不同林龄尾巨桉人工林的生物量及其分配特征
2014-12-22付威波彭晚霞宋同清曾馥平温远光徐慧芳
付威波,彭晚霞 ,宋同清,曾馥平,杜 虎,温远光,徐慧芳,4
(1.中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,长沙 410125;2.中国科学院环江喀斯特生态系统观测研究站环江 547100;
3.广西大学林学院,南宁 530004;4.江西农业大学,南昌 330045)
受人类活动的影响,全球碳循环发生了显著改变,大气CO2等温室气体浓度持续大幅度的增加导致温室效应增强、气候变暖、大气环流发生变化、区域性气候变化加剧等一系列的生态环境问题,减排CO2的呼声日益高涨[1]。森林生态系统是世界上除海洋之外最大的碳库,其碳储量约为1 146 PgC,占全球陆地总碳储量的46%[2],维持着全球植被碳库的86%和土壤碳库的73%[3],对全球碳平衡起着十分重要的作用。但森林生态系统向大气中的碳排放仍然是个很不确定的数字[4],而造成此不确定的最主要原因就在于对森林生物量的估算[5],获得更为准确的森林生物量的数据是进一步了解森林在全球碳循环中作用的关键[6]。生物量是指一个有机体或群落在一定时间内积累的有机质总量。作为森林生态系统的特征数据,生物量是研究森林生态系统结构与功能的基础,对深入研究森林生态系统生物地球化学循环、碳汇功能、评价人工林生态系统生产力与环境因子之间的关系具有重要的科学价值[7]。自20世纪60年代以来,国际上许多学者陆续对森林的生物量进行了广泛而深入的研究[8-10],我国自20世纪70年代末开始展开森林生物量的研究。
桉树约有900多种,绝大部分产于澳大利亚及邻近岛屿,因干形好、生长快、产量高、用途广,全球已有上百个国家和地区引种和广泛栽培[11]。我国南方桉树人工林发展速度快、规模大,其面积已突破200×104hm2[12],但同时也出现了桉树人工林地力衰退、林分产量下降的现象,维持桉树人工林长期稳定的可持续发展已成为国内外关注的焦点。有关学者对桉树人工林的生物量进行了研究[11,13],但这些研究都局限于某一年龄桉树生物量,难以对不同林龄桉树人工林生物量进行准确的估测,也缺乏不同生长发育阶段生物量的组成及分布规律研究。本文基于2011年底广西东南部5个不同年龄阶段的尾巨桉人工林生物量的实测调查,探讨了尾巨桉人工林生物量组成、分配以及各器官生物量随年龄的变化规律,为进一步开展桉树人工林科学发展及碳循环研究提供科学依据。
1 自然概况
研究区位于广西东南部桉树主产区(22°38'—24°24'N,107°48'—111°33'E)。据 2009 年第八次森林资源清查结果统计(广西林业勘测设计院提供),广西桉树面积达1712968 hm2,蓄积67626071 m3,主要分布在广西东南部,该区域属亚热带季风气候区,具有热带向亚热带过度的特点,年平均气温21.5—22℃,1月平均气温12.8—13.5℃,7月平均气温27.9—28.3℃,年积温7190—8030℃。年降雨量1 300—1 800 mm,主要集中在4—9月,占全年降雨量的80%,年总日照1 600—1 800 h,年蒸发量1 600 mm,相对湿度74.8%。试验地土壤为砂页岩发育的赤红壤。
2 研究方法
2.1 样方选择与建立
参照《IPCC优良做法指南》对系统随机抽样的建议和广西桉树人工林的分布特征,基于第八次森林资源清查数据,计算桉树人工林幼龄林(1a)、中龄林(2a)、近熟林(3a)、成熟林(5a)、过熟林(8a)5个不同林龄在广西各县(市)的面积、蓄积综合权重,选择权重最大的县(市),在广西东南桉树主产区建立5个不同林龄尾巨桉人工林样地,样地大小为1 000 m2(50 m×20 m),每个林龄设置3个重复,共计15个样地。将每块样地进一步划分为10个10 m×10 m的样方,对样方内测量胸径(D)≥2 cm尾巨桉的胸径、树高、冠幅和坐标进行每木调查,并挂牌标号,便于后期复查。各样地的基本信息见表1。

