吉林不同生境防风夏季光合特性
2014-12-21韩忠明王云贺林红梅宋来今杨利民
韩忠明,王云贺,2,林红梅,韩 梅,宋来今,杨利民,*
(1.吉林农业大学中药材学院,长春 130118; 2.中国科学院东北地理与农业生态研究所,长春 130012;3.吉林省和龙市科学技术情报研究所,合龙 133500)
光合作用是植物生长和一切代谢活动的生理基础,影响着生长、发育、生殖等植物个体生理过程和生存竞争、环境适应和演化等植物物种的发展过程[1]。尤其是药用植物,不同的地理位置,由于气候环境条件的不同,必然引起植株生长发育特性及光合作用对环境因子适应性的改变[2]。因此,研究不同地理环境下植物的光合特性,揭示植物对环境生态适应性机制,对于阐明植物在不同气候环境条件下的生理反应具有重要的理论和实际意义[2]。
防 风 [Saposhnikovia divaricata (Miq.)Schischk.]为伞形科防风属多年生草本植物,以未抽薹的干燥根入药,是我国常用大宗中药材之一[3]。具有解表祛风、胜湿、止痉的功效,用于治疗头疼感冒、风湿痹痛、风疹瘙痒、破伤风等[4]。野生防风主要分布于东北、西北和华北地区,吉林省主要分布在西部和中部地区,近年来,在吉林、辽宁、黑龙江、内蒙古、河北、山西等地均有大面积栽培,栽培面积迅速扩大,栽培防风已成为我国防风药材的主要来源。目前,防风的研究主要集中在化学成分、药理、药效[4-8]等方面,有关防风对环境因子的生理生态适应性研究较少,仅有少量关于其光合特性[9]和对水分适应性研究报道[10-11],而关于不同生境光合特性的研究尚未见报道。本文以吉林省栽培防风为研究对象,研究不同生境防风净光合速率、蒸腾速率及水分利用效率等日变化特征及其与生态因子之间关系,探讨防风的光合生理规律,为进一步研究防风的光合生理提供依据,同时还可以为未来气候变化下防风对相应生境的适应能力进行预测提供依据。
1 材料与方法
1.1 试验地概况
长春试验区(43°47'50″N、125°24'52″E)位于中国东北松辽平原腹地,属温带大陆性半湿润季风气侯区。年均气温4.8℃,最高35.7℃,最低-39.8℃,年均降水量522—615 mm,降水主要集中在6—8月,这3个月的降水量占全年降水70%,无霜期为140—150 d,日照时数2688 h,最热月(7月)平均气温23℃。
长岭试验区(44°05'15″N,124°08'19″E)位于吉林省中西部,属中温带亚干旱季风气候区。年平均气温4.5℃,最高37℃,最低-36.1℃;该地区年平均降水量为470.6 mm,主要集中在夏季6—8月,降水量约占全年的70%;无霜期136—163 d,年平均日照时数2882 h。
白城试验区(45°52'19″N,122°47'40″E)位于吉林省最北端,属中温带半干旱季风气候区。年平均气温在5℃左右,最高38℃,最低达-38℃;7月份平均气温最高,在23℃左右。年均降水量407.9 mm,无霜期157 d,年均日照时数2919 h。
1.2 试验设计
试验用防风种子来源于吉林农业大学药用植物园,经吉林农业大学中药材学院杨利民教授鉴定确定。5月5—10日在吉林省防风的自然分布区长春、长岭和白城三地同时播种,条播行距30 cm,苗出齐后按株距10 cm定苗。小区长15 m,宽1.2 m,小区面积18 m2。
