不同放牧强度下温性草甸草原土壤微生物群落结构PLFAs分析
2015-01-02谭红妍闫瑞瑞闫玉春陈宝瑞辛晓平
谭红妍,闫瑞瑞,闫玉春,陈宝瑞,辛晓平*
(1.中国农业科学院资源区划所,北京100081;2.呼伦贝尔草原生态系统国家野外科学观测研究站,北京100081)
土壤微生物群落是草地地下生态系统的重要组成成分,其不仅是土壤养分重要的“源”和“库”,而且介导许多控制碳氮循环的关键过程,在草地生态系统过程和功能的维持中扮演着至关重要的角色[1-2]。微生物生命周期短,对生存的微环境十分敏感,土壤微生物群落成为判别干扰条件下土壤生态系统状态的重要生物学指标。脂类物质是构成活体微生物细胞膜的主要成分,它在细胞中含量稳定约占细胞干重的5%,PLFA即为甲基化土壤中提取磷脂成分后得到的脂肪酸产物[3]。不同的微生物通过不同的代谢途径形成不同的PLFA,可以作为一种生物标记来鉴定土壤中微生物的种类和丰富度。近年来,PLFA技术以其快速、准确、重现性好、对试验条件要求较低等优点[4],被广泛用于土壤中微生物群落结构的动态监测。
放牧是草地的主要利用方式之一,对土壤微生物乃至整个土壤生态系统都有深远影响。适度放牧有助于土壤微生物数量的增加,过度放牧会导致微生物数量明显减少;真菌和细菌对放牧强度的响应比放线菌更为敏感[5-6],随着放牧强度的增加,真菌、细菌数量显著下降,放线菌数量升高。赵帅等[7]发现放牧干扰下土壤微生物的PLFAs种类发生改变,细菌与真菌PLFAs比值显著升高,革兰氏阳性菌与阴性菌PLFAs含量比值显著下降。Clegg[8]对英格兰西南一草地研究发现,放牧可以显著提高革兰氏阳性菌和阴性菌的PLFAs含量,放线菌与真菌的指示性PLFAs变化不显著。而Ingram等[9]在美国怀俄明州北部的一个半干旱混合草地研究不同放牧强度对微生物群落结构的影响时发现,不同处理并没有使表征各微生物类群 的PLFAs含量发生显著变化,在轻度放牧区总PLFAs含量略高。
放牧对地上植被的影响研究的报道较多,对土壤性状影响的研究也有一些报道,但大多数的研究内容都局限在土壤三大类群微生物(细菌、真菌、放线菌)的数量、土壤酶活性等研究上。本研究立足于呼伦贝尔草原连续5年的肉牛控制放牧样地,采用磷脂脂肪酸技术研究不同放牧强度对土壤微生物群落结构的影响,旨在揭示长期放牧下土壤微生物多样性及群落结构的演变状况,为深入了解放牧系统地下生态学过程、合理利用草地资源提供依据。
1 材料与方法
1.1 研究区概况
本研究在中国农业科学院呼伦贝尔草地生态系统试验站的肉牛控制放牧样地中进行。研究点位于呼伦贝尔市谢尔塔拉牧场场部东3km,属于大兴安岭西麓丘陵向内蒙古高原的过渡区,地理位置为49°32′-49°34′N,119°94′-119°96′E,海拔670~677m。研究区属于温带半干旱大陆性气候,年均温-3~1℃,≥10℃年积温1580~1800℃,无霜期110d左右,年平均降水量350~400mm,降水期多集中在7-9月。土壤为暗栗钙土。植被类型为羊草+杂草类,主要物种有羊草(Leymuschinensis)、裂叶蒿(Artemisiaincise)、苔草(Carextristachva)、贝加尔针茅(Stipabaicalensis)、展枝唐松草(Thalictrumsquarrosum)、日荫菅(Carexpediformis)、蓬子菜(Galiumverum)、麻花头(Serratulacentauroides)等,伴生种有狭叶柴胡(Bupleurumscorzonerifolium)、斜茎黄芪(Astragalusadsuigens)、山野豌豆(Viciaamoena)、草地早熟禾(Poapratensis)等。
