不同株龄蒙古岩黄芪表型性状的变异特征
2015-01-02周媛媛周向睿周志宇金茜李金辉宋鑫
周媛媛,周向睿,周志宇,金茜,李金辉,宋鑫
(1.草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州730020;2.甘肃农业大学草业学院,甘肃 兰州730070)
毛乌素沙漠南缘地区属于多层次过渡带,生态环境脆弱,是沙丘移动的活跃地带。在干旱气候与不合理的人类活动下[1],成为我国沙漠化较为严重的地区之一。虽然国家早就认识到要对土壤沙漠化进行预防和治理,并采取行政手段大面积封育、飞播和人工造林等手段,经过遥感影像分析发现:在过去30年里,毛乌素沙漠及周边地区的沙漠化得到全面逆转[2],但2000-2005年期间,仅有2.33%的沙漠化土地发生了动态变化。目前,沙尘暴依旧影响着北方地区,并有南侵的趋势。因此防止沙漠化已成为一项长期而艰巨的工程。
榆林地区蒙古岩黄芪在沙地生长良好,可以有效固定流动沙丘,改善沙丘土壤状况[3-4],因此对其他地区沙漠化防治和治理有一定的借鉴和指导意义。蒙古岩黄芪(Hedysarummongolicum),又名踏郎、羊柴、沙欠、山花子等,是一种生长快、耐贫瘠、寒冷、干旱及沙埋等具有很强根蘖能力的多年生豆科灌木[5]。具体来说,蒙古岩黄芪的根状茎具有极强的营养繁殖能力,使它能在半固定、流动沙地上生存,并表现出极强的生命力[5]。因其幼苗生长速率大于土壤干沙层厚度的变化,故是目前飞播效果最好的树种之一。此外,它是良好的蜜源,可以改良土壤,又有较高的饲用价值,被喻为沙漠里的“红花苜蓿”。学者对蒙古岩黄芪的研究主要集中在生物学特性[6]、生长环境与群落特征[7]、抗逆性[8]、育苗造林、抚育管理及病虫害防治技术等方面。
变异是生物界基本特征之一,对形态变异的度量、表述和分析是研究进化问题的基础[9]。时间变异和空间变异是变异的两种类型,目前多数学者[10-11]研究的是植物形态随空间变化而产生的变异,梁坤伦等[12]对青藏高原的3个紫穗槐(Amorphafruticosa)居群的主要形态特征变异进行了分析,杨东华和赵雨森[4]研究了不同生境对踏郎(Hedysrummongolicum)生长及土壤养分空间差异的影响,研究发现:由于环境不同,不同居群内及居群间的形态特征存在一定程度的变异。为了更好、更全面地了解这种西部荒漠地区适宜发展的优良灌木,本文主要应用主成分分析法(principal component analysis,简称PCA)[13]从时间轴这个方向来研究蒙古岩黄芪的表型变异特征,通过本文的研究,可以了解蒙古岩黄芪生长的过程中表型性状变异的趋势及影响这些变异的主要因子,初步判定了这些变异与土壤环境之间的关系,为沙区种植蒙古岩黄芪和沙漠化可持续治理提供理论依据,为生物进化研究奠定理论基础。
1 材料与方法
1.1 研究区概况
