喀斯特山地植被恢复过程主要种生态位梯度变化
2014-12-27欧芷阳庞世龙何琴飞黄小荣彭玉华
欧芷阳,庞世龙,何琴飞,黄小荣,彭玉华
(1. 广西壮族自治区林业科学研究院;2. 国家林业局 中南速生材繁育实验室;3. 广西优良用材林资源培育重点实验室,广西 南宁 530002)
喀斯特山地植被恢复过程主要种生态位梯度变化
欧芷阳1,2,3,庞世龙1,2,3,何琴飞1,2,3,黄小荣1,2,3,彭玉华1,2,3
(1. 广西壮族自治区林业科学研究院;2. 国家林业局 中南速生材繁育实验室;3. 广西优良用材林资源培育重点实验室,广西 南宁 530002)
在群落调查的基础上,通过生态位宽度与生态位重叠指数,结合典范对应分析(CCA),研究了广西平果喀斯特山地主要植物种群在植被恢复过程的4个群落中的生态位梯度变化。结果表明:(1)各主要种群的生态位宽度在不同群落中随种类组成的变化而发生改变,这种改变反映了喀斯特山地植被的恢复效果;(2)主要种群生态位宽度的典范对应分析表明,岩石裸露率与4个群落第一排序轴的相关性都比较强,各种群的生态位宽度在不同资源位上表现不一;(3)各群落优势种群间的生态位重叠程度不高,种群间资源利用差异显著、利用性竞争不强,群落处于演替阶段,群落物种组成不稳定。种群间生态位重叠值的大小与种群生态位宽度间不呈明显相关性。
山地植被恢复;喀斯特山地;生态位宽度;生态位重叠;典范对应分析;广西平果县
生态位( Niche)是生态学理论中一个应用广泛且定义繁多的重要概念,首先由Grinnell[1]引入生态学领域,用于描述一种生物在群落中的地位和作用;至2002年Shea和Chesson[2]重新将生态位定义为物种对每个生态位空间点的反应和效应。生态位是生态学最重要的基础理论之一,生态位的研究已经渗透到许多研究领域。在植物群落研究中,生态位在阐明植物与环境的关系、种间关系以及群落结构方面有着广泛的应用[3-5]。
我国喀斯特地貌主要集中于滇、黔、桂三省内。在热带、亚热带湿润—半湿润气候条件和岩溶极其发育的自然背景下,人类不合理活动导致该区域地表植被退化、土壤严重侵蚀、基岩大面积裸露的土地退化现象[6],部分地段甚至完全石漠化[7]。植被恢复是石漠化治理的主要手段之一,探讨植被自然恢复过程中优势种群的生态位梯度变化,对石漠化地区植被恢复和生态重建措施的制定和实施非常重要。因此,喀斯特山地植物群落优势种群的生态位引起了国内相关学者的关注[8-10]。平果县位于广西西南部右江河畔,该县中部岩溶地貌发育典型,为广西岩溶石山分布较多的县份之一。平果石山区生态环境退化严重,石漠化程度相当突出[11]。研究人员在平果建立示范基地的基础之上,开展了土壤立地划分与生态恢复[12]、土壤改良[13]及土壤微生物量[14]等方面的研究,还有学者对平果喀斯特山地蚬木种群[15]以及主要植被的凋落物[16]展开了相关研究,但对该区域山地植被优势种群的生态位研究却鲜有报道。本研究在群落学调查基础上,对平果县喀斯特山地植物群落主要种的生态位进行研究,分析这些物种在植被恢复过程中的生态位梯度,深入了解种群在不同群落中的功能地位及其环境资源利用状况,研究可为有效进行石漠化治理提供理论依据。
1 研究区概况
研究地区在广西平果县(23°12′~ 23°51′N,107°21′~ 107°51′E)典型喀斯特地貌分布区。