1.3 测定方法
8月中旬选择晴天,用LCpro+全自动便携式光合仪(ADC BioScientific Ltd.,UK)测定自然条件下1年生防风光合作用及环境因子的日变化。测定前选择生长部位相同、大小一致、且完全展开的叶片进行挂牌标记,用CI-203(CID,Inc,USA)手持式激光叶面积仪测定放入光合作用系统叶室部分的防风叶面积,将测得数据输入系统,自6:00—18:00每隔2 h测1次,每次测6片叶。测定指标包括:净光合速率(Pn,μmolm-2s-1)、气孔导度(Gs,molm-2s-1)、蒸腾速率(E,mmolm-2s-1)、胞间 CO2浓度 (Ci,μmol/mol)、光合有效辐射(PAR,μmolm-2s-1)、大气 CO2浓度(Ca,μmol/mol)、大气温度(Ta,℃)、叶片温度(Tl,℃)、空气相对湿度(RH,%)等;水分利用效率
气孔限制值 Ls
每个样地取长健康的防风叶片10—15片,叶片鲜重在采摘后立即用电子天平(精度为0.0001)称量并记录,之后分别装入牛皮纸袋中,带回实验室在75℃恒温干燥箱中烘至恒重,测定其干重,重复3次。叶片含水量(LWC)采用烘干称重法,计算公式为:

土壤含水量soil water content(SWC)采用烘干法,计算公式为:

1.4 数据处理
对所测的数据取平均值,用DPS数据分析软件对数据进行通径分析和偏相关分析。
2 结果与分析
2.1 不同生境环境因子变化
2.1.1 光合有效辐射和大气CO2浓度日变化
由图1可以看出,不同生境PAR均呈午间高峰型单峰曲线,即早晚PAR较低,12:00 PAR最高;Ca6:00较高,然后下降,长春、长岭Ca降幅较大,白城Ca全天变化平缓图1。Ca日均值长春、长岭和白城分别是 366.29、359.19 和 363.19 μmol/mol,差异不显著。
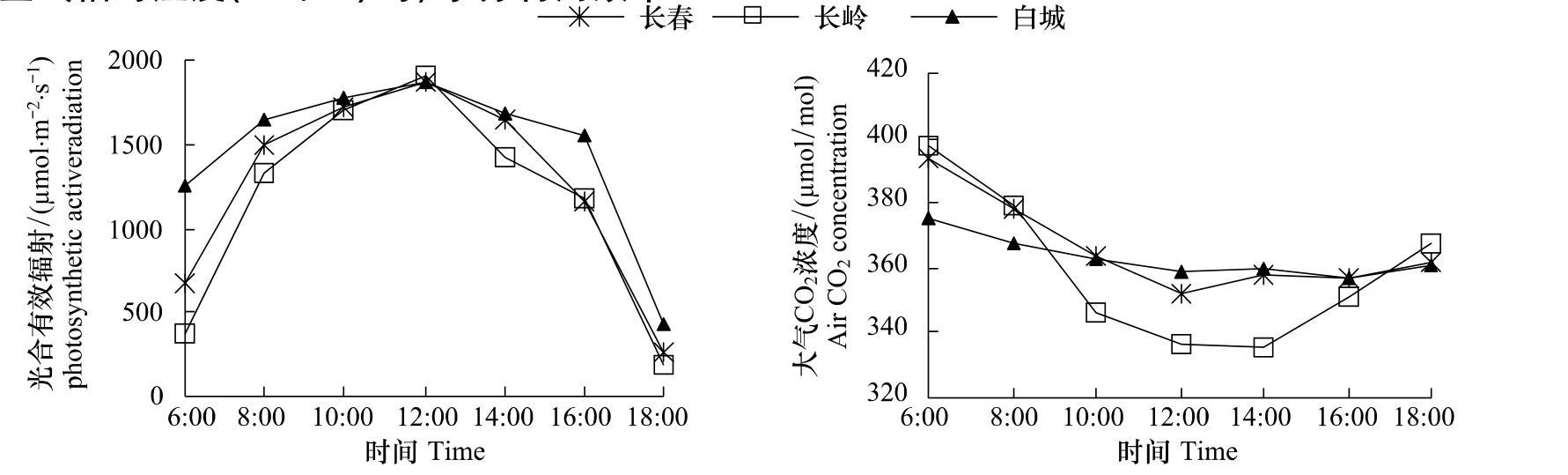
图1 光合有效辐射与大气CO2浓度日变化Fig.1 Diurnal changes of photosynthetic active radiation and air CO2 concentration
2.1.2 大气温度和防风叶片温度日变化
不同生境温度日变化见图2。中午温度高,呈单峰日均叶温分别是 31.35、32.99、22.61 ℃,差异不显著,与日均气温差值分别为0.70、0.67℃和0.27℃,白城地区叶温与气温差值明显低于长春和长岭地区。
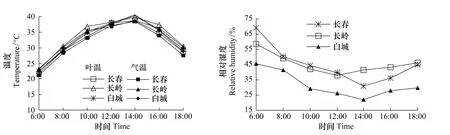
图2 不同生境温度与相对湿度日变化Fig.2 Diurnal changes of temperature and relative hum idity
2.1.3 空气相对湿度日变化
不同时期空气相对湿度日变化见图2,最大值在早晨,随后开始下降,长春和白城地区在14:00下降到最低点,长岭地区在12:00下降到最低点,白城地区的相对湿度明显低于长春和长岭。
2.2 不同生境防风光合特性的日变化