表1 不同林龄尾巨桉人工林样地基本特征Table 1 Sites description of Eucalyptus urophylla×E.grandis p lantation at different stand ages
2.2 生物量获取2.2.1 乔木层
在每木调查的基础上,以2 cm为径阶,根据中央径阶多、两端逐次少的原则,在样地外选择不同林龄和径阶的尾巨桉共18株作为样木,进行生物量的测定。测定其基径、胸径、树高和冠幅并按2 m区分段锯断称量,在树干基部、胸径、中部及顶端分别锯取一个圆盘测定含水量;并分别称取枝条和叶片的鲜质量;根的生物量采用全挖法,并分小根(<2 cm)、中根(2—5 cm)、大根(>5 cm)3组分别称重;然后对各部分器官分别取样300 g带回实验室,将样品在105℃烘箱内杀青2 h,调至75℃下烘干至恒重,求出各器官干鲜质量之比,由此换算出样木各器官的干质量及总干质量。利用样木各器官(干、枝、叶、根)的干重和总干重建立各器官及总生物量与样木胸径(D)的幂回归方程(W=aDb)或指数方程(W=aebD)。根据样地每木调查的结果及相应的回归方程计算乔木层各个体的生物量,并由此获得乔木层的总生物量。2.2.2 灌木层、草本层和地上凋落物
按“品”字型在样地1、5、8号小样方各设置1个2 m×2 m的样方,调查所有灌木种类、株丛数、高度、地径、覆盖度,在每个2 m×2 m的样方内取1各1 m×1 m的小样框,调查草本种类、株丛数、平均高度、覆盖度。为了不破坏复查样地,在相距各样框5 m的范围内选择1个与灌木调查样框植被类型、立地条件等基本一致的样框按全收获法收获样框内所有灌木的枝、叶、花果、根,测定各器官生物量,并以3个样框的平均值来推算样地中灌木层的总生物量。按同样方法在1 m×1 m小样框内获取草本层植物各器官、凋落物生物量及总生物量。
2.2.3 数据处理
所有数据及多重比较分析、回归方程模型建立均在Excel-2003和SPSS 16.0中进行处理。
3 结果与分析
3.1 乔木层生物量回归模型的建立
大量的森林群落生物量研究表明乔木各器官及总生物量与测树因子间普遍存在着相关关系,这种相关关系可以用数学模型进行拟合,其表达式为:W=axb,式中,W为各器官的生物量。自变量x可选用胸径(D)、树高(H)或胸径平方乘以树高(D2H)等测树因子,a、b为方程中的待估参数。该方程能较真实地反应树木生物量随胸径(D)和树高(H)的变化趋势[14-16]。因树高估测误差较大,本研究生物量预测方程采用以树木胸径(D)为单变量的非线性幂函数或指数函数方程,根据相关系数和显著性检验,尾巨桉叶生物量的回归模型为W=a ebD2,其余器官的回归模型均为W=aDb(表2),经统计学检验各器官干、枝、叶、根和总生物量数学模型的相关系数在0.873—0.989之间,t检验均达到极显著水平(P<0.01),表明模型可以对尾巨桉林的生物量进行估测。

表2 尾巨桉人工林生物量估算模型Table 2 Regression models of biomass for Eucalyptus urophylla×E.grandis
3.2 不同林龄尾巨桉人工林乔木层生物量及其分配
