降;6月,由于气温的不断升高,蒸发量急剧增大,致使地表水位有所下降;7月,随着该区域雨季的来临,由于大气降水的补给,地表淹水深度之间增大;9月,上升至全年的最高点57.3cm,其原因一则是大气降水的补给,二则是地下水的掘取量大幅度减少和农闲用水的补给;随后由于天气逐渐变冷,大气降水的减少,到最后一次采样 (10月5日)时,地表水位淹埋深度又大幅度下降。
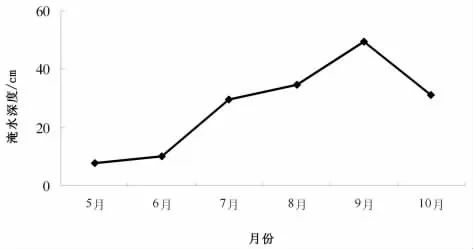
图1 芦苇种群地表淹水深度季节变化
3.2 盐分对芦苇生长的影响
盐分是影响芦苇生长的主要因素,要准确分析盐分对芦苇生长的影响,有必要探讨盐分对芦苇各个生长期的影响,以及盐分对芦苇各生长部分的影响,并进行综合分析,才能研究其影响程度的变化机理。其研究结果对芦苇的人工栽培和对多年根茎芦苇的管护具有重要的作用。根据监测数据计算盐分和对芦苇生长的影响,监测数据见表1。从表1可以看出,盐度对于芦苇的植株密度和植株高度有着较大的影响,随着芦苇种群生长基质土壤盐度的增加,其植株的密度和高度均呈不同程度的减小。
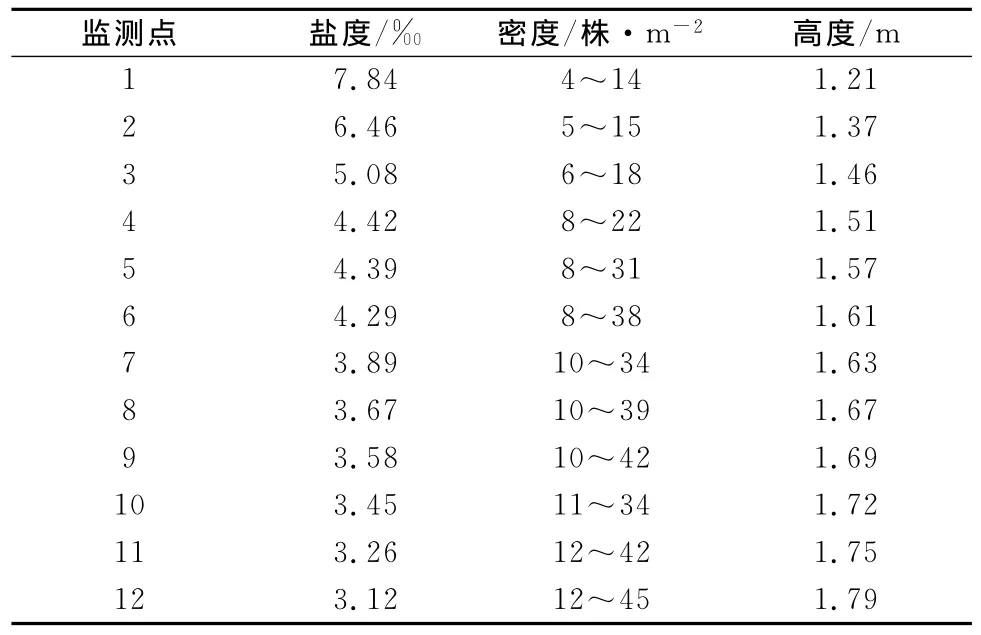
表1 各监测点芦苇生长与盐度监测数据
3.3 盐分对芦苇生物量的影响
在芦苇湿地生态系统中,芦苇生物量的变化,说明了其变化具有明显的季节性,也体现了芦苇对环境的极大适应和在此环境条件下的生长能力。土壤中盐分含量的高低对芦苇的生物量所产生的影响较为明显(图2)。从图2可以看出,随着盐度的增加,芦苇的地上生物量部分呈缓慢下降趋势,说明在该区域,较高盐度显著抑制了芦苇种群的生长,对抗盐胁迫能力具有一定的适应范围,芦苇种群对盐碱胁迫的响应机制有待进一步深入研究,特别是内陆河流域湿地的特殊性诸如水分转化频繁、降水少而蒸发大等对植物生长有着特殊的影响机理。
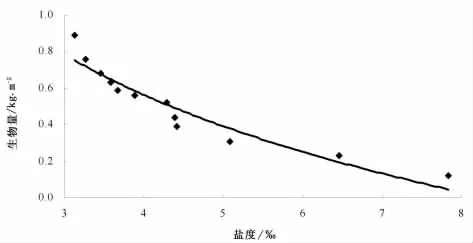
图2 土壤盐度与生物量的关系
3.4 盐度对芦苇根茎相对生长速率的影响
为了探讨土壤含盐量对芦苇生长情况的影响,我们以芦苇根茎生长速率和土壤含盐量为例,进行了相互作用的研究。由图3可知:芦苇根茎相对生长速率的大小受土壤盐分含量的影响十分突出,随着土壤中含盐量的增加芦苇根茎生长的相对速率呈下降趋势。

图3 芦苇根茎相对生长速率与盐度的关系
从不同生境中芦苇的群落组成的变化趋势不难看出芦苇具有相当大的抗盐能力,同时也看出土壤盐分对芦苇群落和个体生长发育也产生一定的抑制作用,说明它不是一种真正的盐生植物,真盐生植物的生长规律是低盐促进,高盐抑制,其生长发育应有一最佳适宜盐度。因此,可以认为芦苇是一种适应性较强的植物,从抗盐机理考虑可以认为它是一种假盐生植物(pseudohalophyte)[14]。在土壤中盐度低于15‰时,芦苇能够存活生长,但土壤中盐度高于15‰时,其生长将出现负增长。
3.5 盐度对芦苇叶片生长的影响
为了探讨盐度对叶片生长的影响,我们进行了叶片死亡与盐度关系的测试,试验结果表明,在土壤盐度达到15‰时,叶片大量死亡,由此确定在内陆河流域湿地芦苇叶片的致死盐度为15‰。从图4可以看出,在土壤盐度没有达到芦苇叶片的致死界限时,芦苇叶片的寿命跟盐度没有明显的直接相关的关系。芦苇的幼苗期与成熟期有很大的不同,它在任何的盐度下都有很高的死亡率。盐度在生长期对叶子的数量影响非常明显。

图4 芦苇叶片死亡与盐度的关系
4 结论与讨论