2.2.1 防风净光合速率日变化
图3表明,长春、长岭、白城3地防风净光合速率日变化呈明显双峰曲线变化,午间均出现光合“午休”现象;3个生境第1个高峰均出现在8:00,第2个高峰长春、长岭出现在14:00,白城出现在16:00。
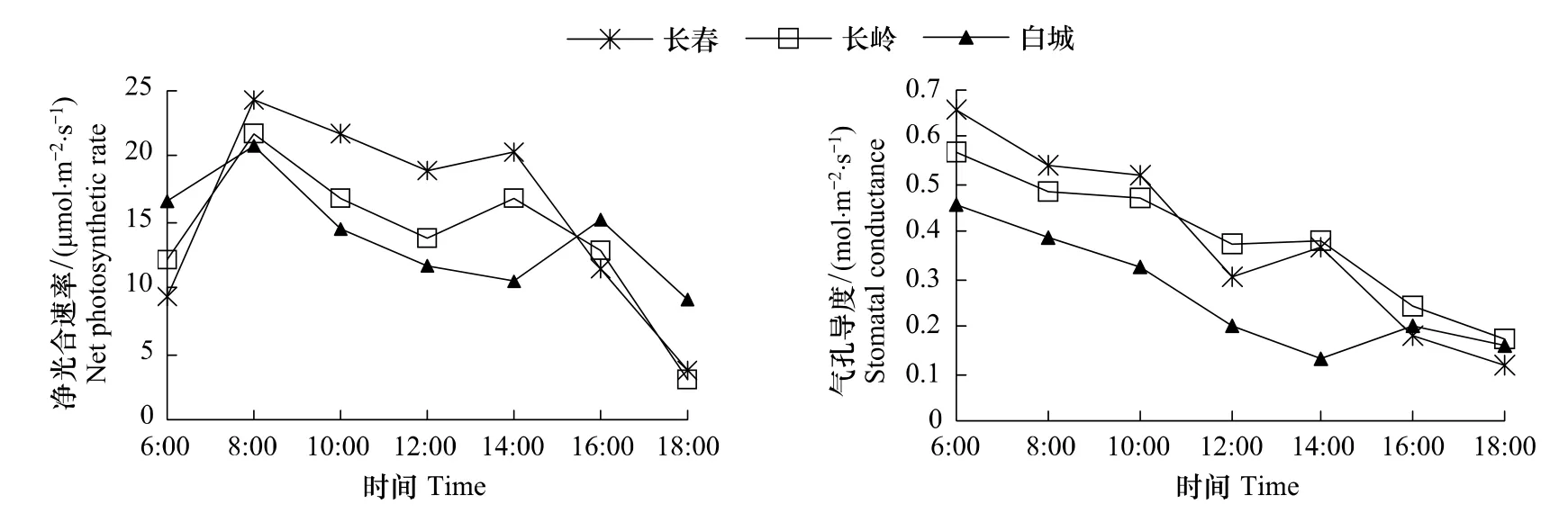
图3 防风净光合速率和气孔导度日变化Fig.3 Diurnal changes of net photosynthetic rate and stomatal conductance of S.divaricata
2.2.2 防风气孔导度日变化
3个生境防风Gs日变化趋势基本一致(图3),都是早晨较高,然后下降,长春、长岭在12:00出现低谷,白城在14:00出现低谷。长春、长岭、白城Gs日均值分别为 0.383、0.385、0.267 molm-2s-1,长春与长岭无显著差异,与白城差异极显著(P<0.01)。
2.2.3 防风胞间CO2浓度日变化
不同生境防风Ci日变化见图4,Ci都是早上高晚上低,中间波动平缓。白城胞间CO2浓度明显低于长春和长岭,长春、长岭和白城3地胞间CO2浓度日均值分别为260、273和234μmol/mol,长春与长岭差异不显著,长春、长岭与白城差异极显著(P<0.01)。
2.2.4 防风蒸腾速率日变化
图4显示防风蒸腾速率日变化均呈单峰曲线,长春最高峰出现在12:00,长岭和白城最高峰出现在14:00;白城防风蒸腾速率早晚高于长春和长岭,午间低于长春和长岭。这是由于白城地区土壤较干旱、午间气温较高,防风在高温、干旱胁迫下通过调节气孔避免损失过多的水分。
2.2.5 防风水分利用效率日变化
3个生境防风水分利用效率见图5,长岭、白城防风水分利用效率早晨6:00较高,之后开始下降;长春水分利用效率8:00最高,之后开始下降;午间3个生境防风水分利用效率降低最低点;随后水分利用效率回升,在16:00达到第2个高峰。
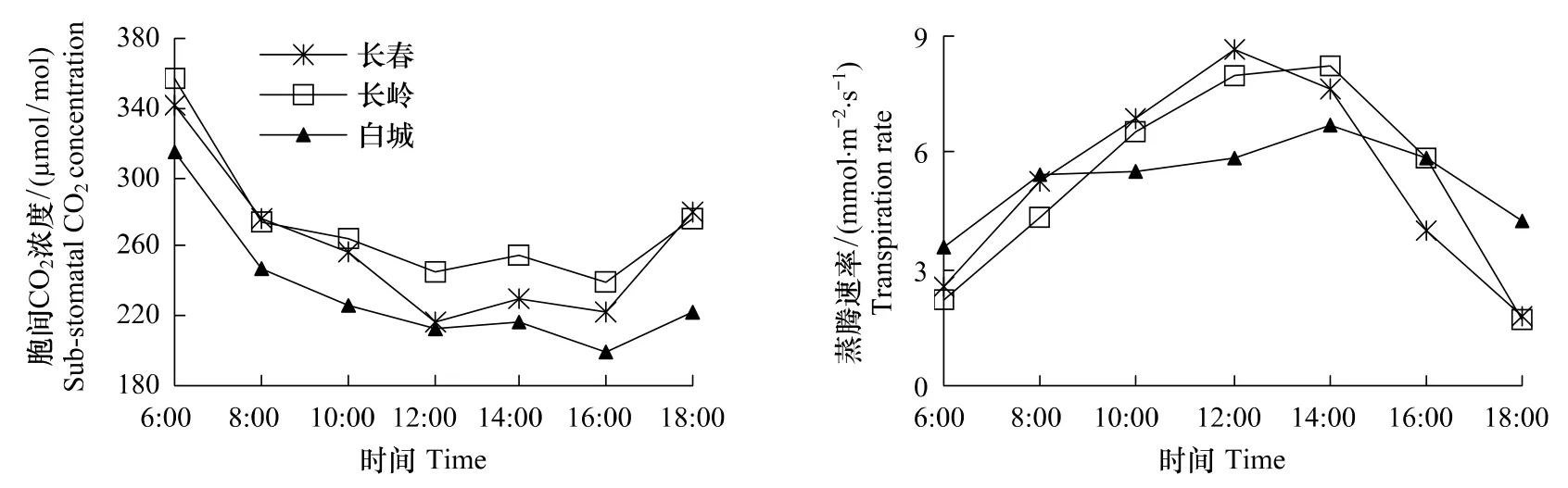
图4 防风胞间CO2浓度和蒸腾速率日变化Fig.4 Diurnal changes of sub-stomatal CO2 concentration and transpiration rate of S.divaricata
2.2.6 防风气孔限制值日变化
不同生境防风气孔限制值日变化具有一定的差异(图5)。长岭Ls在8:00—14:00明显低于长春和白城,以白城最高。长春、长岭、白城Ls日均值分别为0.292、0.242 和0.274,差异显著(P<0.05);长春与长岭差异极显著(P<0.01)。
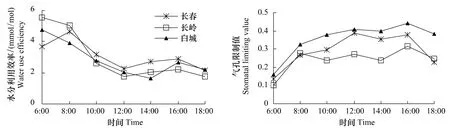
图5 防风水分利用效率和气孔限制值日变化Fig.5 Diurnal changes of water use efficiency and stomatal lim iting value of S.divaricata
2.3 土壤含水量和叶片含水量
不同生境土壤和叶片含水量见表1。不同生境土壤含水量间差异显著,8月份长春土壤含水量36.02%,是白城地区土壤含水量的1.77倍,是长岭地区的1.38倍;长春地区叶片含水量达68.72%,是长岭地区1.11倍,是白城地区1.21倍,3个地区间差异显著。