尾巨桉人工林生物量与其林龄密切相关,总生物量随着林龄的增加迅速增加(表3),1,2,3,5年生和8年生尾巨桉人工林生物量分别为6.84,42.11,54.12,94.15 和130.86 t/hm2,除 2、3 年生达到显著水平外,其它均达到极显著水平。但不同年龄阶段生物量的增加幅度不同,其中以1—2年生林分生物量增加最为迅速,1年间生物量增加了35.26 t/hm2,3—5年间林分生物量增加也较快,平均年增长量为20.01 t/hm2,2—3年生和 5—8年生林分平均年增长量分别为12.01和12.23 t/hm2,相对较慢。这种现象说明尾巨桉生物量效应在不同的林分发育阶段有不同的表现,即存在着“年龄效应”或发育阶段效应,具有两个生长高峰,即呈升—降—升—降的发展趋势,这主要是受尾巨桉林分保存率的影响。2—3a间林分保存率下降后逐渐趋于稳定状态,从而产生了第2个生长高峰。
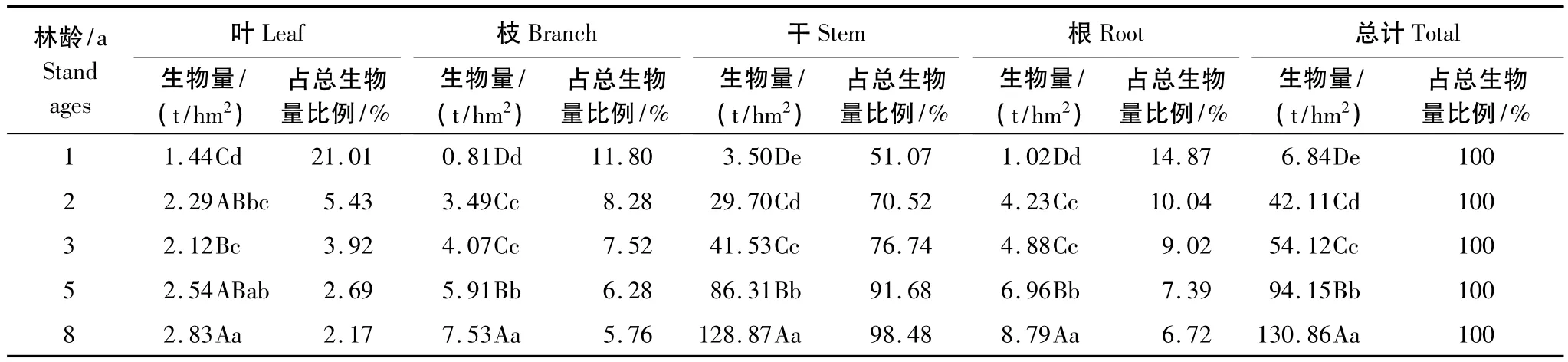
表3 不同林龄尾巨桉人工林乔木层生物量器官分配Table 3 Biomass allocation in arbor layer at different stand ages for E.urophylla×E.grandis plantations
通过分析不同林龄尾巨桉各器官生物量数据可知(表3),各器官生物量除3年生叶生物量略有下降外,其余各器官生物量均随着林龄的增大而增大。但各器官生物量的增加幅度不同,其占总生物量的比例也有所变化,不同林龄尾巨桉生物量在各器官中的分配比例大小略有不同,1年生林分为:干>叶>根>枝,2—8年生为:干>根>枝>叶,叶、根和枝在不同生长阶段分配比率大小不同,表明了尾巨桉叶、根和枝在不同发育阶段其生长上的差异,1年生尾巨桉叶的生长大于根和枝,2年生后根的生长则大于枝和叶。
随着林龄的增加各器官生物量的分配体现一定的规律性,在各个年龄阶段均以树干的生物量最大,占整个生物量的51.07%—98.48%,并随着年龄的增加,树干生物量所占比例逐渐增大,根、枝、叶占总生物量的比例与其他树种不同,均表现出了随年龄增加而下降的趋势。表明随着尾巨桉年龄的生长越来越多的干物质储存在树干中,干材随着年龄的增长是一个净积累的过程,根、枝、叶在积累的同时却有部分的衰老组织在脱落,具有比较强烈的更新换代过程。
3.3 不同林龄尾巨桉人工林灌木层生物量及其分配
尾巨桉人工林由1年生发展至2年生林分郁闭度增加,灌木层生物量急剧减少(表4),至3年生时由于林分保存率下降,灌木层生物量急剧增加达到最大值,伴随林分保存率的相对稳定和林分不断生长,灌木层生物量又呈下降趋势。灌木层各器官的分配比例在不同的发育阶段有所不同,但规律性不强,与乔木层相比相对稳定,除2年生为枝>叶>根外,其余为枝>根>叶,均以枝的比例最大,达37.89%—56.79%。

表4 不同林龄尾巨桉人工林灌木层生物量器官分配Table 4 Biomass allocation in shrub layer at different stand ages for E.urophylla×E.grandis plantations
3.4 不同林龄尾巨桉人工林草本层及凋落物生物量
不同林龄尾巨桉草本层生物量不同(表5),大小顺序为:5a>1a>3a>8a>2a。1 年生尾巨桉人工林林分较小、空间大、阳光充分,草本层生物量大,2年生尾巨桉林逐渐密闭,草本层生物量最小,随着尾巨桉林的增长及林间空隙的增大草本层生物量逐渐增大,8年生尾巨桉林过度密闭草本层生物量又急剧下降,除8年生地下生物量>地上外,其余的均是地上>地下。随着尾巨桉林龄的增加凋落物逐渐增加(表5),至5年生尾巨桉人工林凋落物的生物量最高,达6.51 t/hm2,至8年生尾巨桉林因枝、叶的过分脱落和更新换代能力的急剧下降,凋落物生物量反而减小。