1.2 试验设计及样品采集
放牧样地共存在6个水平的放牧梯度处理,载畜率分别为 G0.00:0.00Au/hm2、G0.23:0.23Au/hm2、G0.34:0.34Au/hm2、G0.46:0.46Au/hm2、G0.69:0.69Au/hm2、G0.92:0.92Au/hm2。其中以500kg肉牛为一个标准家畜肉牛单位,用250~300kg的放牧肉牛头数来控制不同的放牧强度,6个放牧梯度肉牛头数分别为0,2,3,4,6,8头,共用69头肉牛。每种处理3个重复,共包括15个放牧区和3个对照封育区。各处理的分布遵循随机区组试验设计原则,每个小区为300m×160m,试验区总面积90hm2(包括牧道),具体设计见图1。从2009年开始,每年6月中旬开始放牧,10月中旬结束,为期120d。整个放牧期肉牛日夜均在放牧区,肉牛饮水通过拉水供应。
本项研究中选取了4个放牧强度:对照区G0.00、轻度放牧 G0.23、中度放牧 G0.46、重度放牧 G0.92,野外采样时间为2013年08月15日至19日。每个试验小区随机设置3个采样点,每个采样点用直径5cm的土钻取0~20cm土壤(3点混合),剔除石块和土壤中的动植物残体后,采用“四分法”保留约500g土样。一部分土样带回实验室后立即过2mm筛保存于-80℃超低温冰箱;另一部分土样自然风干,用于测定土壤基本养分。

图1 肉牛不同放牧强度试验平面图Fig.1 Design diagram of cattle different grazing gradients

表1 估算微生物生物量的脂肪酸Table 1 PLFA for calculating soil microbial biomass
1.3 分析指标与方法
1.3.1 土壤磷脂脂肪酸提取与分析 将置于超低温冰箱中的土样放入超低温冷冻干燥机中处理后,取3.00g左右的冻干土,利用Blight/Dyer法通过氯仿-甲醇-柠檬酸缓冲液震荡提取总脂,经SPE柱层析分离得到磷脂脂肪酸;将得到的磷脂脂肪酸甲酯化。PLFA的定性与定量分析用HP6890气相色谱和Sherlock软件,以正十九烷脂肪酸甲酯为内标物。各类群微生物标记如表1所示。命名法采用X:YωZ,其中X是指主链碳原子个数;Y为双键个数;Z为甲基链离双键的位置。i,a,Me分别表示异构、反异构、甲基支链,cy表示环丙基。
1.3.2 土壤基本养分分析 土壤有机质用重铬酸钾容量法—外加热法进行测定;全磷采用NaOH熔融-钼锑抗比色法;全氮采用半微量凯氏定氮法;土壤pH采用电位法;速效磷采用0.5mol/L NaHCO3浸提-钼锑抗比色法;速效钾采用NH4OAC浸提-火焰光度法;碱解氮采用1.0mol/L NaOH碱解扩散法[16]。
1.4 数据分析
微生物群落结构用CANOCO 4.5中的除趋对应分析(detrended correspondence analysis,DCA)并作图。其余数据用SPSS 20.0分析,Excel作图。
2 结果与分析
2.1 不同放牧强度下温性草甸草原土壤中所含PLFA的特征