表1 不同生境土壤含水量和叶片含水量/%Table 1 Soil moisture content and leaf water content in different habitats
2.4 防风光合速率与环境因子的关系
3个生境防风Pn与环境因子通径分析和偏相关分析结果见表2和表3。长春环境因子中对Pn直接作用由大到小为 Tl>Ta>PAR>Ca>RH,其中 PAR 与Ta、Tl、Pn呈显著正相关(P<0.05),Ca与 Ta、Tl呈极显著负相关(P<0.01),与RH呈极显著正相关(P<0.01);Ta与 Tl呈极显著正相关 (P<0.01);Ta、Tl与RH均呈极显著负相关(P<0.01)。直接作用最大的 Tl,为负向,通过 PAR、Ta、RH 对 Pn的作用均为正向;环境因子中PAR与Pn相关系数最大,但受Ta的影响,对Pn的直接作用小于Tl。
长岭环境因子中对Pn直接作用由大到小为PAR>Tl>RH>Ca>Ta,其中 PAR 与 Ta、Tl呈显著正相关;Ca与 Ta、Tl呈极显著负相关,与RH呈极显著正相关;Ta与Tl呈极显著正相关,Ta、Tl与RH呈极显著负相关。环境因子中对Pn直接作用最大的PAR为正向,通过Ta与RH为正向,通过Ca、Tl为负向。
白城环境因子对Pn直接作用由大到小为Ta>Tl>RH>PAR>Ca。其中 Ca与 Ta、Tl呈显著负相关(P<0.05),Ca与 RH,Ta、Tl与 RH 呈极显著负相关(P<0.01)Ta与Tl呈极显著正相关(P<0.01);Ta对Pn直接作用最大,为正向,通过 PAR、Ca的作用为正 向,通过Tl和RH对Pn的作用为负向。
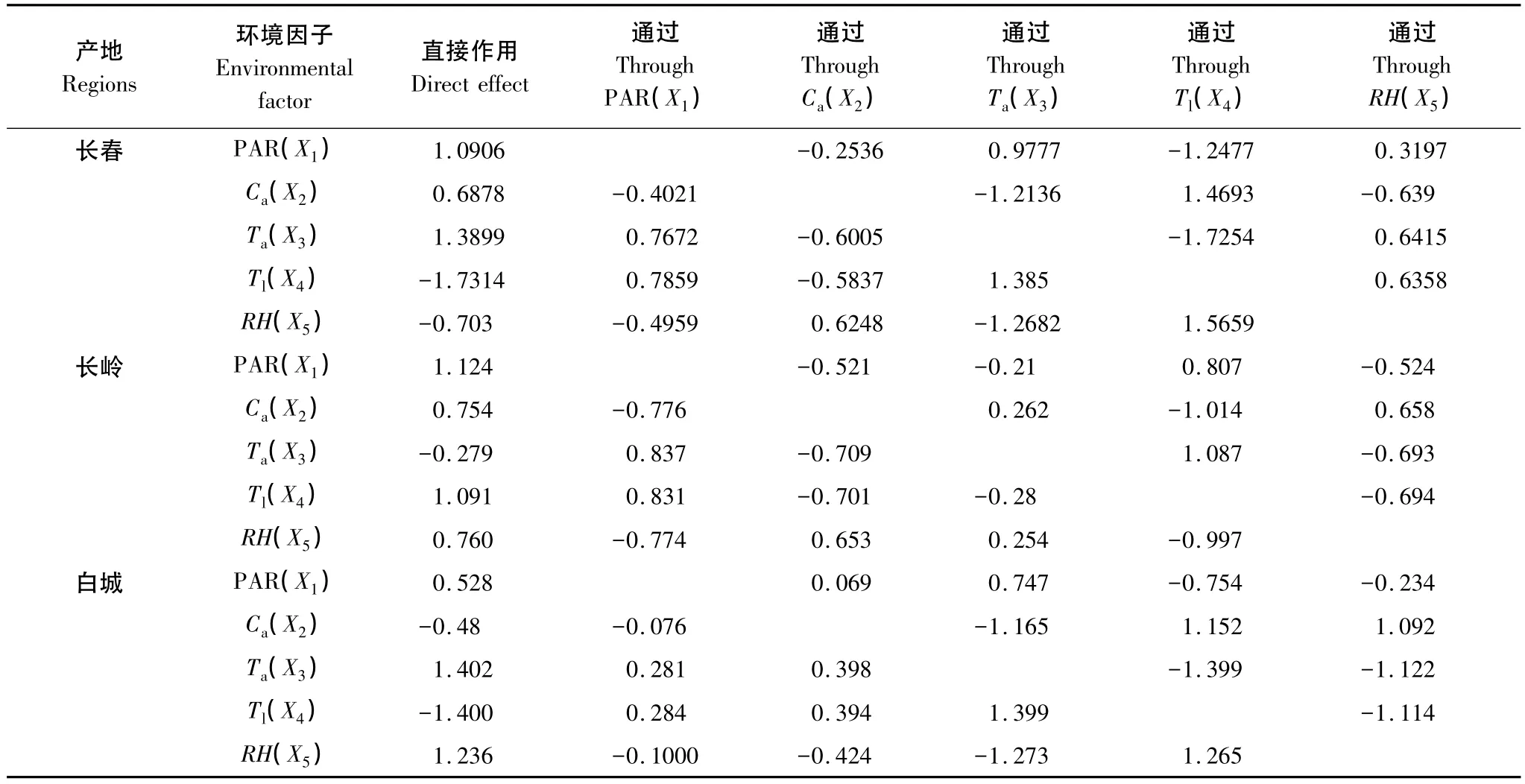
表2 环境因子对防风净光合速率的通径系数Table 2 Path coefficients between environmental factors and P n of S.divaricata

表3 不同生境环境因子与防风净光合速率偏相关系数(n=6)Table 3 Partial correlation coefficients between environmental factors and P n of S.divaricata
2.5 讨论与结论
植物光合作用是植物生产过程中物质积累与生理代谢的基本过程[12],通常植物光合作用日变化均有规律可循,变化曲线呈双峰型或单峰型[13]。本研究结果表明,3个生境防风净光合速率均呈双峰型曲线变化,具有明显的光合“午休”现象,参考Farquhar和Sharkeyy[14]提出的“当午间光合速率降低时,如果细胞间隙CO2浓度(Ci)随之降低,而气孔限制值(Ls)升高则属于光合作用的气孔限制,反之,而当午休出现时,Ci上升,Ls下降,此时光合速率的下降以非气孔限制”的判断原则:长春和长岭的防风Pn和Ci在午间是同步降低的,而Ls则呈同步增加的趋势,表明长春和长岭两地防风的Pn下降主要是由气孔导度(Gs)的下降造成的。白城的防风Pn在中午出现降低时,虽然Ci和Ls的升降不明显,但总体呈Ci升高,Ls降低的趋势,表明白城的防风Pn下降主要是由叶肉光合能力下降造成的,即非气孔限制因素引起的,这可能与白城地区土壤水分有关,白城地区夏季高温少雨、干旱,在干旱情况下防风的光合作用受到了极大的非气孔限制,光合能力下降,这是防风为了适应水分亏欠而形成的一种生理反应机制。