表5 不同林龄尾巨桉人工林草本层、凋落物层生物量分配Table 5 Biomass allocation in herb and litter at different stand ages for E.urophylla×E.grandis p lantations
3.5 不同林龄尾巨桉人工林总生物量组成及其分配
由表6可知,尾巨桉人工林总生物变化规律与乔木层一致,随着林龄的增加而增加,除2年生和3年生差异显著外,其它均达到极显著水平。其中活体植物的贡献达85.60%—97.61%,地上凋落物的总量占2.39%—14.40%。各层生物量所占比例总体趋势为乔木层>凋落物层>草本层>灌木层,以乔木层占绝对优势,所占比例为54.80%—91.56%,除3年生外随着林龄的增加逐渐增加,其次为地上凋落物所占比例为2.39%—14.40%,且随林龄的增加逐渐下降,草本层生物量所占比例除1年生大于凋落物外其余均很小,且除2年生外均呈下降趋势,灌木层生物量所占比例最小,为1.02%—6.47%,且除2年生外随林龄的增加也逐渐减小。
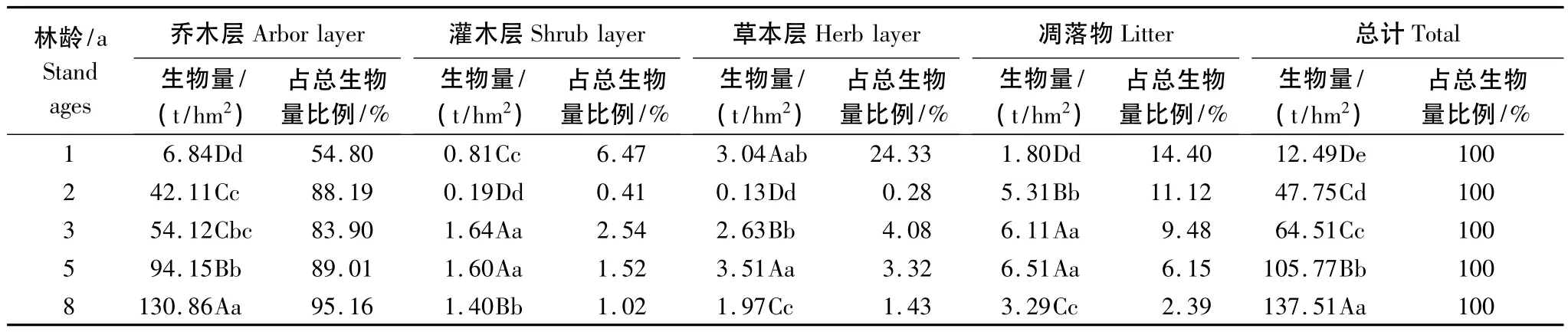
表6 不同林龄尾巨桉人工林各层次生物量分配Table 6 Biomass allocation in different layers at different stand ages for E.urophylla×E.grandis p lantations
2.6 尾巨桉人工林生物量优化增长模型
为了了解尾巨桉人工林各器官、各层次及总生物量随林龄增长的动态变化,以林龄(a)为自变量(X),以乔木层不同器官生物量、乔木层、灌木层、草本层、凋落物各层及总生物量(t/hm2)为因变量(Y),采用多种线性与非线性模型进行模拟,并选用相关系数最高者作为优化回归模型,得出如表7所示的尾巨桉人工林乔木层各器官、各层次及总生物量的增长模型。t检验表明,灌木层和草本层因人为抚育措施的破坏,其生物量的动态模型达不到显著水平,其余各器官、各层次和总生物量均达到了显著或极显著水平,能很好地反映各器官和整个系统的生物量随时间变化的动态过程。

表7 尾巨桉人工林乔木层不同器官、各层及总生物量的增长模型Table 7 Grow th models of different organs in arbor layer,shrub,herb,litter,and total biomass
4 讨论和结论
4.1 讨论
4.1.1 生物量模型的选择
森林生物量的获得可以通过直接或间接两种途径:前者是将一定面积内的树木皆伐以获取其生物量,准确度高,因其对环境的破坏性大,对森林的主要产品干材的损耗太多,很少使用;后者是确定生物量的主要方法,也叫生物量模型法,根据尺度不同可以分为单木生物量模型和大尺度森林生物量模型,单木生物量模型是通过样木观测建立树木的相对生长方程进行生物量的估测[16-17],大尺度森林生物量模型利用多种遥感信息参数进行模拟,许多学者利用多种生物量模型来估计林木的生物量[18-19]。目前相对生长方程的拟合多采用对数形式,常用的模型变量有胸径(D)、树高(H)、密度(S)。常见的预测模型包括以胸径为单变量的模型、以胸径和树高为变量的模型以及以胸径、高度、密度为变量的模型。但在森林群落调查中,各种参数获得的准确性并不一致。胸径在绝大多数的调查中可以较准确的获得,而高度通常会被测量,但其测量均存在着较大的误差[6,20]。且生物量与胸径之间存在着很好的相关关系,因此在森林的生物量预测中以胸径为单变量的模型被广泛运用[5,21-22]。本研究证明以胸径为单变量的尾巨桉人工林生物量数学模型具有较高的精度,各器官及总生物量数学模型的相关系数均达到极显著水平,能够对不同林龄尾巨桉人工林各组分、各层次及总生物量进行有效预测。