试验区位于107°28′-111°15′E,36°57′-39°34′N,海拔980~1534m,绝对气温-27.0~37.6℃,平均气温7.9℃,无霜期150d,年平均降水438.4mm(主要集中在7-9月),属于半干旱内陆性季风气候的陕西省榆林市靖边县毛乌素沙漠南缘。地貌主要为流动、半固定或固定沙丘;土壤为各类风沙土;植被类群主要为沙生植物:樟子松(Pinussytvestrisvar.mongolica)、河北杨(Populushopeiensis)、花棒(Hedysarumscoparium)、紫穗槐、柠条(Caraganakorshinskii)、酸刺(acidthorn)、蒙古韭(Alliummongolicum)、苦豆子(Sophoraalopecuroides)、沙柳(Salixpsammophila)、沙蒿(Artemisiaordosica)、沙枣(Elaeagnusangustifolia)、沙打旺(Astragalusadsurgens)、沙蓬(Agriophyllumsquarrosum)等。
1.2 样品的采集
2012年8月在榆林市靖边县毛乌素沙漠南缘,选取1957年(55龄)、1984年(28龄)、2004年(8龄)种植的蒙古岩黄芪各3个样地(每个样地面积为100m×100m),在各样地内随机选取3株中等大小样株,选择茎干相对粗壮、冠幅大的植株(母株)作为实验材料,与由种子掉落而生长出来的或是根状茎产生的弱小幼龄新株相互区别,供采样分析。单株植物测定冠幅、自然高度、叶厚、叶长、叶宽(20片叶子)、主枝(茎干较粗较高)直径、主枝重量、一级侧枝长度、一级侧枝直径、一级侧枝重量、一级侧枝总长度、二级侧枝长度、二级侧枝直径、二级侧枝重量、二级侧枝总长度、主根直径、根重量等形态指标。其中叶长、叶宽、叶厚、主枝直径、一级侧枝直径、二级侧枝直径及根(基部)直径使用游标卡尺测定,枝序的测定采用离心法。
1.3 土样采集与理化分析
去除落叶层,用土壤刀从植株基部开始逐段、逐层挖去上层覆土,追踪根系的伸展方向,沿侧根找到须根部分,剪下分枝,然后轻轻抖动,落下的为非根际土,仍粘在根上的为根际土,收集保存,供分析用[14-15]。将野外带回的样品放置在干燥通风的室内,使自然风干(防止污染)。样品风干后,拣去杂质,研细并全部通过1mm孔径的筛子,装、封袋后储藏备用。
土壤全氮全磷用凯氏消化法测定,硝态氮和氨态氮用2mol/L KCl浸提,并用FIA star 5000全自动流动注射仪(瑞典FOSS公司生产)测定;土壤有机碳采用重铬酸钾氧化-外加热法测定;速效磷用0.5mol/L NaHCO3(pH值8.5)钼锑抗比色法测定;全钾用微波消解法消化,之后利用火焰分光光度计测定。
1.4 数据分析
采用SPSS 17.0和Microsoft Excel软件进行统计分析,应用单因素方差分析来比较不同株龄蒙古岩黄芪各形态指标的差异性;用主成分分析法(PAC)先对各指标的数据进行标准化,即将原始数据归一化处理成均值为0、方差为1的分析数据,通过对相似矩阵和协方差进行分析,提取特征值和特征向量,确定主要因子[16],并对前两个主成分进行排序(先算出每个主成分的得分再排序);最后用Person相关分析确定前面主成分分析得出的主要因子与根际土壤的全氮、铵态氮、硝态氮、全磷、速效磷、全钾及有机碳的关系。
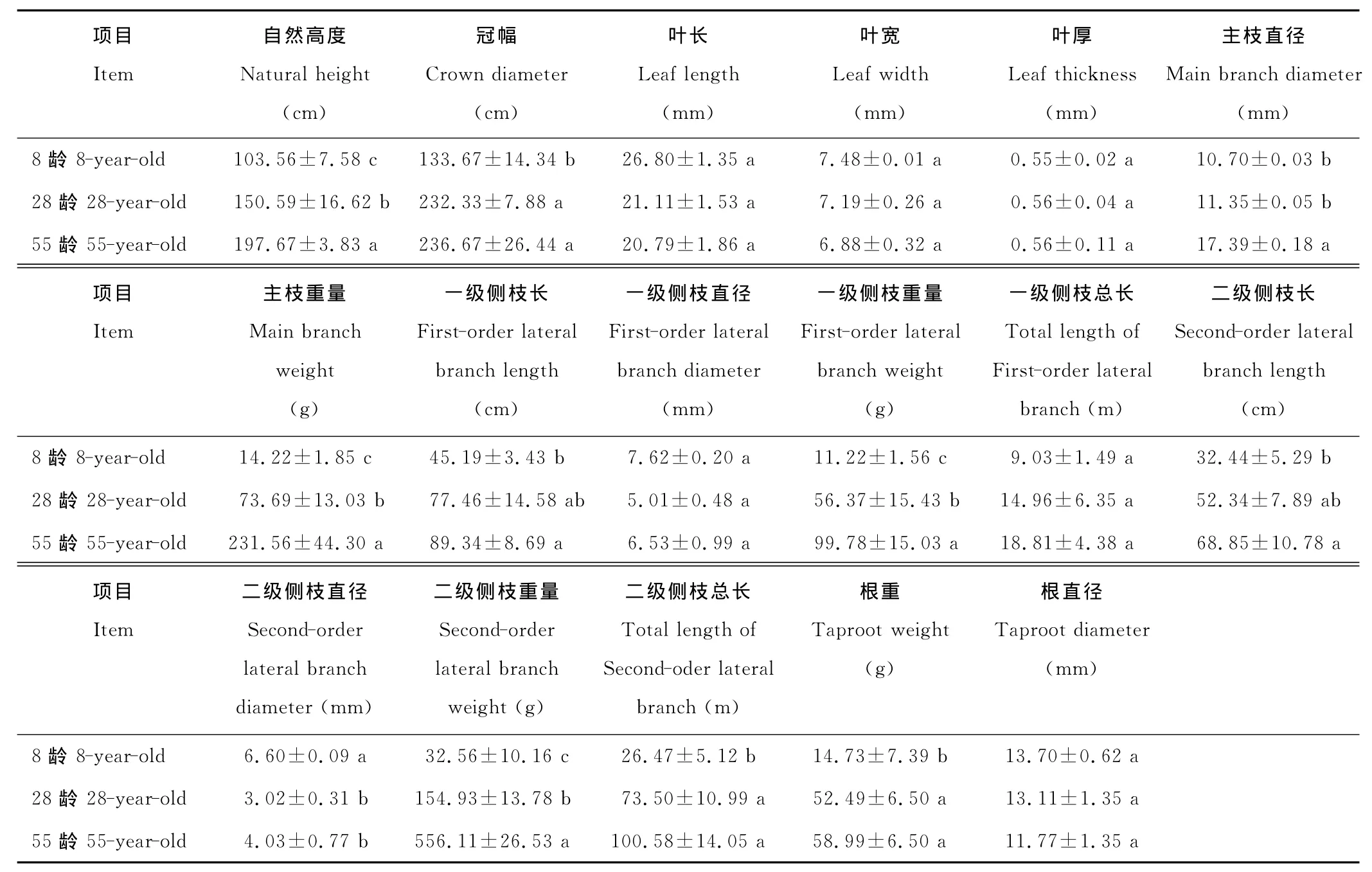
表1 不同株龄蒙古岩黄芪17个表型性状的差异Table 1 The difference about 17indicators of H.mongolicumin different ages
2 结果与分析
2.1 不同株龄蒙古岩黄芪的表型特征