本实验共检测出44种磷脂脂肪酸生物标记,其中G0.23处理下PLFA种类最多,G0.92处理下最少。不同放牧强度下土壤的PLFA种类存在明显差异,比如15:0iso 3OH 与18:1 2OH 为 G0.23放牧处理所特有,16:1ω9c为G0.46放牧处理所特有。但4个放牧梯度下优势类群并未发生改变,均为16:00、16:0 10-methyl、18:1ω7c、19:0 cycloω8c,它们含量之和所占比例分别为36.06%,35.00%,36.84%,34.80%。表明不同放牧强度下磷脂脂肪酸组成改变,即放牧改变了土壤微生物的区系组成和丰富度,却不会影响优势菌的地位(表2)。
2.2 不同放牧强度对土壤微生物各菌群生物量的影响
微生物区系组成中,细菌比例最高,其次为放线菌、真菌、AMF。不同放牧强度下,各大类微生物的特征标记PLFAs含量发生了改变(图2)。土壤总微生物量随着放牧强度的增加表现为先减少后增加的趋势,即为对照区最高,中度放牧G0.46最低,重度放牧G0.92出现小幅度的回升;细菌、革兰氏阴性菌、腐生真菌、AMF生物量也呈现相同变化。革兰氏阳性菌、放线菌生物量则随着放牧强度的增加而增加(表3)。此外,AMF对于放牧干扰最为敏感(各放牧强度下极显著差异P<0.01)。

表2 不同放牧强度下土壤微生物主要磷脂脂肪酸构成特征(平均值±标准误)Table 2 The profiles of main PLFAs of microbes in soils from different grazed degrees(Mean±SE)nmol/g
2.3 不同放牧强度对土壤微生物群落结构的影响
为了更直观了解不同放牧强度下土壤微生物群落的分布格局,对36个土样和主要的磷脂脂肪酸进行排序,其中前两个排序轴的特征值分别为0.526和0.167,累积贡献率之和为78.3%。排序图基本上能将4个放牧轻度的样方区分开,轻度放牧G0.23同对照区G0.00群落结构较为相似,中度放牧G0.46同重度放牧G0.92群落结构较为相似。表明当放牧干扰超过某一强度时,会引起土壤微生物群落结构大幅度的变化。

表3 不同放牧强度下土壤各微生物类群生物量(平均值±标准误)Table 3 The microbial groups PLFAs content from different grazed degrees(Mean±SE)nmol/g
图3为主要磷脂脂肪酸在除趋势对应分析中的因子载荷分析结果。第一排序轴上的载荷值较高的脂肪酸可以分为两类,表征真菌的脂肪酸(18:2ω6、16:1ω5)和表征革兰氏阴性菌的脂肪酸(17:1ω8、18:1ω9、18:1ω7),它们随着放牧强度的增加而减少。表征革兰氏阳性菌的脂肪酸(a17:0、a15:0、i14:0、i15:0、i16:0)和表征放线菌的脂肪酸(10Me18:0、10Me17:0、10Me16:0)在两排序轴载荷值都较低,说明这几种脂肪酸在中度放牧 G0.46含量最低。
2.4 各菌群PLFAs含量与土壤基本养分相关性分析
通过对各菌群PLFAs含量与土壤养分相关性分析表明(表4),土壤总PLFAs含量和细菌、真菌、革兰氏阴性菌、丛枝菌根真菌的PLFAs含量均同有机质、全磷、速效磷、速效钾呈显著正相关(P<0.05),尤其是腐生真菌、丛枝菌根真菌的PLFAs与有机质、速效磷相关水平达到了极显著程度(P<0.01)。由此说明真菌和AMF可用作评价土壤肥力与健康的生物指标,同时有机质、速效磷是影响微生物数量和种类的重要养分因素。相关分析结果还显示,革兰氏阳性菌、放线菌PLFAs含量同大多数养分指标成负相关,各菌群PLFAs含量与全氮、碱解氮、pH间无显著相关性。