植物的光合生理过程与叶片组织细胞结构的变化有着密切的关系[15]。经统计分析,3个生境防风的Pn与Gs之间均呈显著的正相关关系。白城的防风Pn最低,主要是白城的Gs较低,比其他地区的低44%左右,气孔导度受环境因子的影响很大,适宜的光强有利于气孔开张,气孔阻力降低,气孔导度增大,在土壤水分充足时,植物为避免高温灼伤叶片,增大气孔开度,通过蒸腾作用带走叶片热量,但当土壤水分亏缺、气温较高时,植物则减小气孔的开度,降低蒸腾作用,这是植物适应生态环境实现自我保护的生理机制。长春地区土壤含水为36.02%,叶片含水为68.72%,空气中相对湿度也较高,当空气温度升高时,防风叶片可以通过蒸腾作用带走叶片热量,因此呈现较高的净光合速率;而白城地区土壤为沙土,土壤水分含量很低,只有20.35%,而空气温度较高,强辐射使气孔叶片蒸腾失水加剧,叶温升高,防风磷酸化、电子传递、酶活性等受到抑制,光合速率降低。
水分利用效率是评价植物对环境适应能力的综合指标[16],适宜的水分可以调节水分利用过程,从而提高水分利用率。防风叶片WUE取决于Pn与E的比值,光合速率和蒸腾速率的不同步是引起防风水分利用效率变化的主要原因。长岭、白城防风水分利用效率6:00较高,之后开始下降;长春水分利用效率8:00最高,之后开始下降;光合“午休”现象的出现导致午间3个生境防风水分利用效率降至最低点,随后水分利用效率回升,在16:00达到第2个高峰。有研究表明[17],适度干旱条件下植物的水分利用效率较高,但过度干旱反而导致水分利用效率降低,白城地区防风水分利用效率日均值最低,这主要与白城地区气候条件有关,白城地区属于中温带半干旱季风气候,降水偏少,从而导致土壤比较干旱,含水量只有20.35%,仅是长春土壤含水量的56.50%,土壤干旱,防风从土壤吸收的水分减少,从而导致防风叶片含水量也很低,仅仅为56.60%,加上午间高温,防风磷酸化、电子传递、酶活性等受到抑制,光合速率降低,从而导致水分利用效率降低。
生境条件的差异可以引起植物生理特征的可塑性反应,许多生理过程均与相关酶的活性有着密切的联系。而温度与酶活性紧密相关,Pn对Ta敏感性较高[18]。一般来说 Pn与 Ta呈抛物线形关系[19-20],当Ta小于临界值时,Ta增加会提高光合酶的活性[21],光合速率增大,一旦超过临界值Ta增加将会导致光合速率的降低[22]。长期高温胁迫可明显降低植物Pn、叶绿素含量和光化学效率等[23]。本研究中白城地区防风出现“午休”现象时,最高温度达40.2℃,造成Ta对Pn的抑制效应,直接通径系数最大,达1.402,呈负相关;而长春、长岭地区的防风Ta对Pn的直接通径系数较小,呈正相关。通径系数的大小能够在一定程度上反映出因果变量之间的作用强度,通径分析结果表明:不同生境防风叶片对不同环境因子的反映有所不同,环境因子中对Pn直接作用由大到小长春为 Tl>Ta>PAR>Ca>RH,长岭为 PAR>Tl>RH>Ca>Ta,白城为 Ta>Tl>RH>PAR>Ca。环境因子中,长春、长岭PAR与Pn相关系数最大,而白城地区Ca与Pn相关系数最大,在影响防风光合作用的4个环境因子中,PAR是驱动因子,它的变化驱动了Ta、Tl的变化,Ta升高引起空气密度的降低是午间RH和Ca降低的一个重要原因。