如表1,不同株龄的蒙古岩黄芪,除了叶长、叶宽、一级侧枝直径、二级侧枝直径及根直径之外,其余指标都是55龄最高,且均是8龄<28龄<55龄,其中自然高度,主枝重量,一级侧枝及二级侧枝重量在各年份中的值差异显著,8,28,55龄的自然高度分别为103.6,150.6和197.7cm,三者的一级侧枝重量分别为11.2,56.4和99.8 g。差异幅度最大的是主枝重量和二级侧枝重量,55龄的(分别为231.6与556.1g)比8龄的(14.2与32.6g)分别增加了15和16倍。其次,最高值比最低值增加倍数的大小顺序是:一级侧枝重量(7.9倍)>根重(3.0倍)(55龄59.0g,8龄14.7g)>二级侧枝总长(2.8倍)(55龄100.6m,8龄26.5m)>二级侧枝直径(2.2倍)(28龄3.0mm,8龄6.6mm)>二级侧枝长(1.1倍)(55龄68.9cm,8龄32.4cm)>一级侧枝总长(2.1倍)(55龄18.8m,8龄9.0m)。剩余指标最大值比最小值增加百分数的排列顺序是:一级侧枝长(97.6%)(55龄89.3 cm,8龄45.2cm)>自然高度(91.0%)>冠幅(77.0%)(55龄236.7cm,8龄133.7cm)>主枝直径(62.6%)(55龄17.4mm,8龄10.7mm)>一级侧枝直径(51.9%)(28龄5.0mm,8龄7.6mm)>根直径(17.1%)(8龄13.7mm,55龄11.7mm)>叶厚(1.8%)(55龄0.56mm,8龄0.55mm)。而比较特殊的是叶长(8,28和55龄分别为26.8,21.1和20.8mm)和叶宽(7.5,7.2和6.9mm),株龄小的反而大(图中平均值为绝对平均值)。
2.2 不同株龄蒙古岩黄芪形态变异主成分分析
通过对不同株龄蒙古岩黄芪各指标主成分分析,找出了4个主成分(特征值大于1)(表2),其中前两个主成分累积贡献率达78.4%,基本可以概括生长情况的大部分信息,所以只取前两个主成分,表3列出了各指标对前两个主成分的载荷系数,即特征向量。第一主成分的特征值为9.428,贡献率达55.5%,自然高度(0.946)、根重(0.910)、二级侧枝重量(0.892)、二级侧枝总长(0.891)、冠幅(0.850)、主枝重量(0.845)、一级侧枝长(0.822)这7个指标所占的负荷量绝对值较大。说明这些指标之间有很高的相关性,存在信息上的重叠,它们代表了蒙古岩黄芪枝、根重量和长度方面的信息。第二主成分占总效益权重的22.907%(特征值为3.894),特征向量较大的是一级侧枝直径(0.819)和根直径(0.810),代表了蒙古岩黄芪根、枝直径方面的信息。这里选取了负荷量绝对值大于0.8的指标作为主要因子,因此,自然高度、根重、二级侧枝重量、二级侧枝总长、冠幅、主枝重量、一级侧枝长、一级侧枝直径及根直径9个指标是造成不同株龄蒙古岩黄芪表型差异的主要因素。

表2 主成分的特征值、贡献率和累积贡献率Table 2 The character value,contribution proportion and cumulative contribution proportion
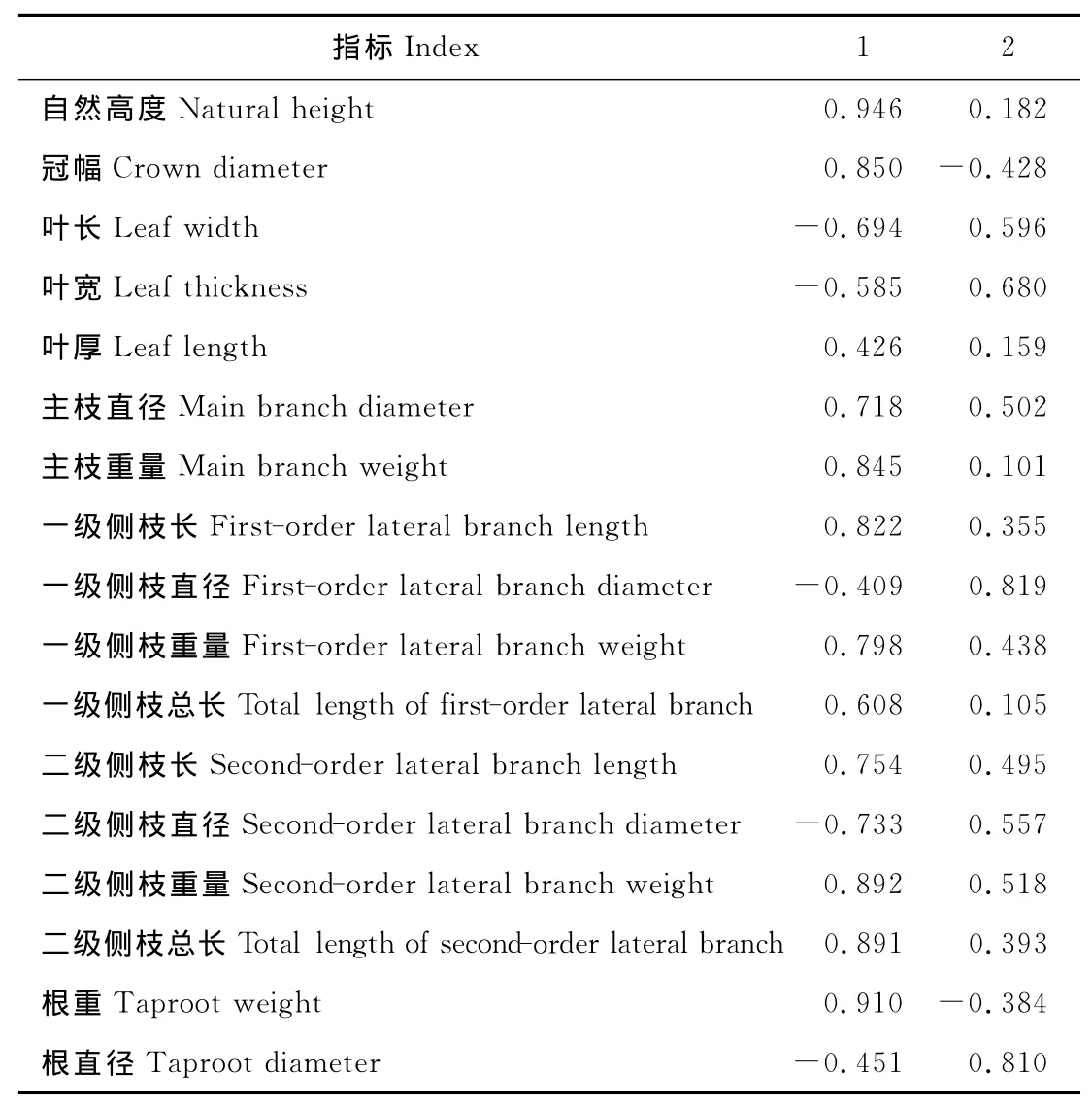
表3 17个性状对前2个主成分的负荷量Table 3 Loading dose of two principal components for seventeen characters
2.3 不同株龄蒙古岩黄芪前两个主成分排序