平果县位于广西西南部, 全县土地总面积2 385 km2,其中岩溶山地面积占到80% 以上(吕仕洪等,2005)。平果县毗邻右江河谷,属亚热带季风气候区,热量丰富。该县年平均温度19~21.5 ℃,年平均降雨量1 200~1 500 mm,年平均日照时数1 619.4 h,年均蒸发量1 572 mm,平均相对湿度80% 以上,降雨多集中在5月至9月之间[17]。喀斯特山地土壤以棕色和黑色石灰土为主,土壤特征表现为土层薄且保水保肥能力差。调查区域植被以灌丛、灌木、疏林地占较大比例,村屯后山保留着原生植被遭受破坏后残留下来的次生林地,在坡度较大的山体上部,常分布着灌木物种东矮化成灌木型的乔木树种。乔木种类主要以蚬木Excentrodendron hsienmu和仪花Lysidice rhodostegia占优势;灌木主要有黄荆Vitex negundo、红背山麻杆Alchornea trewioides)、灰毛浆果楝Cipadessa cinerascens 等;草本植物主要有莠竹Microstegium nodosum、蔓生莠竹Microstegium vagans、飞机草Chromolaena odoratum等;藤本植物主要有雀梅藤Sageretia thea、飞龙掌血Toddalia asiatica等。
2 研究方法
2.1 样地调查
2012年4月至6月上旬,运用典型取样法对处于不同恢复阶段的植物群落进行调查。乔木林、乔灌过渡林、灌木林调查样方面积为20 m × 20 m,灌丛调查样方面积为10 m ×10 m,共设置44个样方,总调查面积为12 600 m2。每个样方进一步划分为5 m ×5 m的测量单元进行木本植物调查,对样方内树高(height, H)≥1.3 m、胸径(diameter at breast height, DBH)≥1 cm的立木进行每木检尺,记录其种名、高度、胸径、冠幅、枝下高等数值。在每个样方的四个角和中心点处各设一个2 m ×2 m小样方,记录小样方内出现的灌木和草本植物的物种名、株数、平均高度和盖度。同时记录海拔、坡向、坡度、坡位及岩石裸露率等生境因子信息,岩石裸露率(percentage of covered rock, Rock)借鉴宋同清等[18]的方法用网格法测定,即将100 cm× 100 cm的框架分成20 cm × 20 cm网格,在样方内按对角线5点覆盖来测定。
选取25个主要种作为研究对象(表1),分析它们在植被恢复过程中4个不同群落类型中的生态位变化。这4个群落分别是I. 黄荆+灰毛浆果楝+红背山麻杆-莠竹;II. 黄荆+潺稿树Litsea glutinosa-莠竹;III. 仪花-薄皮酒饼簕Atalantia henryi-石山棕Guihaia argyrata;IV. 蚬木-薄皮酒饼簕-石山棕+槲蕨。
2.2 数据处理与分析
Levins生态位宽度计测:

式(1)中,Bi为种i的生态位宽度;Pij=nij/Ni+,代表种i在第j个资源状态下的个体数占该种所有个体数的比例,r为样方数(张金屯,数量生态学)。
Moristia-Horn生态位重叠指数:

式(2)中,Oik为种i和种k的生态位重叠值,k为不同于i的另一种。
建立物种多度矩阵数据表,运用CANOCO 4.5 软件进行典范对应分析 (canonical correspondence analysis, CCA),以分析各群落主要种群与环境因子的关系,采用蒙特卡罗法(Monte Carlo) 进行显著性检验(α=0.05)。