[1] Shi SL,Wang Y C,Zhou H B,Zhou JH.Comparative analysis of water related parameters and photosynthetic characteristics in the endangered plant Tetraena mongolica Maxim.and the closely related Zygophyllum xanthoxylon (Bunge) Maxim.Acta Ecologica Sinica,2012,32(4):159-169.
[2] Li H,Li D.Study on photosynthesis characteristics of Dipsacus asper in different habitats.Journal of Hubei University for Nationalities:Natural Science Edition,2011,29(2):227-231.
[3] Editorial Committee of the Pharmacopoeia of People's Republic of China.Pharmacopoeia of the People's Republic of China(PartⅠ).Beijing:China Chemical Industry Press,2010:140-140.
[4] Jiang Y Y,Liu B,Shi R B,Tu G Z.Isolation and structure identification of chemical constituents from Saposhnikovia divaricata(Turcz.)Schischk.Acta Pharmaceutica Sinica,2007,42(5):505-510.
[5] Han ZM,Wang Y H,Han M,Zhao S J,Yang LM.Separation and purification of prim-O-glucosylcimifugin and 5-O-methylvisammioside from Saposhnikovia Divaricata by high-speed counter-current chromatography.Chinese Journal of Analytical Chemistry,2009,37(11):1679-1682.