通过对第一、二主成分排序,可以看出在第一主成分上不同株龄的蒙古岩黄芪随年龄的增加而划分为3个较为明显的区域,其中1,2,3指55龄的3个重复,4,5,6指28龄的3个重复,依此,1,2,3是8龄的3个重复。且55与28龄蒙古岩黄芪居群之间的距离小于28与8龄之间的距离。除第一主成分外,其余各主成分说明的是样本内部的各方面特征,这些重复在第二主成分上排序较为混乱(图1)。
2.4 不同株龄蒙古岩黄芪形态差异的主要因子与根际土壤生境因子的相关性分析
由表4可以看出,在P<0.05的水平上时,一级侧枝长,一级侧枝重量及二级侧枝总长与氨态氮含量呈显著负相关(相关系数为分别为-0.767,-0.763,-0.721),一级侧枝直径与速效磷和全钾含量呈显著负相关关系(相关系数分别为-0.794和-0.792),其他指标与全氮、全磷、有机碳、氨态氮、硝态氮、速效磷和全钾有相关性,但相关系数不大,即相关关系不显著。
综上,由表2可以看出,3个不同株龄中,55龄蒙古岩黄芪长势最好,方差分析很好地反映了不同株龄蒙古岩黄芪间各形态指标的变异情况,但由于指标多,为了用尽可能少的维数去反映样本之间的联系,进行降维,分析结果显示蒙古岩黄芪17个表型性状最后可以归为4个主要成分,由于前两个主成分占总效益权重的78.365%,因此这两个主要成分就可以反馈原来变量所要反映的主要信息。而自然高度、冠幅等9个指标所占的负荷量大,故是造成这种变异的主要因子。相关性分析表明氨态氮,速效磷和全钾的含量与这些主要因子中的某些因子之间存在显著的相关关系。
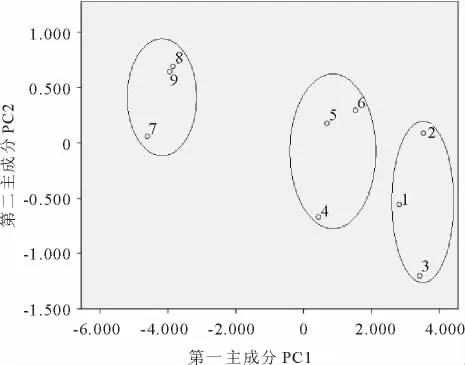
图1 第一、二主成分排序图Fig.1 Coordinate diagram of principal components

表4 各主要因子与根际土壤养分的相关性系数矩阵Table 4 The correlation coefficient matrix between every main factors and the nutrient of the rhizosphere soil
3 讨论
种内变异是个体和居群在时空变化发展中的产物,对变异规律的深入研究,使我们可以全面准确地了解种内个体的形态差异,对揭示物种的形成与进化机制提供帮助[16]。通过本文研究发现,随着株龄的增长,蒙古岩黄芪的部分表型形状发生了不同程度的变异,呈不均衡性,表明这些性状对环境适应的敏感程度不同。
选择合适的数学模型对所得数据进行分析,有助于揭示变异规律以及对环境的适应能力[17]。本文先计算出不同株龄蒙古岩黄芪随时间变异的总趋向及差异性。发现55龄的蒙古岩黄芪长势优于其他株龄,除叶长、叶宽、一级侧枝直径、二级侧枝直径、根直径之外,其他指标都随年龄增加呈增大趋势,并且自然高度和一级侧枝重量这两个指标在各株龄间差异显著。主枝重量和二级侧枝重量变异最大。这些现象说明蒙古岩黄芪在干旱沙化地区生长的过程中,随着株龄的增加,地上和地下指标为适应环境变化向最优、最适宜生存的方向发展,即在干旱沙地的恶劣气候环境下,生长和繁衍所采取的向空间扩展来提高对空间资源利用的生存策略,使蒙古岩黄芪更容易获得水分、光照和养分等[18-19]。叶片是植物的重要组成部分,叶长、叶宽随株龄减小而增大,这与主成分分析出来的结果(载荷量为-0.694,-0.585的负向作用)一致,然而叶厚呈递增趋势,引起这一现象的原因可能是由于株龄大的蒙古岩黄芪面临较多的逆境而形成的适应缺水,强光,温差大等环境气候来减少水分损失和保持养分的机制[20-21]。