图2 不同放牧梯度下土壤微生物群落PLFA的除趋势对应分析Fig.2 Detrended correspondence analysis of PLFA profiles from soil microbialcommunities of different grazed degrees
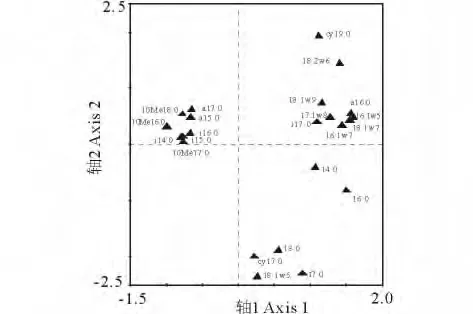
图3 不同放牧梯度下土壤微生物群落PLFA载荷因子贡献Fig.3 Eigenvector loadings of PLFA contributing to soil communities ordination pattern of different grazed degrees

表4 PLFAs与土壤养分相关性分析Table 4 Correlation between soil PLFAs content and soil nutrients
3 讨论
放牧干扰既引起植物生产力的下降,也降低了植物多样性。生产力下降导致地上的叶凋落物、地下死亡根系的凋落物和根系分泌物的减少,使得微生物可利用营养物减少[17],最终引起微生物数量的下降。物种多样性降低会引起植物产物的改变和凋落物有机组分的变化[18-22],从而影响微生物群落的组成和功能。与王启兰等[20]、Raiesi和Asadi[22]研究结果一致:随着放牧压力的增大,土壤总微生物显著下降。放牧显著降低了地上生物量和活根生物量,微生物分解底物减少。此外,植被盖度的下降以及牲畜的过度践踏,引起土壤的含水量、透水性和透气性的下降[20],土壤微环境遭到破坏,也会降低微生物的繁殖速率。
不同菌群微生物因自身生长繁殖特性不同,对放牧干扰存在不同的响应。革兰氏阴性菌与腐生真菌生物量的下降,干扰的机制可能是:通过改变土壤环境来直接影响微生物生长,比如有机质含量、养分的有效性[2],并且家畜活动会严重破坏腐生真菌的菌丝。而在重度放牧G0.92中,革兰氏阴性菌、真菌、AMF和总微生物量都出现小幅回升,原因可能在于不稳定基质(尿液与粪便)的输入为微生物提供易利用的营养元素,适当地促进微生物生长[21]。此外,本研究中 AMF生物量在4个放牧强度下极显著差异(P<0.01)。Jansa等[23]和Schnoor等[24]曾采用PLFA技术研究不同微生物菌群对外界干扰响应时也发现:AMF对干扰反映最为敏感。AMF与植物是共生关系,植物物种的变化对其影响最为显著。羊草为该研究区的主要优势种,也是牲畜较为喜欢的牧草之一。牧压的增加必然会引起羊草优势度的显著下降,那么与羊草存在共生关系的AMF因菌丝结构的破坏,其生物量定会显著下降。有研究指出[24],放牧干扰引起活根生物量的变化是影响微生物群落改变的主要原因,其中AMF为典型代表。随放牧强度的增加,革兰氏阳性菌和放线菌生物量呈现出增加的趋势,同多数文献研究结果不太一致,有待于进一步验证。Klumpp等[25]在寻找高强度放牧导致土壤碳丢失的原因时,指出高强度的干扰通过影响植物根系及由它们控制的微生物,加快了土壤中营养元素的循环。是不是可以有这样一种猜测:高强度放牧使得营养循环效率提高的原因在于,革兰氏阳性菌和放线菌生物量的增加?