[6] Han ZM,Yang R Y,Wang Y H,Han M,KongW W,Yang L M.Extraction of chromone from Saposhnikovia divaricata by ultrasonic wave.Lishizhen Medicine and Materia Medica Research,2008,19(12):3035-3037.
[7] Li L,Zhang C,Liu Y Y,Geng L D,Xiao Y Q.Quality specification standards of Saposhnicovia divaricata (Turcz.)Schischk.Journal of Beijing University of Traditional Chinese Medicine,2005,28(3):58-61.
[8] Xiao Y Q,Li L,Yang B,Huang L Q.Studies on chemical constituents from root of Saposhnikovia divaricata (Turcz.)Schischk.China Journal of Chinese Materia Medica,2001,26(2):117-119.
[9] Han ZM,Wang Y H,Han M,Yang LM.Comparative study on photosynthesis of medicinal plant Saposhnikovia divaricata in different periods. Acta Ecologica Sinica, 2009, 29(3):1382-1390.
[10] Han ZM,Zhao S J,Wang Y H,Han M,Yang L M.Effect of water supply on photosynthetic characteristics and growth of Saposhnikovia Divaricata.Journal of Irrigation and Drainage,2010,29(3):98-101.
[11] Han ZM,Wang Y H,Zhao S J,Han M,Yang LM.Effects of water supply on growth and quality of Saposhnikovia divaricata.Journal of South China Agricultural University,2009,30(3):4-7.
[12] Kobayashi T,Okamoto K,Hori Y.Differences in field gas exchange and water relations between a C3dicot(Plantago asiatica)and C4monocot(Eleusine indica).Photosynthetica,1999,37(1):123-130.
[13] Zou Q.Studies on Physiological Drought Resistance of Crop.Ji'nan:Shandong Science and Technology Press,1994:155-163.