4.1.2 尾巨桉人工林与其他森林类型生物量的比较
森林生物量与许多生物学因素和非生物学因素密切相关,如区域的水热条件、土壤条件以及森林的
类型、年龄、优势种的组成,活立木密度等[6,23],本研究表明,8年生尾巨桉生物量为137.51 t/hm2,与其林龄密切相关,乔木层各器官凋落物和总生物量拟合的模型均能很好的反映随林龄变化的动态过程。与其他森林相比,8年生尾巨桉人工林生物量高于广东雷州半岛地区25年生更新橡胶林生物量[24]、与海南30 年生的木莲人工林生物量持平[25],低于热带雨林及其他森林类型生物量[5,14-15,26-29],但尾巨桉人工林平均生产力达 17.19 t hm-2a-1,高于10 年生尾叶桉林 (15.98 t hm-2a-1)[11]、32 年生楠木人工林(8.57 t hm-2a-1)[29]等一般的森林类型的年平均生产力,接近海南尾细桉人工林年净生产力(17.56 t hm-2a-1)[30],表明巨尾桉具有很高的光合作用效益,固碳潜力大,是一个速生丰产的优良造林树种。
4.2 结论
我国南方桉树人工林发展迅速,但也存在林地力衰退、林分产量下降等问题,如何实现桉树人工林可持续发展已成为国内外关注的焦点。本研究发现尾巨桉人工林乔木层各器官、地上凋落物及总生物量具有良好的优化增长模型,其总生物量的增长模型为 Y=-1.693×103+3.337×103X-1.761X2;林分总生物量随林龄而增加,1,2,3,5年生和8年生尾巨桉人工林生物量分别为 12.49,47.75,64.51,105.77和137.51 t/hm2;层次分配方面,乔木层占绝对优势,占54.80%—91.56%,且随林龄的增加而增大,其次为凋落物,灌木层和草本层生物量较小,分别占1.02%—6.47%和0.28%—24.33%,均随林龄的增加呈递减趋势;乔木层以干所占比例最高,占51.07%—98.48%,且随林龄而增加,枝、叶、根分别占5.76%—11.80%,2.17%—21.01%和6.72%—14.87%,均随林龄而下降;灌木层以枝所占比例最高,为37.89%—56.79%,叶和根分别占16.35%—34.24%和19.52%—39.52%,随林龄的变化均不大;草本层分配1—5年生以地上所占比例较大,8年生地下所占比例高达63.87%;8年生尾巨桉人工林总生物量与30年生的木莲人工林持平,低于热带雨林,但其年均净生产量高达17.19 t/hm2,是一个光合效率高、固碳潜力大的速生丰产优良造林树种。
致谢:本研究样点的布置得到了广西壮族自治区林业勘测设计院李春干老师的帮助;样地调查得到了中国科学院亚热带农业生态研究所植物生态学科组及广西大学林学院林学专业、生态专业等老师和学生的大力支持,特此致谢。
[1] Ding Z L,Fu B J,Han X G,Ge Q S,Guo ZT,Wang Y,Fan J,Huang Y,Yu G R,Cai Z C,Shi X Z,Gu X F.Brief introduction to a cluster of Projects of“Research in Key Issues of International Negotiation with Regard to Coping with Climate Change”by CAS.Bulletin of the Chinese Academy of Sciences,2009,24(1):8-17.