DCA分析结果表明,过度放牧会引起土壤微生物群落结构的显著改变。本研究中取土深度为0~20cm,微生物大部分属于根际微生物[19]。微生物群落能够利用的能源物质主要为根际沉积物(根系分泌物、脱落根细胞、根残体),不同植物根系沉积物元素组成差异很大[26],最终引起异养微生物群落结构和功能的变化。当存在外部干扰时,植物将更多的营养元素分配给地下部分,根系各种元素的相对含量也会发生变化[27]。有机质、速效磷与微生物菌群极显著正相关,是微生物生长和活性的主要营养因子,可能植物本身碳、磷含量是影响微生物种类和数量的生物因素。
4 结论
放牧改变了土壤微生物的组成,但不影响功能菌的优势地位。土壤总微生物量随着放牧强度的增加表现为先减少后增加的趋势,即为对照区最高,中度放牧最低,重度放牧出现小幅度的回升;细菌、革兰氏阴性菌、腐生真菌、AMF生物量也呈现相同变化。革兰氏阳性菌、放线菌生物量则随着放牧强度的增加而增加。此外,AMF对于放牧干扰最为敏感(各放牧强度下极显著差异P<0.01)。除趋势对应分析结果表明,对照区和轻度放牧处理下土壤的微生物群落结构较为相似,中度放牧和重度放牧处理导致微生物群落结构发生大幅度改变。有机质、速效磷是影响微生物种类和数量的重要养分因素。
Reference:
[1]Bach E M,Baer S G,Meyer C K,etal.Soil texture affects soil microbial and structural recovery during grassland restoration.Soil Biology and Biochemistry,2010,42(12):2182-2191.
[2]Mårtensson L,Olsson P A.Reductions in microbial biomass along disturbance gradients in a semi-natural grassland.Applied Soil Ecology,2012,62:8-13.
[3]Wu Y P.Studies on soil microbial community structure based on phospholipid fatty acid(PLFA)analysis[D].Hangzhou:Zhejiang University,2009.
[4]Zhang Q F,Liu B,Lin Y Z,etal.The diversity of phospholipid fatty acid(PLFA)biomarker for the microbial community in soil.Acta Ecologica Sinica,2006,29(8):4127-4137.
[5]Yan R R,Yan Y C,Xin X P,etal.Changes in microorganisms and enzyme antivities in soil under different grazing intensities in meadow steppe,Inner Mongolia.Ecology and Environmental Sciences,2011,20(2):259-265.
[6]Zhang C X,Nan Z B.Changeable characteristics of three soil microbial groups under different grazing intensities in Loess Plateau.Pratacultural Science,2010,27(11):131-136.
[7]Zhang S,Zhang J N,Lai X,etal.Analysis of microbial biomass C,N and soil microbial community struture ofStipasteppes using PLFA at grazing and fenced in Inner Mongolia,China.Journal of Agro-Environment Science,2011,30(6):1126-1134.
[8]Clegg C D.Impact of cattle grazing and inorganic fertiliser additions to managed grasslands on the microbial community composition of soils.Applied Soil Ecology,2006,31(1-2):73-82.
[9]Ingram L J,Stahl P D,Schuman G E,etal.Grazing impacts on soil carbon and microbial communities in a mixed-grass ecosystem.Soil Science Society of America Journal,2008,72(4):939.
[10]Allison V J,Miller R M,Jastrow J D,etal.Changes in soil microbial community structure in a tallgrass prairie chronosequence.Soil Science Society of America Journal,2005,69(5):1412.
[11]Zelles L.Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil:a review.Biology and Fertility of Soils,1999,29(2):111-129.
[12]Zogg G P,Zak D R,Ringelberg D B,etal.Compositional and functional shifts in microbial communities due to soil warming.Soil Science Society of America Journal,1997,61(2):475.
[13]Hu L,Wang C T,Wang G X,etal.Changes in the activities of soil enzymes and microbial community structure at different degradation successional of alpine meadows in the headwater region of Three Rivers,China.Acta Prataculturae Sinica,2014,23(3):8-19.
[14]Bardgett R D,Hobbs P J,Frostegard A.Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland.Biology and Fertility of Soils,1996,22:261-264.
[15]Olsson P A.Signature fatty acids provide tools for determination of the distribution and interactions of mycorrhizal fungi in soil.FEMS Microbiology Ecology,1999,29:303-310.
[16]Bao S D.Soil agricultural chemistry analysis(The third edition)[M].Beijing:China Agriculture Press,2000:28-49.
[17]Xia B C.Effect of vegetation on structure on soil microbial community.Chinese Journal of Applied Ecology,1998,9(3):296-300.
