NADPH氧化酶及其检测方法的研究进展*
2014-12-05马松涛
邓 碧,马松涛
成都医学院 药学院(成都 610500)
NADPH氧化酶是一种与胞浆膜相关的多酶复合物,主要分布在胚层来源的细胞中,其以胞质NADPH为电子供体,催化胞外的O2生成超氧阴离子(O2-)。NADPH氧化酶最早在吞噬细胞中被发现,由 gp91phox、p22phox、p47phox、p67phox、p40phox和Rac 6种亚基组成。后来在非吞噬型细胞,如成纤维细胞(fibroblasts)、内皮细胞(endothelial cells)、血管平滑肌细胞(vascular smooth muscles,VSMC)、肾小球系膜细胞(mesangial cells)、肾小管细胞(renal tubular cells)中发现有类似吞噬细胞型的NADPH氧化酶。由于NADPH氧化酶生成的低量ROS可作为第二信使影响氧化还原敏感信号通路(redox-sensitive signal transduction pathways)。当 NADPH 氧化酶的活性上调时,大量生成的ROS可导致氧化应激的形成。动物模型和人体研究发现,在炎症、高血压、糖尿病、动脉硬化和阿尔茨海默病等疾病中,NADPH氧化酶介导的ROS过度产生是这些疾病的重要发病机理。NADPH氧化酶及作为第二信使的ROS在临床疾病的预防、诊断和治疗中有重要意义。目前对其常用的检测方法有分光光度法、化学发光法、荧光光度法和电子自旋共振法,但其原理和检测结果均有一定差异。因此,本文就NADPH氧化酶的相关研究进展及活性检测方法综述如下。
1 NADPH氧化酶
1.1 NADPH氧化酶结构
NADPH氧化酶主要由p40phox、p47phox、p67phox、小GTPase和Rac形成复合体RhoGDI存在于细胞质中,p22phox和gp91phox以异二聚体cyt b558存在于细胞膜中,其中gp91phox存在一系列同 系 物,分 别为 NOX1、NOX2(gp91phox)、NOX3、NOX4、NOX5、DUOX1、DUOX2,统称为NOX蛋白家族。按照结构的不同(图1),可将其分为3组:NOX1~4:结构中包含部分伸向胞浆的C末端和疏水跨膜区 N端,C端包含 NADPH-和FAD-结合位点,N端有6个保守的跨膜区,其中在第3个和第5个跨膜区各有2个结合亚铁血红素的保守氨基酸残基;NOX5:其结构除了NOX1~4的结构域外,在N末端有4个EF手结构域,为Ca2+结合位点;DUOX氧化酶:包含NOX1~5结构(与NOX5不同的是其含有2个EF手结构),在N末端有类超氧化物酶样区域。
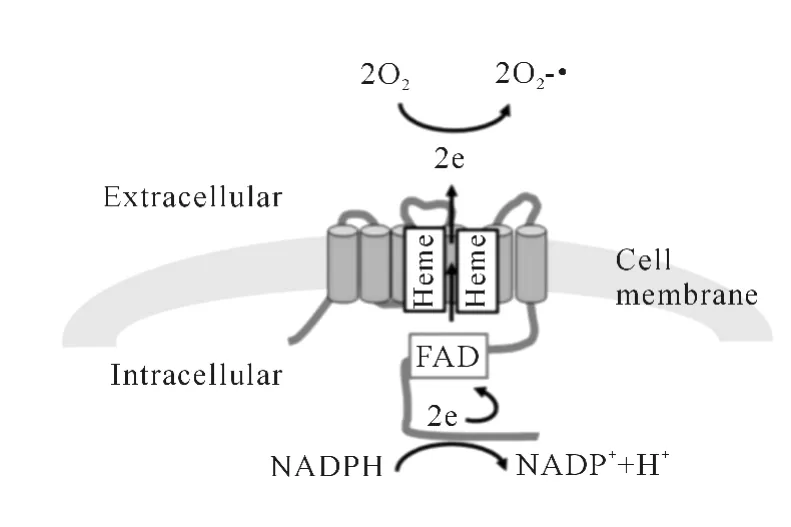
图1 NOX催化反应示意图
1.2 NADPH氧化酶的激活与ROS的产生
静息状态下,NADPH氧化酶无活性。在受到细胞外信息(图1),如激素、细胞因子甚至细菌等的刺激时,NADPH氧化酶通过其胞质成分p47phox的磷酸化后,分子间紧密结合打开,自动抑制区释放,同时SH3区和PX区暴露,p67phox利用C端的SH3区结合p47phox的PRR区,即实现了磷酸化的p47phox携带p67phox、p40phox和Rac靠近胞膜,通过与p22phox亚基上富含脯氨酸的尾巴结合形成复合体,使gp91phox的构象发生改变,即形成有活性的NADPH氧化酶[1]。复合体的激活还需2种低分子量的鸟嘌呤核苷酸结合蛋白:Rac2和Rap1A[2],其中,Rac2位于胞浆中,与 Rho-GDI形成二聚体,激活时,Rac2结合GTP后与p67phox结合。发生吞噬时,质膜内化,成为吞噬小泡的内壁[3]。