表1 优势物种名及序号Table1 Name and sequence number of dominant species
3 结果与分析
3.1 优势种群的生态位宽度
测度25个主要种在4个群落类型中的生态位宽度值,结果见表2。群落I为灌丛群落,其生境最为干旱贫瘠。该群落灌木层以灰毛浆果楝具有最大的生态位宽度,其次为黄荆。结合物种在群落中的重要值来看,二者为群落建群种,它们对资源的共同需求,必然会导致竞争的存在。群落I的草本层以莠竹表现为最宽的生态位宽度和最大的重要值,表明群落生境有利于对土壤和水分要求不严格的植物物种生存。乔木树种小叶榕和黄牛木在群落I中的生态位宽度值最小,意味着该群落环境对这两个种群并不太适宜,它们在群落中的地位是次要的或暂时的。群落II是为灌木群落,胸径≥2.0 cm的立木以潺稿树的重要值占优势,灌木层以红背山麻杆重要值最大。从生态位宽度来看,群落II乔木层以黄牛木的生态位宽度最大,灌木层以岩柿的生态位宽度最大,草本层以石山棕的生态位最大、莠竹的生态位宽度有所下降。分析结果表明随着植被恢复,群落土壤水分和郁闭度有所增加、群落生境得到改善,有利于对土壤和水分要求较为严格的物种生存,群落物种组成发生改变。

表2 优势种群在群落中的生态位宽度值Table2 Niche breadth values of dominant species in communities
群落III是以仪花占优势的次生林,该群落乔木层以圆叶乌桕种群的生态位宽度指数最大,灌木层仍以岩柿的生态位宽度值最大,草本层以石山棕表现为最宽的生态位宽度(表2)和最大的重要值,群落生境的改善更有利于该物种的生存。乔木层优势种仪花和伴生种鱼骨木的生态位宽度值较为接近且低于圆叶乌桕,意味着二者在资源利用方面并不占据优势,而它们对养分的共同需要又导致了激烈的竞争。同时,乔木层与灌木层间的物种存在竞争,群落稳定性差。群落IV是以蚬木为建群种的次生常绿阔叶林,其乔木层以圆叶乌桕的生态位宽度最大、蚬木其次,灌木层仍以岩柿的生态位宽度最大,草本层以重要值较高的槲蕨表现为最大的生态位宽度,表明该群落环境在一定程度上适宜对土壤和水分要求更为严格的物种生存。结合物种重要值来看,蚬木在构建群落的内部生境和决定群落的组成结构方面占绝对优势,但在资源利用方面受到圆叶乌桕的竞争。
3.2 主要种群生态宽度的典范对应分析
平果喀斯特山地森林群落优势种群的生态位宽度的典范对应分析(CCA)见图1~图4。图中空心三角形代表优势种群,环境因子用箭头表示,空心三角形与箭头连线的相对位置解释了优势种群与环境因子的相关性。
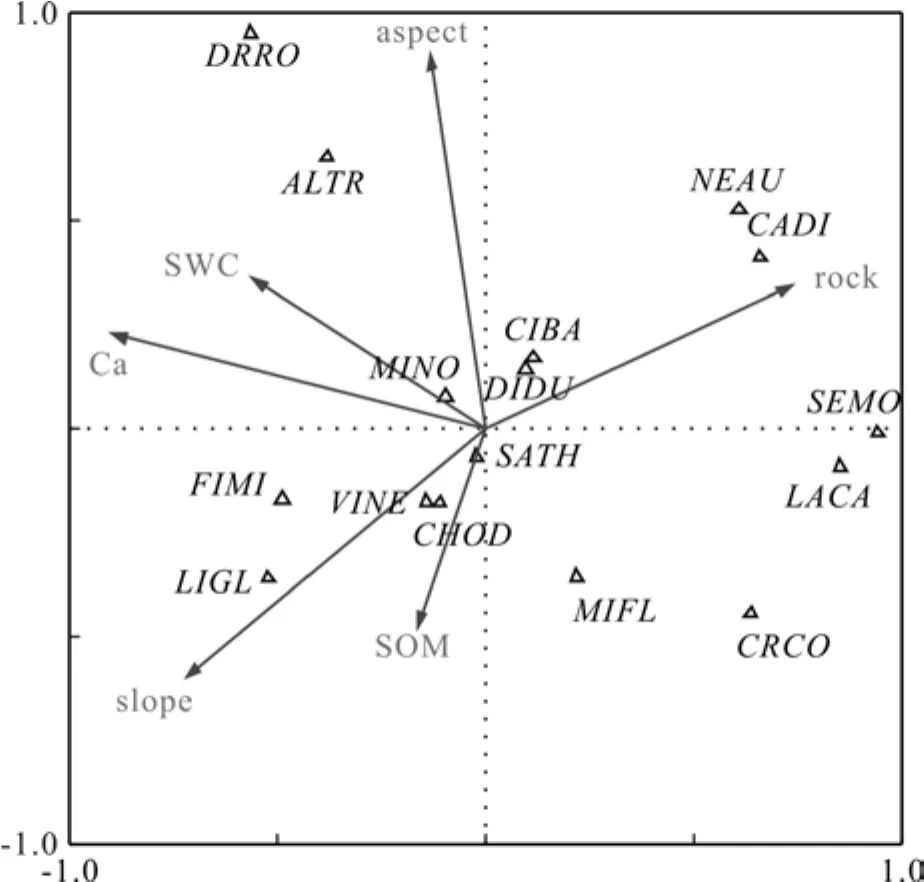
图1 群落-I 优势种群的CCA排序Fig.1 Two-dimensional CCA ordination of dominant species in community-I

图2 群落-II优势种群的CCA排序Fig.2 Two-dimensional CCA ordination of dominant species in community-II

图3 群落III优势种群的CCA排序Fig.3 Two-dimensional CCA ordination of dominant species in community-III