4.1 芦苇作为一种广布植物,在不同生境中产生了不同的生态类型[15],芦苇的生长与土壤盐度和水分有极其密切的相关性。随着盐度的增加,芦苇的地上生物量部分呈缓慢下降趋势;盐度对于芦苇的植株密度和植株高度也有较大的影响。在土壤中盐度低于15‰时,芦苇能够存活生长,但土壤中盐度高于15‰时,其生长将出现负增长。对于土壤盐分含量影响芦苇生长的机理有待进一步的研究。
4.2 在土壤盐度没有达到芦苇叶片的致死界限时,芦苇叶片的寿命跟盐度没有明显的直接相关关系。芦苇的幼苗期跟成熟期有很大的不同,它在任何的盐度下都有很高的死亡率。盐度在生长期对叶子的数量影响非常明显。但土壤盐度致死芦苇植株死亡的界限值有待进一步研究。
[1]Wilson J B,Agnew A D Q.Positive-feedback switches in plant communities[J].Advances in Ecological Research,1992,23(3):263-336
[2]Kupfer J A,Cairns D M.The suitability of montane ecotones as indicators of global climatic change[J].Prog Phys Geog,1996,20(3)253-272
[3]Allen C D,Breshears D D.Drought-induced shift of a forest-woodland ecotone:rapid landscape response to climate variation[J].Proc Natl Acad Sci USA,1998,95(25):14839-14842
[4]Risser P G.The status of the science examining ecotones[J].Bioscience,1995,45(5):318-325
[5]Laurance W F,Didham R K,Power M E.Ecological boundaries:a search for synthesis[J].Trends in Ecology & Evolution,2001,16(2):70-71
[6]Clements F E.Research methods in ecology[M].Lincoln Nebraska USA:University of Nebraska Publishing Company,1905:334
[7]吴庆定,易林,梁盛.响应面发优化芦苇杆粉末高压无胶成形工艺[J].中南林业科技大学学报,2012,32(1):144-149
[8]田莉,袁茂强,马庆勇,等.南方高秆芦苇收割装备的适应性设计与探讨[J].中南林业科技大学学报,2011,31(11):153-156
[9]张友民,卢曦,杨允菲.芦苇种群的繁殖方式对湿地生态环境的适应机制[J].吉林农业大学学报,2008,30(3):279-283
[10]仲崇庆.湿地植被与土壤环境因子关系研究进展[J].安徽农学通报,2009,15(13):170-172
[11]张友民,刘兴土,肖洪兴.三江平原芦苇湿地植物多样性的初步研究[J].吉林农业大学学报,2003,25(1):58-61
[12]张友民,刘兴土,曲同宝.三江平原芦苇营养器官的生态解剖学研究[J].吉林农业大学学报,2003,25(2):161-164
[13]中国湿地植被编辑委员会.中国湿地植被[M].北京:科学出版社,1999:59
[14]倪红伟,王宏韬,李君霞,等.三江平原小叶章种群生长速率的季节动态[J].国土与自然资源研究,1996(3):67-71