O2-通过酶复合体释放到囊泡中,反应为2O2+NADPH→2O2-·+NADP++H+,以 O2-为原料,产生一系列的活性氧(ROS),包括超氧化物、羟自由基、过氧化氢和一些能轻易转化为自由基的物质(次氯酸、臭氧及单态氧),如:2O2-·+2H+→O2+H2O2;Rap1A是一种胞膜蛋白,其和cyt b558通过囊泡融合和分泌形式转移到质膜,从而促进蛋白释放到细胞外。一般把O2-·当成NOX的最初产物,但在DUOX和NOX4催化的反应中,可在没O2-·情况下检测到H2O2,由于对NOX产物的不确定性,一般把O2-·和H2O2当成NOX的产物,即ROS。
1.3 NADPH氧化酶及ROS在疾病中的作用
NADPH酶通过不同的氧化还原信号介导不同生物组织中的功能。慢性肉芽肿(chronic granulomatous disease,CGD)主要由NOX2的缺陷引起[4]。血管NOX产生的ROS对维持正常的心血管健康起重要作用,ROS通过减少内皮细胞来源的松弛因子,降低NO的生物利用度,进而调节血压[5-6]。健康肾脏ROS主要由 NOX3产生,其通过控制Na+转运,球管反馈及肾脏氧化等途径调控肾脏功能[7-8]。肺脏中 NOX2参与气管和血管的重塑[9]。结肠黏膜中DUOX和NOX1介导产生的ROS能促进血清素的合成,对调节结肠分泌和运动功能十分重要。小胶质细胞表达NOX2和p22phox,这两种酶在神经因子匮乏时参与小角质细胞增殖和凋亡的调控[10]。NADPH 酶产生的ROS通过参与调节TLR4介导VSMC增殖和炎症表型可作为第二信使,激活细胞内的信号转导通路,减少有活性的NO导致的内皮细胞功能障碍,从而在动脉粥样硬化的内膜增生和炎症反应过程中起重要作用[11]。NOX家族的过表达是许多肿瘤细胞的特征之一,其中ROS在肿瘤形成的各阶段发挥重要作用。研究[12]表明,NOX4是治疗肿瘤转移的一个潜在靶点。通过PKC介导NOX4的过度表达参与肾小球硬化和细胞外基质的重建,最终诱导糖尿病肾病的发生[13]。在阿尔茨海默病中,研究[14-16]发现NADPH氧化酶过度激活是其重要的发病原因。
2 NADPH氧化酶活性检测方法
NADPH氧化酶的活性检测中,大部分研究通过运用能与ROS反应生成具有特殊性能产物的探针来对其进行表征。ROS的测定方法主要有分光光度法、化学发光法、荧光光度法和电子自旋共振法(表1)。
2.1 电子自旋共振光谱检测
电子自旋共振(electron spin resonance,ESR)光谱法主要通过未配对的电子对其进行检测,由于O2-·的半衰期较短,在细胞中的浓度很低及检测方法灵敏度不够,故采用电子自旋捕捉剂如苯基叔丁基氮氧化合物 PBN、1-氧基-4-吡啶基-N-叔丁基氮氧化合物POBN等对不稳定的自由基进行捕捉。刘玲等[17]通过采用ESR光谱技术研究食品加工储藏中与氧化相关的自由基变化,其借助捕集剂能够检测出美拉德反应时及时出现的小分子不稳定自由基。虽然捕集剂能增强ESR信号,但其仍存在一些不利影响:由于细胞环境有一定还原性,很多以硝基酮作为捕集剂的分子很快会转化为无ESR信号的分子,即导致出现假阴性结果[18]。体内细胞和培养细胞中的ESR信号会通过酶代谢或Vc等物质还原,故细胞环境ESR信号实际只代表细胞外信号[19]。但在严格设有对照组的情况下,ESR光谱可以用来检测O2-·和NOX活性。
2.2 分光光度法