图4 群落IV优势种群的CCA排序Fig.4 Two-dimensional CCA ordination of dominant species in community-IV
群落I典范对应分析3个排序轴的特征根分析为0.302、0.128和0.046,群落与环境因子3个排序轴的相关系数各为0.996、0.983和0.835,蒙特卡罗检验达到显著水平(P<0.05),排序效果理想。6个环境因子与第一、第二排序轴均有不同程度的相关性。其中,土壤交换性钙、岩石裸露率与第一排序轴相关性较强,相关系数分别为-0.894 1和0.733 3;坡向与第二排序轴的正相关性较强,相关系数为0.880 9。群落I中,雀梅藤(2.99)在岩石裸露率资源位上的生态位宽度最大,飞机草(2.95)其次;黄荆(2.47)和灰毛浆果楝(2.46)在土壤含水量上的生态位宽度最大,这两种植物在干旱土壤环境下的适应性较强。
群落II典范对应分析前3个排序轴的特征根分析为0.320、0.219、0.120,物种与环境因子3个排序轴的相关系数各为0.957、0.980和0.928。岩石裸露率与第一排序轴有较强的负相关(r=-0.731 6),交换性钙与第二排序轴有较强的正相关(r= 0.711 5)。群落II中,优势种群红背山麻杆(2.95)在岩石裸露率上的生态位宽度最大,黄荆(2.99)在坡度上的生态位宽度最大,小叶榕在坡向和土壤有机质(2.00)资源位上的生态位宽度都是最大的,黄牛木(2.48)和雀梅藤(2.86)分别在土壤交换性钙和含水量上的生态位宽度最大。
群落III典范对应分析前3个排序轴的特征根分析为0.808、0.417、0.227,物种与环境因子3个排序轴的相关系数各为0.981、0.987和0.948,蒙特卡罗检验达到显著水平(P<0.05)。群落III中,岩柿(2.75)在岩石裸露率资源位上的生态位宽度最大,雀梅藤(2.80)和黄荆(2.50)分别在坡度和坡向上的生态位宽度最大,圆叶乌桕(3.24)在土壤有机质资源位上的生态位宽度最大,小叶榕(3.31)和红背山麻杆(3.02)分别在在土壤交换性钙和含水量上的生态位宽度最大。
群落IV典范对应分析前3个排序轴的特征根分析为0.474、0.310、0.103,物种与环境因子3个排序轴的相关系数各为0.983、0.822和0.688,第一典范轴的蒙特卡罗检验达到显著水平(P<0.05)。群落IV中,小栾树和江南卷柏在岩石裸露率资源位上的生态位宽度最大(2.00),肾蕨在坡度(2.78)、潺稿树在坡度(2.78)和坡向(3.53)上的生态位宽度均达到最大,圆叶乌桕在土壤有机质(3.28)和含水量(2.95)资源位上的生态位宽度都是最大,槲蕨在土壤交换性钙(3.58)上的生态位宽度最大。
3.3 物种生态位重叠分析
表3为优势种在各群落中生态位重叠值的分级表,表中显示各种对间的生态位重叠值普遍偏低。群落I中16个主要种构成120个物种对,超过80.0%种对间的生态位重叠值低于0.5,有15个种对间未形成生态位重叠。群落II中23个主要种构成了253个物种对, 78.2%种对间的生态位重叠值低于0.5,有28个物种对间未形成生态位重叠。群落III中24个主要种构成了276个物种对,74.6%种对间的生态位重叠值低于0.5,共有15个种对间未形成生态位重叠。群落IV中23个主要种构成了253个物种对,生态位重叠值低于0.5的种对达79.0%,其中有29个种对间未形成生态位重叠。