大容量人源Fab噬菌体展示抗体库的构建及抗CD276抗体筛选
2014-10-31刘婕陈俊亨徐天殊王丽萍赵琦
刘婕,陈俊亨,徐天殊,王丽萍,赵琦
大容量人源Fab噬菌体展示抗体库的构建及抗CD276抗体筛选
刘婕,陈俊亨,徐天殊,王丽萍,赵琦
518055 深圳,中国科学院深圳先进技术研究院抗体药物研究中心(刘婕、陈俊亨、徐天殊、赵琦);130012 长春,吉林大学生命科学学院(陈俊亨、徐天殊、王丽萍)
构建大容量人源 Fab 噬菌体库,筛选并鉴定抗 CD276 的特异性抗体。
通过采集 10 位健康成人的外周血淋巴细胞,提取总 RNA,利用 RT-PCR 扩增人 Fab 抗体基因片段,插入噬菌体载体 pCANTAB5H 中,构建人源 Fab 噬菌体抗体库。通过固相化的抗原对抗体库进行 3 轮筛选后,随机挑取96 个单克隆进行 phage-ELISA 鉴定,筛选出与抗原具有较强结合性的噬菌体克隆。
成功构建库容为 5 × 1010的噬菌体抗体库,从中筛选到 11 株阳性克隆,对其进行测序鉴定,表明该 11 株克隆的序列均一致,ELISA 证实其对抗原 CD276 具有较强的结合性。
构建了大容量人源 Fab 抗体库,从中获得具有抗 CD276 的人源 Fab 抗体片段,为进一步的研究和应用提供实验基础。
免疫球蛋白 Fab 片段; 噬菌体展示抗体库; CD276
CD276,又叫 B7-H3,是 2001 年在人树突状细胞 cDNA 文库中鉴定出的B7 超家族共信号分子[1]。近年来的研究表明共信号分子不仅能提供T 细胞活化所需的共信号分子促进 T 细胞免疫应答[2],在抗炎和抗肿瘤免疫应答中发挥关键作用,同时也可以提供下调免疫应答的负性共信号分子,维持 T 细胞免疫耐受,在肿瘤免疫逃逸中发挥重要作用[3-5]。而 CD276 在正常组织、细胞中不表达或极低表达,高表达于多种肿瘤组织中[6]。本文以 CD276 为靶标,利用噬菌体抗体库技术克隆出全套人源抗体基因,并在噬菌体表面表达,构建大容量人源 Fab 抗体库,并从中筛选出特异性抗 CD276 的 Fab 抗体片段,为肿瘤药物的研制提供新的候选因子。
1 材料与方法
1.1 材料
噬菌体载体 pCANTAB5H 通过对商用载体 pCANTAB5E 进行酶切位点改造获得[7];大肠杆菌 TG1 购自美国Lucigen 公司;HB2151 细菌由香港大学张美云教授惠赠;Taq DNA 聚合酶、限制性内切酶和 T4 DNA 连接酶购自中国NEB 公司;Trizol、DNA 回收试剂盒、质粒小量抽提试剂盒购自德国 Qiagen 公司;RT-PCR 试剂盒购自美国Promega 公司;HRP 标记的鼠抗 Flag 抗体购自美国 Sigma 公司;HRP 标记的鼠抗 M13 抗体和淋巴细胞分离液购自美国 GE Healthcare 公司;CD276 蛋白购自美国R & D 公司;人外周血淋巴细胞来自 10 位健康人捐献的 2 L 外周血,采用 Ficoll 密度梯度离心分离;所有引物由美国Life Technologies 中国公司合成。