细胞色素C还原法和NBT法是分光光度法常用的方法,该法是检测NADPH氧化酶活性使用最广泛的方法。由于细胞色素C具有比O2-·高的电子势能,O2-·将其未配对的电子转移给高铁细胞色素C(Fe3+)将其转变为亚铁细胞色素 C(Fe2+),导致其在550nm处的吸光度增加。由于存在很多分子和酶能还原高铁细胞色素C,吸光度的改变不能说明是O2-·引起的。细胞色素C不能透过细胞膜,对于检测较低浓度的O2-·,该法不是很灵敏,主要用来检测细胞裂解物中NOX活性和吞噬细胞中的NOX2。NBT可渗透细胞,进入细胞后,被O2-·通过两步反应还原,其中包括一步电子反应和一步自由基中间体反应。NBT首先通过O2-·还原为NBT+·,NBT+·被歧化或者通过再接收O2-·的1个电子形成稳定的NBTH+,NBTH+是一种蓝色结晶,可通过550nm的吸光度对其进行检测,大部分实验用DMSO溶解该结晶后,通过检测630nm处的吸光度对其进行检测。NBT法主要用于对CGD病人的确诊。

表1 用于检测ROS的探针
2.3 化学发光法
化学发光法是检测自由基较为灵敏的方法。目前为止,已经有很多化学发光探针用来检测NOX。由于在一定条件下,ROS能与鲁米诺(Luminol)、光泽精(Lucigenin)及其类似物等发光试剂发生反应,产生不同强度和发射波长的化学光辐射。目前鲁米诺化学发光法用于精子与精液、血液与组织匀浆上清液、细胞等[20]样品中 ROS检测,在李颖畅等[21]报道中用于检测蓝莓花色苷对氧自由基的清除作用。但·OH、O2-·、H2O2均能氧化鲁米诺产生化学发光,故不用来检测单一的ROS。光泽精主要用于细胞外自由基的测定。这2种探针主要的区别是:1)光泽精主要被O2-·还原为光泽精阳离子,而鲁米诺主要被氧化发光;2)光泽精对O2-·有选择性而鲁米诺检测ROS类型不具有明确性;3)光泽精不能渗透细胞而鲁米诺可渗透细胞,故可用来检测细胞内和细胞外自由基。
除此以外,使用较多的为L-012,其为光泽精类似物,对于检测NOX的活性和灵敏度是光泽精的100倍左右,其用于检测体内依靠NOX2产生的O2-·[22]。另一种使用较多的是 MCLA,其对于O2-·具有敏感性和特异性,无自氧化作用,稳定性好,故其广泛用于检测肝脏、心脏、肺脏和血管组织的ROS,但MCLA的使用需注意:过氧亚硝基阴离子(ONOO-)能诱导其发光,还能发生自氧化导致较强的荧光背景,因此在实验中可考虑加入活性氮抑制剂,增加结果的准确性。
2.4 荧光光度法
荧光光度法用合适的探针作用于细胞,与ROS反应后,探针的化学结构发生变化,生成具有强荧光的产物,通过检测反应产物的强度可反映ROS的水平。目前使用较多的是二氯荧光素(dichlorodihydrofluorescein,DCFHDA)和氢化乙啡啶(hydroethidine,HE)。DCFHDA主要用于检测ROS和H2O2,DCFHDA能被细胞吸收并水解为DCFH,DCFH不发荧光,细胞内的DCFH被H2O2氧化为具有较强荧光的DCF,其激发波长为498nm,发射波长为525nm。Wang等[23]在研究中用此方法检测高糖处理的大鼠肾小球和肾脏系膜细胞的NADPH氧化酶活性的变化。通过对其标准操作进行稍微修改,DCFHDA可用于低定量检测nanoparticles(NPS)诱导的细胞氧化应激,通过将细胞与2mM DCFHDA孵育20min,用流式细胞仪检测细胞内的DCF含量,用来间接表示NADPH氧化酶活性[24]。对于DCFHDA的使用需注意:其除了能被 H2O2氧化外,ONOO-和 HOCl[22,25]也能氧化DCFH为DCF;DCF的反应会产生中间自由基,其很快与O2反应产生O2-·并生成H2O2,这种氧化还原循环会导致DCF荧光信号的放大。由于DCF及其荧光分子不具有选择性,其还能被·OH、ROOH、NO·等多种ROS氧化,实验所得结果不能简单当成H2O2或者NOX活性,还需相关的控制条件或者使用特定的NOX抑制剂进行辅助判断。