由生态位重叠值>0.8的种对数可以看出,从灌丛群落到次生乔木林群落,随植被恢复时间加长,高生态位重叠的种对数基本呈增加趋势。生态位宽度值较大的物种与其他种间的生态位重叠值一般较高,表明群落中主要种群对生态因子的适应性互补性方面趋同性增强[3]。但群落I中具有较大生态位宽度种群之间的生态位重叠值并不高,这可能是由于物种本身生态学特性不同、对资源段位要求不完全一致,从而导致种间生态位重叠不高。

表3 优势种群在各群落中的生态位重叠值分级Table3 Grading standards of niche overlap of dominant species in communities
4 结论与讨论
生态位能很好地反映种群的生态适应性、资源利用和竞争能力,物种生态位越大,其对环境的适应能力越强,对资源的利用越充分,在竞争中就占有优势[19]。本研究25个主要种在植被恢复不同阶段的生态位宽度并不一致。群落I为灌丛群落,乔木树种在资源利用方面不占优势,其生态位宽度小。该群落灌木层以灰毛浆果楝和黄荆的生态位宽度指数较大,草本层以莠竹的最大;群落II乔木层以黄牛木的生态位宽度最大,灌、草层分别以岩柿和石山棕的生态位宽度最大;群落III乔木层以圆叶乌桕种群的生态位宽度指数最大,灌、草层仍以岩柿和石山棕的生态位宽度最大;群落IV乔、灌层也分别以圆叶乌桕和岩柿的生态位宽度最大,草本层以重要值较高的槲蕨表现为最大的生态位宽度。分析结果显示,在植被演替的早期阶段,重要值较大的种群一般具有较宽的生态位。黄荆、灰毛浆果楝和红背山麻杆能适应喀斯特贫瘠、干旱的生境,分布范围较广,是桂西南喀斯特山地灌丛群落的优势种,具有较宽的生态位,其他研究也具有相似的结论[10]。随着植被恢复、群落生境的改善,对生境要求更为严格的物种出现,乔木树种对喀斯特环境资源利用的尺度增大、其适应能力进一步增强,群落的物种组成发生了变化。但在由乔灌阶段向乔木林恢复的进程中,乔木层优势种的生态位宽度小,在资源利用方面不占优势,与群落共有种间存在较强的竞争,与灌木层物种间也存在较强的竞争,群落结构不稳定。蚬木作为调查区域次生林的优势种,在构建群落的内部生境和决定群落的组成结构方面占绝对优势,其在资源利用方面受到圆叶乌桕的竞争。此外,岩柿作为喀斯特生境的泛化种,其资源利用能力强、对喀斯特环境资源的利用尺度大,因而生态位宽度值在植被不同恢复阶段都比较大。
生态位重叠意味着两个物种在其与生太因子联系上具有相似性[20],或2个种对环境资源有互补的要求,体现了物种对同等级资源的利用程度及空间配置关系。存在生态位重叠的两个物种间,其关系较为复杂。在资源丰富的群落中,生态位重叠值大的物种共享同一资源,二者竞争并不激烈;而在资源受到限制的生境中,物种间较高的重叠值可能导致强烈种间竞争。另外,当一个种为另一个种的生存提供有利条件时,生态位重叠值较大不仅不是竞争,而是一种互惠[21]。本研究群落I中生态位重叠值大于0.8的物种对有5对,占总种群数的4.2%; 群落II中生态位重叠值大于0.8的有19对,占总种群数的7.5%; 群落III中生态位重叠值大于0.8的共有26对,占总种群数的9.4%,群落IV生态位重叠值大于0.8的物种对比例有所下降,占总种群数的6.3%。数据表明,各群落优势种群间的生态位重叠程度不高,种群间利用资源的差异显著、利用性竞争不强,群落处于演替阶段,群落的组成类型不稳定。