[2] Watson R T.Intergovernmental Panel on Climate Change(IPCC)Special Report:Land Use, Land-use Change and Forestry.Cambridge:Cambridge University Press,2000.
[3] Woodwell G M,Whittaker R H,Reiners W A,Likens G E,Delwiche C C,Botkin D B.The biota and the world carbon budget.Science,1978,199(4325):141-146.
[4] Schimel D S.Terrestrial ecosystems and the carbon cycle.Global Change Biology,1995,1(1):77-91.
[5] Keller M,Palace M,Hurtt G.Biomass estimation in the Tapajos National Forest,Brazil:Examination of sampling and allometric uncertainties.Forest Ecology and Management,2001,154(3):371-382.
[6] Brown S,Gillespie A JR,Lugo A E.Biomassestimationmethods for tropical forestswith applications to forest inventory data.Forest Science,1989,35(4):881-902.
[7] Garkoti S C.Estimates of biomass and primary productivity in a high-altitude maple forest of the west central Himalayas.Ecological Research,2008,23(1):41-49.
[8] Brown S,Lugo A E.Biomass of tropical forests:A new estimate based on forest volumes. Science, 1984, 223(4642):1290-1293.
[9] Houghton R A.Aboveground forest biomass and the global carbon balance.Global Change Biology,2005,11(6):945-958.
[10] ZakrzewskiW T,Duchesne I.Stem biomassmodel for jack pine(Pinus banksiana Lamb.) in Ontario. Forest Ecology and Management,2012,279:112-120.
[11] Wen Y G,Liang HW,Zhan L J,Zhou M Y,He B,Wang L H,Wei SH,Zheng B,Liu D J,Tang Z S.Biomass production and productivity of Eucalyptus urophylla.Journal of Tropical and Subtropical Botany,2000,8(2):123-127.
[12] Wen Y G,Chen F,Liu SR,Liang HW,Yuan C A,Zhu H G.Relationship between species diversity and biomass of Eucalyptus plantation in Guangxi.Scientia Silvae Sinicae,2008,44(4):14-19.
[13] Wen Y G.Ecological and Economic Problems of Eucalyptus and Scientific Development.Beijing: China Forestry Publishing House,2008.
[14] LüX T,Tang JW,He Y C,Duan W G,Song JP,Xu H L,Zhu SZ.Biomass and its allocation in tropical seasonal rain forest in Xishuangbanna,Southwest China.Acta Phytoecologica Sinica,2007,31(1):11-22.
[15] Ren H,Peng S L,Xiang Y C.Biomass and net primary productivity in an Acacia mangium plantation in Heshan,Guangdong,China.Acta Phytoecologica Sinica,2000,24(1):18-21.
[16] Wang W F,Lei Y C,Wang X F,Zhao H C.A review of forest biomassmodels.Journal of Northwest Forestry University,2008,23(2):58-63.
[17] Foody G M,Boyd D S,Cutler M E J.Predictive relations of tropical forest biomass from Landsat TM data and their transferability between regions.Remote Sensing of Environment,2003,85(4):463-474.
[18] Ketterings Q M,Coe R,van Noordwijk M,Ambagau Y,Palm C A.Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests.Forest Ecology and Management,2001,146(1/3):199-209.
[19] Nelson B W,Mesquita R,Pereira J L G,de Souza S G A,Batista G T,Couto L B.Allometric regressions for improved estimate of secondary forestbiomass in the central Amazon.Forest Ecology and Management,1999,117(1/3):149-167.
[20] Chave J,Riéra B,Dubois M A.Estimation of biomass in a neotropical forest of French Guiana: spatial and temporal variability.Journal of Tropical Ecology,2001,17(1):79-96.
[21] Peichl M,Leava N A,Kiely G.Above-and belowground ecosystem biomass,carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland.Plant and Soil,2012,350(1/2):281-296.
[22] Preece N D,Crowley G.M,Lawes M J,van Oosterzee P.Comparing above-ground biomass among forest types in the Wet Tropics:Small stems and plantation types matter in carbon accounting. Forest Ecology and Management, 2012, 264:228-237.
[23] Li G F,Ren H.Biomass and net primary productivity of the forests in different climatic zones of China.Tropical Geography,2004,24(4):306-310.