HE是一种亲脂物质,可穿过细胞膜,在细胞内,HE氧化为带正电的E+插入DNA,发出红色荧光,故把红色荧光产物(E+)作为细胞内O2-·的标志物。Heim等[26]用流式细胞仪通过检测细胞内的荧光强度来表示细胞内NADPH氧化酶的活性,马晓东等[27]发现使用激光共聚焦显微镜结合HE荧光标记技术可原位实现观察活细胞内NADPH氧化酶活性的变化。kalyanaraman 等[28]证实了2-OH-E+而不是E+是O2-·与 HE的反应产物,形成速率为(2.6±0.6)×105m-1s-1,激发波长为520nm,发射波长为610nm,用于定量测定细胞内O2-·,但其准确度相对较低。
目前虽然有很多的方法用于检测NOX活性及来源于NOX的O2-·和H2O2,但所有方法都存在一定的局限性。因此探针的使用应考虑实验的要求,故在选择探针时,可从以下几方面考虑:1)所使用探针的化学结构及性质是否需要加入辣根过氧化物等催化剂,探针本身是否会发生自氧化;2)方法的特异性,即所选用的探针是否对H2O2或O2-·具有选择性,如果没有,是否可能会存在干扰;3)组织或细胞是否会吸收探针;4)所测的实验材料(血清、细胞、组织)中的ROS含量是否与所测信号成线性关系;5)明确所分析的细胞或组织中表达哪种NOX或是NOX的哪种亚型;6)可同时使用几种探针来共同测定NOX来源的O2-·或H2O2;7)NOX是否参与ROS的产生,可考虑加入NOX特异性抑制剂如DPI,如果仍有ROS产生,可判断ROS不是来源于NOX。
2.5 检测NADPH氧化酶活性的其他方法
除了通过检测ROS的活性来间接表示NADPH氧化酶活性以外,组织和细胞中可通过下列方法来进行定量检测[29]:可通过用Trizol试剂提出 mRNA 后测定 p22phox、gp91phox、p40phox、p47phox、p67phox的mRNA表达情况,若其中某一亚基mRNA表达增多,则可判断NADPH氧化酶活性升高;用裂解液提取细胞和组织中的蛋白后,通过western blot方法检测蛋白表达情况,通过对比蛋白条带亮度,对相关亚基的表达情况进行定量分析。血清中各亚基的表达情况可通过用酶联免疫吸附试验进行比较[30]。除此以外,对于动物体内的氧化应激程度可通过检测24h尿液中的8-羟基脱氧鸟苷(8-hydroxy-2-deoxyguanosine,8-OHdG)和丙二醛(malondialdehyde,MDA)的含量来进行表示[31]。
3 结论与展望
NADPH氧化酶及NOX蛋白家族近年来受到广泛关注,其结构功能方面的研究已取得一定的进展,基因敲除技术及单克隆抗体的使用,有望在NOX家族的研究中发挥重要作用。NADPH氧化酶的过表达,介导很多疾病的发生发展,目前相关的抑制剂都缺乏特异性,能特异抑制NADPH氧化酶亚基的药物应该有很好的应用前景。对于NADPH氧化酶活性的检测,本文提供了一些目前较常用的方法,分析比较了各方法的选择性、灵敏度、适用性及优缺点,以便研究者开展相关实验时,能做出较好的选择,但在实验过程中应严格设置对照实验或使用NADPH氧化酶抑制剂做对照。除此以外,大部分研究除对ROS进行检测外,还通过RT-PCR检测NADPH氧化酶各亚基mRNA的表达情况,以及通过Western-Blot或酶联免疫检测氧化酶亚基的表达情况,从而使实验结果更具说服力。
[1]张洁洁,彭军.NADPH氧化酶激活机制和病理意义[J].中国药理学与毒理学杂志,2014,28(1):139-142.
[2]Diebold BA,Bokoch GM.Molecular basis for Rac2regulation of phagocyte NADPH oxidase[J].Nat Immunol,2001,2(3):211-215.