1.2 方法
1.2.1 人外周血淋巴细胞总 RNA 的抽提及 RT-PCR 从采集的 10 个健康人的 2 L 血液分离出淋巴细胞,用 Trizol 试剂提取淋巴细胞总 RNA,按总 RNA 提取试剂盒说明书所述步骤操作。以 Oligo(dT) 为引物反转录合成第一链 cDNA,具体方法按说明书进行。
1.2.2 重链和轻链基因的 PCR 扩增 引物设计参考文献[8]提供的轻链及重链家族特异性引物序列组合进行 PCR 扩增抗体 VH、λ 和κ片段编码区。然后,VH和 CH1 通过重叠 PCR 进行拼接获得 Fd。再通过重叠 PCR 分别拼接 Fd 和λ 或κ片段,获得Fab。PCR 的反应条件为:94 ℃预变性 5 min;94 ℃变性 30 s,52 ℃退火 30 s,72 ℃延伸1 min,30 个循环;72 ℃延伸 5 min。PCR 产物用琼脂糖凝胶电泳纯化回收备用。
1.2.3 Fab 噬菌体抗体库的构建 Fab 基因及 pCANTAB5H 载体质粒分别用I 酶切,产物分别回收后进行连接。连接物再经脱盐回收,电转化感受态 TG1 细菌,电转化后迅速加入 2YT 培养基,37 ℃振荡培养 1 h,取 100 μl 经 10 倍稀释后铺 2YT 平板检测库容,余下接入 250 ml 的 2YT 液体培养基(含 100 μg/ml 氨苄青霉素、1% 葡萄糖)中,37 ℃培养 6 h,加入辅助噬菌体侵染 1 h,去除培养基,接入 1 L 新 2YT 液体培养基(含 100 μg/ml 氨苄青霉素、50 μg/ml 卡那霉素)中,30 ℃培养过夜。次日从培养液中以 PEG 沉淀噬菌体,加入甘油保存于–80 ℃。
1.2.4 噬菌体抗体库的筛选 抗体库筛选参照文献[9],将人重组 CD276 用 50 mmol/L 碳酸氢钠(pH 10.0)包被 ELISA 板,PBS/2% 牛奶封闭后加入噬菌体库,室温缓摇 1 h,经 PBS/0.1% Tween 洗 6 次,酸性洗脱液(0.1 mmol/L HCl-Gly,pH 2.2,0.1% BSA)洗脱结合的噬菌体,再用 1 mol/L Tris-HCl(pH 9.1)中和洗脱液,测滴度,扩增噬菌体用于下一轮筛选。共进行 3 轮筛选,后 2 轮筛选过程同第一轮,但减少包被抗原量,并且逐渐增加 Tween 浓度,分别为 0.3%、0.5%。