主成分最大的优点就是方差贡献率大,其中各性状载荷值的大小体现了各性状在主成分中的重要程度,于是自然高度、根重、二级侧枝重量、二级侧枝总长、冠幅、主枝重量、一级侧枝长、一级侧枝直径及根直径这9个指标所占负荷量较大,故是造成表型差异的主要因素,也就是蒙古岩黄芪的枝与根的重量和直径、枝的长度能反映出其生长的大体情况。因此在育种时注意这些方面可以来评价蒙古岩黄芪种的优劣性。这些主要因素的变异越大说明植株适应环境的能力越强,丰富的变异有助于保持物种和生态系统的多样性和稳定性[22-23]。赵俊卉等[24]对长白山云杉、冷杉和红杉的研究发现,竞争是引起枝长和直径变异的主要因素。
对各重复前两个主成分进行排序,在第一数轴上,不同株龄的重复在时间增加的基础上进行分类,聚为3个不同年份的团体,与实际情况相吻合,即个体形态特征在一定程度上发生了分化[25]。这个结果与第一主成分综合原始指标信息的能力最强,可以全面反映各指标状况的综合指标,能说明原始数据变动的总趋势的功能相呼应[26],很好地反映了蒙古岩黄芪随年龄增加表型性状在可遗传的、有序的、稳定的变异轨迹中的变化,符合进化论的基本规律,表明蒙古岩黄芪植株在形态学功能上对时间变异的适应性,同时也印证了关系越密切的植株排序越近的理论[27]。类似的实验结果有:通过环境因子对青南地区植物群落进行排序发现植物群落在空间地理上分布很有规律[28]。
影响生物表型变异的因素主要有两个,即遗传因素和环境因素。遗传因子是受基因控制对植物表型变异影响较为缓慢、稳定,而环境因子能够在短期内影响和改变植物表型变异,二者对种内形态变异相辅相成,共同控制着表型变异的趋向[29-30]。影响生物表型的环境因子[16]主要包括:土壤环境、物候环境,种的生态分布区等。本研究从土壤环境这方面进行研究,对9个主要因子与之对应的根际土壤全氮等7个养分因子做了相关性分析,一级侧枝长度,二级侧枝总长及重量与氨态氮之间存在显著的负相关性(即氨态氮与第一主成分的三个主要因子的相关关系显著,因此可以把第一主成分的主要因子看作是氨态氮主控因子),速效磷和全钾与一级侧枝直径之间也呈显著的负相关性。表明蒙古岩黄芪的这9个主要因子与土壤养分存在一定的联系,这是因为植物在生长的过程中受环境影响,同时也影响着土壤环境[31-33],而作为影响蒙古岩黄芪的主要因子,与土壤养分之间必然有密切的关系。土壤是蒙古岩黄芪营养元素的主要源泉,而蒙古岩黄芪根系、落叶物残体为土壤提供腐殖质,并随着株龄的增加对土壤改良具有显著的效果[34]。氨态氮之所以与第一主成分的三个主因子成负相关关系,可能是因为在蒙古岩黄芪自身生长过程中吸收了大量氨态氮以使侧枝快速生长。表型变异是变异的重要线索,本文从植物进化年龄,即株龄变化这个方向对蒙古岩黄芪表型性状进行了研究,但这种变异具体是因遗传因素占的份额大还是环境因素大,还有待于从分子方面进行进一步研究。
Reference:
[1]Hao C Y,Wu S H,Yang Q Y.Research on relationship between sandy desertification and land-use in Mu Us Region.Journal of Desert Reseach,2005,25(1):33-39.