有研究表明种群的生态位宽度越大,与其他种群的生态位重叠的机会就越高,生态位宽度较小的物种与其他种群间的生态位重叠的机会较小[3,8,22],但也有研究结果显示种对之间的生态位重叠值大小与其生态位宽度大小不具明显的规律性[9,23-26]。本研究群落I中生态位宽度较大的黄荆、灰毛浆果楝和红背山麻杆与其他物种间的生态位重叠值均比较低,这与物种本身的生态学特性及它们对资源段位的需求不完全一致有关。另三个群落生态位宽度较大的物种与其他种群的生态位重叠值相对较高,表明随着演替进程,群落中优势种群对资源的利用能力相似,种群间对资源的利用性竞争可能加强。
[1] Grinnell J. The niche-relationship of the Califomia thrasher[J].Auk, 1917, 34: 427-433.
[2] Sheak K, Chesson P. Communily eeology theory as a framework for biological invasions[J]. Trends in Ecology & Evaluation,2002, 17 (4): 170-176.
[3] 郭逍宇, 张金屯, 宫辉力, 等. 安太堡矿区植被恢复过程主要种生态位梯度变化研究[J]. 西北植物学报, 2004, 24(12):2329-2334.
[4] 张 璐, 苏志尧, 陈北光, 等. 广东石坑崆森林群落优势种群生态位宽度沿海拔梯度的变化[J]. 林业科学研究, 2007,20(5): 598 –603.
[5] 伊力塔, 豪树奇, 韩海荣, 等. 灵空山辽东栎林种群生态位特征[J]. 浙江农林大学学报, 2012, 29(1): 46-51.
[6] 卢 峰. 广西岩溶土地现状与石漠化治理模式探析[J]. 广西林业科学, 2012, 41(2): 183-185.
[7] 刘 方, 王世杰, 刘元生, 等. 喀斯特石漠化过程土壤质量变化及生态环境影响评价[J]. 生态学报, 2005, 25(3): 639-644.
[8] 张忠华, 梁士楚, 胡 刚. 桂林岩溶石山阴香群落主要种群生态位研究[J]. 林业科学研究, 2009, 22(1): 63-68.
[9] 李安定, 李苇洁, 彭 熙, 等. 贵州喀斯特区野生葛藤群落主要种群生态位[J]. 浙江农林大学学报, 2012, 29(4): 491-497.
[10] 马姜明, 占婷婷, 莫祖英, 等. 漓江流域岩溶区檵木群落不同恢复阶段主要共有种生态位变化[J]. 西北植物学报, 2012,32(12): 2530-2536.
[11] 李先琨, 吕仕洪, 蒋忠诚, 等. 喀斯特峰丛区复合农林系统优化与植被恢复试验[J]. 自然资源学报, 2005, 20(1): 92-98.
[12] 吕仕洪, 陆树华, 李先琨, 等. 广西平果县石漠化地区产地划分与生态恢复试验初报. 中国岩溶, 2005, 24(3): 196–201.
[13] 罗为群, 蒋忠诚, 覃小群. 岩溶石山区石灰土改良方法及对比试验研究——以广西平果县龙何屯为例[J]. 地球与环境,2008, 36(1): 87-92.
[14] 徐广平,何成新,张德楠,等. 桂西南岩溶山地不同土地利用方式土壤微生物量及其活性特征[J]. 广西植物, 2013, 33(3):331-337.