[3]Takahashi M,Dillon TJ,Liu C,etal.Protein kinase A-dependent phosphorylation of Rap1regulates its membrane localization and cell migration[J].J Biol Chem,2013,288(39):27712-27723.
[4]Desjardins A,Coignard-Biehler H,Mahlaoui N,etal.Chronic granulomatous disease:pathogenesis and therapy of associated fungal infections[J].Med Sci:Paris,2012,28(11):963-969.
[5]Cifuentes ME,Pagano PJ.Targeting reactive oxygen species in hypertension[J].Curr Opin Nephrol Hypertens,2006,15(2):179-186.
[6]Moncada S,Higgs EA.The discovery of nitric oxide and its role in vascular biology[J].Br J Pharmacol,2006,147(1):193-201.
[7]Wilcox CS.Oxidative stress and nitric oxide deficiency in the kidney:a critical link to hypertension?[J].Am J Physiol Regul Integr Comp Physiol,2005,289(4):913-935.
[8]Kambayashi Y,Ogino K.Reestimation of Cypridina luciferin analogs(MCLA)as a chemiluminescence probe to detect active oxygen species--cautionary note for use of MCLA[J].J Toxicol Sc,2003,28(3):139-148.
[9]Hoidal JR,Brar SS,Sturrock AB,etal.The role of endogenous NADPH oxidases in airway and pulmonary vascular smooth muscle function[J].Antioxid Redox Signal,2003,5(6):751-758.
[10]Mander PK,Jekabsone A,Brown GC.Microglia proliferation is regulated by hydrogen peroxide from NADPH oxidase[J].J Immunol,2006,176(2):1046-1052.
[11]皮燕.NADPH氧化酶调节TLR4介导的血管平滑肌细胞增殖和炎症表型的实验研究[D].重庆:第三军医大学,2013.
[12]张彪.NADPH氧化酶4是一个药物干预肿瘤转移的潜在靶点[D].杭州:浙江大学,2013.
[13]Thallas-Bonke V,Jha JC,Gray SP,etal.Nox-4deletion reduces oxidative stress and injury by PKC-alpha-associated mechanisms in diabetic nephropathy[J].Physiol Rep,2014,2(11):1-13.