[2]Guo J,Wang T,Han B S,etal.Dynamic process of aeolian desertification land variation in Mu Us sandy land and its surrounding area in recent 30years.Journal of Desert Reseach,2008,28(6):1017-1021.
[3]Zhang C S,Liu G B,Xue S,etal.Anti-erodibility and microaggregate fractal features of soils under different psammophytes.Bulletin of Soil and Water Conservation,2012,32(2):1-6.
[4]Yang D H,Zhao Y S.Effects of site condition on growth and spatial variations ofHedysarumlaeve.Scientia Silvae Sinicae,2011,47(2):180-186.
[5]Lv Y L.The preliminary observation on the shape and anatomical structure on rhizome ofHedysarummongolicum.Journal of Beijing Forestry University,1986,(1):91-95.
[6]Gu S.The relationship between the growth characteristics and climatic factors ofHedysarumscoparium,HedysarumleaveandKorshinsk peashrubat seedling stage.Arid Zone Research,1994,11(2):60-63.
[7]Jiang G M.Species and habitat variablity of photosynthesis,transpiration and water use efficiency of different plant speicies in Maowusu sand area.Acta Botanica Sinica,1999,41(10):1114-1124.
[8]Guo J P,Gao S H,Wang L M,etal.Response ofHedysarummongolicumturcz.to high CO2concentration and soil drought stress.Journal of Plant Resources and Environment,2002,11(1):14-16.
[9]Ge S,Hong D Y.Biosystematic studies onAdenophorapotaniniikorsh.Complex(campanulaceae).Acta Phytotaxonomica Sinica,1995,33(5):433-443.