[24] Zhou Z Z,Zheng H S,Yin G T,Yang Z J,Chen K T.Biomass equations for rubber tree in Southern China.Forest Research,1995,8(6):624-629.
[25] Zhou G Y,Zeng Q B,Lin M X,Chen B F,Li Y D,Wu ZM.Biomass and nutrient allocation in Manglietia hainanensis plantation ecosystem at Jianfengling.Forest Research,1997,10(5):453-457.
[26] Huang Q,Li Y D,Lai J Z,Peng G J.Study on biomass of tropicalmountain rain forest in Limushan,Hainan Island.Chinese Journal of Plant Ecology,1991,15(3):197-206.
[27] Dang C L,Wu Z L,Zhang Q.A study on biomass of the ravine tropical rain forest in Xishuangbanna.Acta Botanica Yunnanica,1997,19(S9):123-128.
[28] Feng Z L,Zheng Z,Zhang JH,Cao M,Sha L Q,Deng JW.Biomass and its allocation of a tropical wet seasonal rain forest in Xishuangbanna.Acta Phytoecologica Sinica,1998,22(6):481-488.
[29] Ma M D,Jiang H,Liu Y J.Biomass,carbon content,carbon storage and their vertical distribution of Phoebe bourmei artificial stand.Scientia Silvae Sinicae,2008,44(3):34-39.
[30] Shi Z J,Xu T P,Gao JX,Song A Y,Yu C T,Zhang N N,Hu Z S.Carbon storage and its distribution of Eucalyptus urophylla×E.tereticornis plantations in Hainan Island,Southern China.Scientia Silvae Sinicae,2011,47(10):21-28.
参考文献:
[1] 丁仲礼,傅伯杰,韩兴国,葛全胜,郭正堂,王毅,樊杰,黄耀,于贵瑞,蔡祖聪,史学正,顾行发.中国科学院“应对气候变化国际谈判的关键科学问题”项目群简介.中国科学院院刊,2009,24(1):8-17.
[11] 温远光,梁宏温,招礼军,周敏毅,何斌,王凌晖,韦善华,郑白,刘德杰,唐再生.尾叶桉人工林生物量和生产力的研究.热带亚热带植物学报,2000,8(2):123-127.
[12] 温远光,陈放,刘世荣,梁宏温,元昌安,朱宏光.广西桉树人工林物种多样性与生物量关系.林业科学,2008,44(4):14-19.
[13] 温远光.桉树生态、社会问题与科学发展.北京:中国林业出版社,2008.
[14] 吕晓涛,唐建维,何有才,段文贵,宋军平,许海龙,朱胜忠.西双版纳热带季节雨林的生物量及其分配特征.植物生态学报,2007,31(1):11-22.
[15] 任海,彭少麟,向言词.鹤山马占相思人工林的生物量和净初级生产力.植物生态学报,2000,24(1):18-21.
[16] 王维枫,雷渊才,王雪峰,赵浩彦.森林生物量模型综述.西北林学院学报,2008,23(2):58-63.
[23] 李高飞,任海.中国不同气候带各类型森林的生物量和净第一性生产力.热带地理,2004,24(4):306-310.
[24] 周再知,郑海水,尹光天,杨曾奖,陈康泰.橡胶树生物量估测的数学模型.林业科学研究,1995,8(6):624-629.
[25] 周光益,曾庆波,林明献,陈步峰,李意德,吴仲民.海南木莲人工林生物量及养分分配.林业科学研究,1997,10(5):453-457.
[26] 黄全,李意德,赖巨章,彭国金.黎母山热带山地雨林生物量研究.植物生态学与地植物学学报,1991,15(3):197-206.
[27] 党承林,吴兆录,张强.西双版纳沟谷热带雨林的生物量研究.云南植物研究,1997,19(S9):123-128.
[28] 冯志立,郑征,张建侯,曹敏,沙丽清,邓继武.西双版纳热带湿性季节雨林生物量及其分配规律研究.植物生态学报,1998,22(6):481-488.
[29] 马明东,江洪,刘跃建.楠木人工林生态系统生物量、碳含量、碳贮量及其分布.林业科学,2008,44(3):34-39.
[30] 时忠杰,徐大平,高吉喜,宋爱云,于春堂,张宁南,胡哲森.海南岛尾细桉人工林碳贮量及其分布.林业科学,2011,47(10):21-28.