[15] 袁铁象, 欧芷阳, 苏志尧, 等. 桂西南喀斯特山地蚬木野生种群结构与动态[J]. 中南林业科技大学学报, 2013, 33(3): 1-8.
[16] 彭玉华, 欧芷阳, 曹艳云, 等. 桂西南喀斯特山地主要植被类型凋落物累积量及其持水特性[J]. 中南林业科技大学学报,2013, 33(2): 81-85.
[17] 于明含, 孙保平. 广西平果县退耕还林土壤环境效应及生态效益价值估算[J]. 浙江农业学报, 2012, 24( 1): 114 -119.
[18] 宋同清, 彭晚霞, 曾馥平, 等. 木论喀斯特峰丛洼地森林群落空间格局及环境解释[J]. 植物生态学报, 2010,34(3):298-308.
[19] 刘晓宁, 张钦弟, 闫 明, 等. 五鹿山国家级自然保护区油松群落优势种生态位研究[J]. 植物研究, 2012, 32(6): 712-716.
[20] 王 刚.关于生态位定义的探讨及生态位重叠计测公式改进的研究[J]. 生态学报, 1984, 4(2): 119-126.
[21] 王仁忠. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究[J]. 植物生态学报, 1997, 21(4): 304-311.
[22] 程红梅, 沈年华, 汤庚国, 等. 大蜀山短毛椴落叶阔叶林优势种群生态位特征[J]. 南京林业大学学报(自然科学版), 2009,33(1): 146-150.
[23] 林伟强, 贾小容, 陈北光, 等. 广州帽峰山次生林主要种群生态位宽度与重叠研究[J]. 华南农业大学学报,2006,27(1):84-87.
[24] 郑世群, 刘金福, 黄志森, 等. 戴云山罗浮栲林主要乔木树种营养生态位研究. 热带亚热带植物学报, 2012, 20(2): 177-183.
[25] 周庆贤, 赵菊鹏, 李 恒, 等. 青花梨在广州北部山区引种初探[J]. 生物灾害科学, 2013, 36(3): 301-303.
[26] 许 仕, 谢冬明, 金国花. 赣江源自然保护区森林生态系统服务功能价值初步评价[J]. 生物灾害科学, 2013,36(4):383-388.
Gradient changes of dominant species’ niche in the process of vegetation restoration in karst mountain area of Pingguo county, Southwest Guangxi province
OU Zhi-yang1,2,3, PANG Shi-long1,2,3, HE Qin-fei1,2,3, HUANG Xiao-rong1,2,3, PENG Yu-hua1,2,3
(1.Guangxi Forestry Research Institute; 2.Central South Key Lab. of Fast-growing Tree Cultivation Attached to China Forestry Ministry;3.Guangxi Key Lab. of Superior Timber Trees Cultivation. Nanning 530002, Guangxi, China)
Based on fi eld survey in the karst mountains in Pingguo county, southwest Guangxi province, by using niche breadth, niche overlap and canonical correspondence analysis (CCA), the ecological gradient changes of 25 main species’ niche in four communities were investigated in the process of vegetation restoration. The results show that (1) the niche breadth of main populations varied with species composition of community, which ref l ected the effects of vegetation restoration in karst mountains in the study area; (2)the fi ndings of CCA indicate that the bare rock ratio highly correlated with the fi rst sorting axes of the four communities, the main populations’ niche breadth at different resource states were different respectively; (3) the niche overlaps of 25 main populations in the four communities were all not higher, which revealed that the resources utilization of the main populations was signif i cant difference, and the resources utilizing competition was not strong. The secondary communities stayed in the stages of succession, and the community composition was not stable. The intra-population niche overlap was not obviously related to the niche breadth.
vegetation restoration in hilly area; karst mountain area; niche breadth; niche overlap; canonical correspondence analysis;Pingguo county of Guangxi
S718.5
A
1673-923X(2014)12-0096-06
2014-04-04
广西优良用材林资源培育重点实验室自主课题(12B0401)
欧芷阳(1975-),女,工程师,博士,主要从事森林生态学研究工作;E-mail:ozhiyang@126.com
[本文编校:文凤鸣]