[10]Zhang S P,Wang R Q,Zhang Z G,etal.Study on morphological variation ofPhragmitesaustralisin the Yellow River downstream wetland.Acta Phytoecologica Sinica,2003,27(1):78-85.
[11]Zhao X L,Ling Y,Zhang G F,etal.Community characteristics of beach wetland vegetations along a habitat gradient in Dafeng Milu reserve of Jiangsu province.Chinese Journal of Ecology,2010,29(2):244-249.
[12]Liang K L,Jiang W Q,Zhou Z Y,etal.Variation in main morphological characteristics ofAmorphafruticosaplants in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2012,32(1):311-318.
[13]Yu S X.Introduction to mathematical ecology.Beijing:Science and Technology Publishing House,1995:207-213.
[14]Li W H.The research on the difference of nitrogen content and pH between rhizosphere and bulk soil ofQuercusvariabilis,Cunninghamia lanceolataandPinustaeda.Journal of Nanjing Forestry University,1996,20(2):49-52.
[15]Zeng S C,Su Z Y,Chen B G,etal.A review on the rhizospherenutritionecology research.Journal of Nanjing Forestry University(Nature Science Edition),2003,27(6):79-83.
[16]Yang J.Infraspecific variation in plant and the exploring methods.Journal of Wuhan Botanical Research,1991,9(2):185-195.
[17]Sneath P,Sokal R.Mumerical Taxonomy[M].Beijing:Science Press,1984.
[18]He M Z,Zhang J G,Wang H,etal.Analysis of branching architecture factors of desert plants.Journal of Desert Reseach,2006,26(4):625-630.
[19]Zhao Z J.The Study of Branch and Root Morphometries ofMedicugofalcate[D].Changchun:Northeast Normal University,2007.
[20]Lambrecht S C,Dawson T E.Orrelated variation of floral and leaf traits along a moisture availability gradient.Oecologia,2007,151(4):574-583.
[21]Santiago L S,Kitajima K,Wright S J,etal.Coordinated changes in photosynthesis,water relations and leaf nutritional traits of canopy trees along aprecipitation gradient in lowland tropical forest.Oecologia,2004,139(4):495-502.
[22]Yu H,Kiang Y T.Genetic variation in South Korean natural populations of wild soybean(Glycinesoja).Euphytica,1993,68(3):213-221.
[23]Jiang X L,Yue J,Zhang W G,etal.Biodiversity,ecosystem functioning and spatio-temporal scales.Acta Prataculturae Sinica,2010,19(1):219-225.
[24]Zhao J H,Kang X G,Zhang H D,etal.Relationships between coefficient of variation of diameter and height and competition index of main coniferous trees in Changbai Mountains.Chinese Journal of Applied Ecology,2009,30(8):1832-1837.
[25]Ma H J,He P,Deng H P,etal.Analyses of morphological differentiation within six populations ofMalushupehensis(Rosaceae).Journal of Southwest China Nomal University(Natural Science),2002,27(4):559-562.
[26]Zhang P.The Principal Component Analysis Based Research in Comprehensive Assessment[D].Nanjing:Institutes of Technology of Nanjing,2004.
[27]Luan Q S,Sun J,Song S,etal.Canonical correspondence analysis of summer phytoplankton community and its environment in the Yangtze river estuary,China.Journal of Plant Ecology(Chinese Version),2007,31(3):445-450.
[28]Wang W Y,Wang C Y,Wang Q J,etal.The gradient analysis of plant communities at sounthern Qinghai of Qinhai-Tibet Plateau.Journal of Lanzhou University(Natural Science),2004,40(3):60-65.
[29]Soltis P S,Soltis D E.Genetic variation in endemic and wide spread plant species:examples from Saxifragaceae and Polystichum(Dryopteridaceae).Aliso,1991,13:215-223.
[30]Ge S.Genetic Diversity[M].Hangzhou:Zhejiang Science and Technology Press,1997:11-19.
[31]Huang Y P,Chen Y,Guo F X,etal.Effect of N,P,K combinations fertilizion on the cultivation quality ofGlycyrrhizauralensisseedlings.Acta Prataculturae Sinica,2012,21(2):233-240.
[32]Zhu Q,Pan Y Z,Zhao L.etal.The effects of N,P,K,and Ca on plant growth and nutrient content of Lily leaves.Acta Prataculturae Sinica,2012,21(5):274-284.
[33]Zhang L,Wang C T,Liu W,etal.Relationships of dominant species root activity,plant community characteristics and soil micro-environment in artificial grassland over different cultivation periods.Acta Prataculturae Sinica,2012,21(5):185-194.