[14] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis.Annual Review of Plant Physiology,1982,33(1):317-345.
[15] Guo X Y,Yang Y F,Dong L J.Studies on the photosynthetic characteristics of the leaves of Phragmites communis in different dry land habitats in the Songnen Plains in China.Acta Prataculturae Sinica,2003,12(3):16-21.
[16] Ueda Y,Nishihara S,Tomita H,Oda Y.Photosynthetic response of Japanese rose species Rosa bracteata and Rosa rugosa to temperature and light.Scientia Horticulturae,2000,84(3/4):365-371.
[17] Stanhill G.Water use efficiency.Advances in Agronomy,1986,39:53-85.
[18] Xu Q Z,Huang B R.Effects of differential air and soil temperature on carbohydrate metabolism in creeping bentgrass.Crop Science,2000,40(5):1368-1374.
[19] Zhang X Q,Xu D Y.Effects of temperature on the photosynthetic physio-ecology of 18-year-old Chinese fir (Cunninghamia lanceolata(lamb.)hook.).Scientia Silvae Sinicae,2002,38(3):27-33.
[20] ZhangW,LiX Y,Li LG,Zhao L Z.The study of photosynthetic rate and transpiration rate on Hippophae rhamnoides L in the west of inner Mongolia.Acta Agriculturae Boreali-Sinica,2007,22(3):97-100.
[21] Haldimann P,Feller U.Inhibition of photosynthesis by high temperature in oak(Quercus pubescens L.)leaves grown under natural conditions closely correlates with a reversible heatdependent reduction of the activation state of ribulose-1,5-bisphosphate carboxylase /oxygenase. Plant Cell and Environment,2004,27(9):1169-1183.
[22] Salvucci M E,Crafts-Brandner S J.Inhibition of photosynthesis by heat stress:the activation state of Rubisco as a limiting factor in photosynthesis.Physiologia Plantarum,2004,120(2):179-186.
[23] Liu X Z,Huang B R.Photosynthetic acclimation to high temperatures associated with heat tolerance in creeping bentgrass.Journal of Plant Physiology,2008,165(18):1947-1953.
参考文献:
[1] 石松利,王迎春,周红兵,周健华.濒危种四合木与其近缘种霸王水分关系参数和光合特性的比较.生态学报,2012,32(4):159-169.
[2] 李慧,丁莉.不同生境条件下续断的光合特性研究.湖北民族学院学报:自然科学版,2011,29(2):227-231.
[3] 国家药典委员会.中华人民共和国药典(Ⅰ部).北京:化学工业出版社,2010:140-140.
[4] 姜艳艳,刘斌,石任兵,涂光忠.防风化学成分的分离与结构鉴定.药学学报,2007,42(5):505-510.
[5] 韩忠明,王云贺,韩梅,赵淑杰,杨利民.高速逆流色谱分离纯化防风中升麻素苷和5-O-甲基维斯阿米醇苷.分析化学,2009,37(11):1679-1682.
[6] 韩忠明,杨荣艳,王云贺,韩梅,孔伟伟,杨利民.超声波提取防风色原酮的研究.时珍国医国药,2008,19(12):3035-3037.
[7] 李丽,张村,刘元艳,耿立冬,肖永庆.防风药材质量标准研究.北京中医药大学学报,2005,28(3):58-61.
[8] 肖永庆,李丽,杨滨,黄璐琦.防风化学成分研究.中国中药杂志,2001,26(2):117-119.
[9] 韩忠明,王云贺,韩梅,杨利民.药用植物防风(Saposhnikovia divaricata)不同时期光合作用特性比较.生态学报,2009,29(3):1382-1390.
[10] 韩忠明,赵淑杰,王云贺,韩梅,杨利民.水分供给量对"防风"光合特性和生长发育的影响.灌溉排水学报,2010,29(3):98-101.
[11] 韩忠明,王云贺,赵淑杰,韩梅,杨利民.不同供水量对防风生长发育和品质的影响.华南农业大学学报,2009,30(3):4-7.
[13] 邹琦.作物抗旱生理生态研究.济南:山东科学技术出版社,1994:155-163.
[15] 郭晓云,杨允菲,李建东.松嫩平原不同旱地生境芦苇的光合特性研究.草业学报,2003,12(3):16-21.
[19] 张小全,徐德应.温度对杉木中龄林针叶光合生理生态的影响.林业科学,2002,38(3):27-33.
[20] 张玮,李晓燕,李连国,赵灵芝.内蒙古西部地区沙棘光合与蒸腾特性研究.华北农学报,2007,22(3):97-100.