[18]Hu Y L,Wang S L,Yan S K.Research advances on the factors influencing the activity and community structure of soil microorganism.Chinese Journal of Soil Science,2006,37(1):170-176.
[19]Wang G H,Jin J,Xu M N,etal.Effect of plant,soil and soil management on soil microbial community diversity.Chinese Journal of Ecology,2006,25(5):550-556.
[20]Wang Q L,Wang C T,Du Y G,etal.Grazing impact on soil microbial biomass carbon and relationship with soil environment in alpineKobresiameadow.Acta Prataculturae Sinca,2008,17(2):39-46.
[21]Wang K H,Mcsorley R,Bohlen P,etal.Cattle grazing increases microbial biomass and alters soil nematode communities in subtropical pastures.Soil Biology and Biochemistry,2006,38(7):1956-1965.
[22]Raiesi F,Asadi E.Soil microbial activity and litter turnover in native grazed and ungrazed rangelands in a semiarid ecosystem.Biology and Fertility of Soils,2006,43(1):76-82.
[23]Jansa J,Mozafar A,Anken T R,etal.Diversity and structure of AMF communities as affected by tillage in a temperate soil.Mycorrhiza,2002,12(5):225-234.
[24]Schnoor T K,Lekberg Y,Rosendahl S,etal.Mechanical soil disturbance as a determinant of arbuscular mycorrhizal fungal communities in semi-natural grassland.Mycorrhiza,2011,21(3):211-220.
[25]Klumpp K,Fontaine S,Attard E,etal.Grazing triggers soil carbon loss by altering plant roots and their control on soil microbial community.Journal of Ecology,2009,97(5):876-885.
[26]Donald R Z,William E H,David C W,etal.Plant diversity,soil microbial communities,and ecosystem function:are there and links.Ecology,2003,84(8):2042-2050.
[27]Zhang T,Weng Y,Yao F J,etal.Effect of grazing intensity on ecological stoichiometry ofDeyeuxiaangustifoliaand meadow soil.Atca Prataculturae Sinica,2014,23(2):20-28.
[3]吴愉萍.基于磷脂脂肪酸(PLFA)分析技术的土壤微生物群落结构多样性的研究[D].杭州:浙江大学,2009.
[4]张秋芳,刘波,林营志,等.土壤微生物群落磷脂脂肪酸PLFA生物标记多样性.生态学报,2006,29(8):4127-4137.
[5]闫瑞瑞,闫玉春,辛晓平,等.不同放牧强度下草甸草原土壤微生物和酶活性研究.生态环境学报,2011,20(2):259-265.
[6]张成霞,南志标.放牧对草地土壤理化特性影响的研究进展.草业科学,2010,27(11):131-136.
[7]赵帅,张静妮,赖欣,等.放牧与围栏内蒙古针茅草原土壤微生物生物量碳、氮变化及微生物群落结构PLFA分析.农业环境科学学报,2011,30(6):1126-1134.
[13]胡雷,王长庭,王根绪,等.三江源不同退化演替阶段高寒草甸土壤酶活性和微生物群落结构的变化.草业学报,2014,23(3):8-19.
[16]鲍士旦.土壤农化分析(第3版)[M].北京:中国农业出版社,2000:28-49.
[17]夏北成.植被对土壤微生物群落结构的影响.应用生态学报,1998,9(3):296-300.
[18]胡亚林,汪思龙,颜绍馗.影响土壤微生物活性与群落结构因素研究进展.土壤通报,2006,37(1):170-176.
[19]王光华,金剑,徐美娜,等.植被、土壤及土壤管理对土壤微生物群落结构的影响.生态学杂志,2006,25(5):550-556.
[20]王启兰,王长庭,杜岩功,等.放牧对高寒草甸草原土壤微生物量碳的影响及其与土壤环境的关系.草业学报,2008,17(2):39-46.
[27]张婷,翁月,姚凤娇,等.放牧强度对草甸植物小叶章及土壤化学计量比的影响.草业学报,2014,23(2):20-28.