[34]Wang Y W,Liao C Y,Sun C Z,etal.Soil chemical properties of sand-fixing forests in Maowusu Sandland.Chinese Journal of Soil Science,2009,40(4):776-780.
[1]郝成元,吴绍洪,杨勤业.毛乌素地区沙漠化与土地利用研究.中国沙漠,2005,25(1):33-39.
[2]郭坚,王涛,韩邦帅,等.近30a来毛乌素沙地及其周边地区沙漠化动态变化过程研究.中国沙漠,2008,28(6):1017-1021.
[3]张昌胜,刘国彬,薛萐,等.不同沙生植被土壤微团聚体分形特征及抗蚀性.水土保持通报,2012,32(2):1-6.
[4]杨东华,赵雨森.不同生境对踏郎生长及土壤养分空间差异的影响.林业科学,2011,47(2):180-186.
[5]吕悦来.蒙古岩黄茵根状茎的形态及解剖构造的初步观察.北京林业大学学报,1986,(1):91-95.
[6]古松.花棒、杨柴、毛条苗期生长特性与气候因子的关系.干旱区研究,1994,11(2):60-63.
[7]蒋高明.毛乌素沙地若干植物光合作用、蒸腾作用和水分利用效率种间及生境间差异.植物学报,1999,41(10):1114-1124.
[8]郭建平,高素华,王连敏,等.杨柴对高CO2浓度和土壤干旱胁迫的响应.植物资源和环境学报,2002,11(1):14-16.
[9]葛颂,洪德元.泡沙参复合体(桔梗科)的物种生物学研究Ⅲ.性状的遗传变异及其分类价值.植物分类学报,1995,33(5):433-443.
[10]张淑萍,王仁卿,张治国,等.黄河下游湿地芦苇形态变异研究.植物生态学报,2003,27(1):78-85.
[11]赵小雷,凌云,张光富,等.大丰麋鹿保护区不同生境梯度下滩涂湿地植被的群落特征.生态学杂志,2010,29(2):244-249.
[12]梁坤伦,姜文清,周志宇,等.青藏高原紫穗槐主要形态特征变异分析.生态学报,2012,32(1):311-318.
[13]余世孝.数学生态学导论[M].北京:科学技术文献出版社,1995:207-213.
[14]厉婉华.栓皮栎、杉木和火炬松根际与非根际土壤氮素及pH差异的研究.南京林业大学学报,1996,20(2):49-52.
[15]曾曙才,苏志尧,陈北光,等.植物根际营养研究进展.南京林业大学学报(自然科学版),2003,27(6):79-83.
[16]杨继.植物种内形态变异的机制及其研究方法.武汉植物学研究,1991,9(2):185-195.
[17]史尼斯P,索卡尔R.数值分类学[M].赵铁桥,译.北京:科学出版社,1984.
[18]何明珠,张景光,王辉.荒漠植物枝系构型影响因素分析.中国沙漠,2006,26(4):625-630.
[19]赵占军.黄花苜蓿分枝与根系形态的研究[D].长春:东北师范大学,2007.
[23]江小雷,岳静,张卫国,等.生物多样性,生态系统功能与时空尺度.草业学报,2010,19(1):219-225.
[24]赵俊卉,亢新刚,张慧东,等.长白山主要针叶树种胸径和树高变异系数与竞争因子的关系.应用生态学报,2009,30(8):1832-1837.
[25]马洪菊,何平,邓洪平,等.湖北海棠不同居群形态变异的数量分析.西南师范大学学报(自然科学版),2002,27(4):559-562.
[26]张鹏.基于主成分分析的综合评价研究[D].南京:南京理工大学,2004.
[27]栾青杉,孙军,宋书,等.长江口夏季浮游植物群落与环境因子的典范对应分析.植物生态学报,2007,31(3):445-450.
[28]王文颖,王春燕,王启基,等.青藏高原青南地区植物群落生态梯度分析.兰州大学学报(自然科学版),2004,40(3):60-65.
[30]葛颂.遗传多样性[M].杭州:浙江科学技术出版社,1997:11-19.
[31]黄亚萍,陈垣,郭凤霞,等.氮磷钾配施对甘草育苗质量的影响.草业学报,2012,21(2):233-240.
[32]朱峤,潘远智,赵莉.氮、磷、钾、钙对香水百合生长及叶片养分含量的影响.草业学报,2012,21(5):274-284.
[33]张莉,王长庭,刘伟,等.不同建植期人工草地优势种植物根系活力、群落特征及其土壤环境的关系.草业学报,2012,21(5):185-194.
[34]王彦武,廖超英,孙长忠,等.毛乌素沙地固沙林土壤化学性质差异.土壤通报,2009,40(4):776-780.