[14]Choi JS,Islam MN,AliMY,etal.Effect of C-glycosylation on anti-diabetic,anti-Alzheimer,s disease and anti-inflammatory potential of apigenin[J].Food and Chemical Toxicology,2014,64:27-33
[15]Walsh KP,Minamide LS,Kane SJ,etal.Amyloid-beta and proinflammatory cytokines utilize a prion protein-dependent pathway to activate NADPH oxidase and induce cofilin-actin rods in hippocampal neurons[J].PLoS One,2014,9(4):95995.
[16]Ghosh D,Levault KR,Brewer GJ.Relative importance of redox buffers GSH and NAD(P)H in age-related neurodegeneration and Alzheimer disease-like mouse neurons[J].Aging Cell,2014,13(4):631-640.
[17]刘玲,纪淑娟,檀德宏,等.利用电子自旋共振技术研究美拉德反应自由基变化[J].农业工程学报,2013,29(6):280-286.
[18]Dikalov S,Griendling KK,Harrison DG.Measurement of reactive oxygen species in cardiovascular studies [J].Hypertension,2007,49(4):717-727.
[19]Witting PK,Rayner BS,Wu BJ,etal.Hydrogen peroxide promotes endothelial dysfunction by stimulating multiple sources of superoxide anion radical production and decreasing nitric oxide bioavailability[J].Cell Physiol Biochem,2007,20(5):255-268.
[20]Wardman P.Fluorescent and luminescent probes for measurement of oxidative and nitrosative species in cells and tissues:progress,pitfalls,and prospects[J].Free Radic Biol Med,2007,43(7):995-1022.
[21]李颖畅,孟宪军,修英涛,等.化学发光法测定蓝莓花色苷对氧自由基的清除作用[J].食品研究与开发,2008,29(3):72-75.
[22]Daiber A,August M,Baldus S,etal.Measurement of NAD(P)H oxidase-derived superoxide with the luminol analogue L-012[J].Free Radic Biol Med,2004,36(1):101-111.
[23]Wang Z,Ni J,Shao D,etal.Elevated transcriptional coactivator p102mediates angiotensin II type 1receptor upregulation and extracellular matrix overproduction in the high glucose-treated rat glomerular mesangial cells and isolated glomeruli[J].Eur J Pharmacol,2013,702(13):208-217.
[24]Aranda A,Sequedo L,Tolosa L,etal.Dichloro-dihydrofluorescein diacetate(DCFH-DA)assay:aquantitative method for oxidative stress assessment of nanoparticle-treated cells[J].Toxicol In Vitro,2013,27(2):954-963.
[25]Whiteman M,Rose P,Siau JL,etal.Hypochlorous acidmediated mitochondrial dysfunction and apoptosis in human hepatoma HepG2and human fetal liver cells:role of mitochondrial permeability transition[J].Free Radic Biol Med,2005,38(12):1571-1584.
[26]Heim RC,Park TS,Vogler GP,etal.Changes in hip migration afier selective dorsal rhizotomy for spastic quadriplegia in cerebral palsy[J].J Neurosurg,1995(82):567-571.
[27]马晓东,黄行许,朱晓亮,等.一种检测巨噬细胞内NADPH氧化酶活性的新方法[J].第一军医大学学报,2000,20(2):161-165.
[28]Kalyanaraman B,Zhao H,Joseph J,etal.Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence[J].Proc Natl Acad Sci USA,2005,102(16):5727-5732.
[29]Hu C,Cong XD,Dai DZ,etal.Argirein alleviates diabetic nephropathy through attenuating NADPH oxidase,Cx43,and PERK in renal tissue[J].Naunyn Schmiedebergs Arch Pharmacol,2011,383(3):309-319.
[30]真晓雯.亚临床甲状腺功能减低患者氧化应激指标gp91phox与LDl-C和TSH的相关性分析[D].济南:山东大学,2013.
[31]Nam SM,Lee MY,Koh JH,etal.Effects of NADPH oxidase inhibitor on diabetic nephropathy in OLETF rats:the role of reducing oxidative stress in its protective property[J].Diabetes Res Clin Pract,2009,83(2):176-182.