单克隆噬菌体的筛选参考文献[10],挑取96 株经 3 轮筛选获得的阳性克隆,接种于 100 μl 含 100 mg/L Amp 的 2YT 培养液中,37 ℃培养至600= 0.3 ~ 0.4,加入 25 μl 辅助噬菌体进行侵染,然后加入 25 μl 含300 mg/L Kan 的 2YT 培养液,37 ℃培养过夜。富集的各个噬菌体克隆采用 ELISA 方法进行特异性鉴定,每孔包被人重组 CD276 100 ng,加 100 μl 噬菌体,HRP 标记的鼠抗 M13 抗体显色。
1.2.5 Fab抗体的表达和纯化 参考文献[7],在 HB2151 细菌中可溶性表达 Fab 抗体,0.5 mol/L 的 IPTG 低温诱导表达过夜,利用渗透压休克法从细菌周质腔获取可溶性 Fab,镍柱纯化获得 Fab 蛋白,SDS-PAGE 鉴定纯度。
1.2.6 ELISA 分析 纯化的 Fab 抗体采用ELISA 方法进行特异性分析,每孔包被人重组 CD276、IGF1R、gp120 和 BSA 各 100 ng,加梯度稀释的 Fab 抗体,HRP 标记的鼠抗 Flag 抗体显色。
2 结果
2.1 Fd 段、λ 链、κ 链的 PCR 扩增及 Fab 组装
PCR 分别扩增 IgM 重链可变区基因、轻链 λ、轻链 κ 基因。分别获得 IgM 和 IgG 的 6 种重链可变区基因,9 种轻链 λ 基因,6 种轻链 κ 基因,片段大小如图 1A 所示。重链可变区与 CH1 组建 Fd 片段,最后与轻链 λ 或 κ 基因通过重叠 PCR 构建成 1.6 kb 的 Fab 片段(图 1B)。
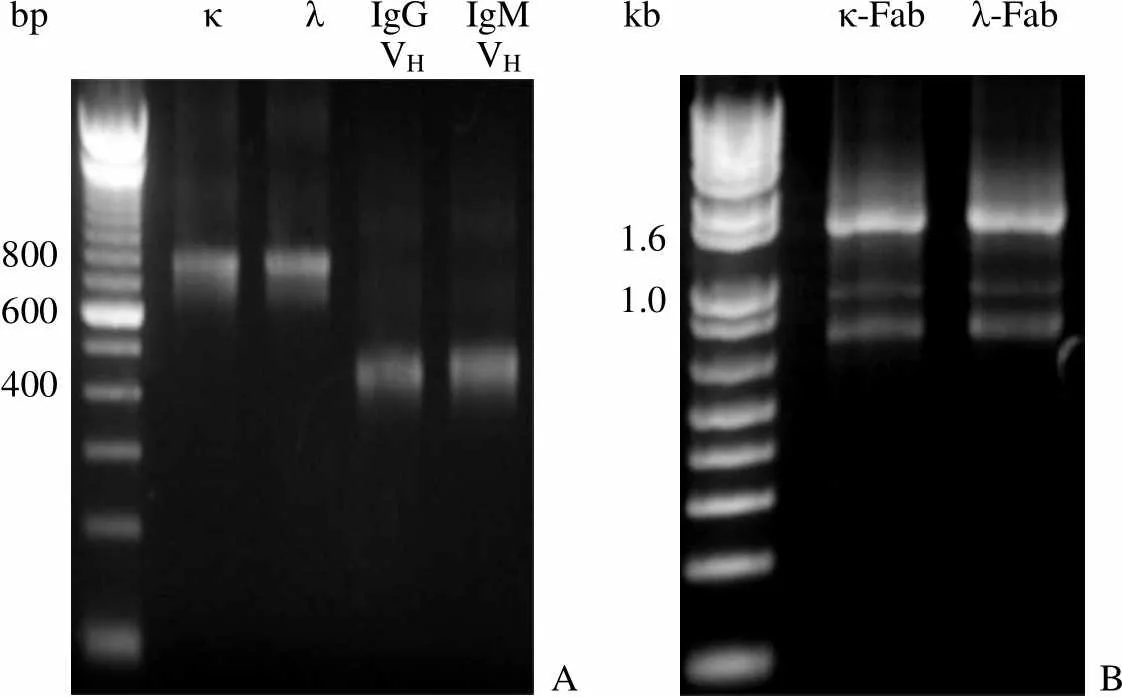
bpκ λ IgG IgMVH VH kbκ-Fab λ-Fab 800600 400A 1.61.0 B
Figure 1 PCR amplification of Fd, λ and κ fragments (A) and assembly of Fab fragments by overlapping PCR (B)
2.2 Fab 噬菌体抗体库的构建
胶纯化的 Fab 产物经I 酶切、回收纯化后与 pCANTAB5H 连接,并电转化至 TG1 菌中,构建 Fab 抗体库。根据转化产物在 2YT 平皿上长出的克隆数,确定其转化子数为 5 × 1010。将电穿孔转化后的细菌扩大培养,以辅助噬菌体感染,得到含有噬菌体抗体的上清,经 PEG 沉淀浓缩后获得噬菌体抗体库。
2.3 CD276 特异性噬菌体的筛选
进行 3 轮筛选,第一轮筛选采用低筛选强度,获得所有与抗原结合的噬菌体克隆,目的是除去大部分与抗原无亲和力的噬菌体,所获得的大部分噬菌体结合强度较弱。三轮筛选中,噬菌体经过扩增,拷贝数明显增加,并且将去污剂 Tween 的浓度逐轮增大(0.1%、0.3%、0.5%),增加冲洗次数,逐步增加筛选压力。最终富集的噬菌体的富集指数提高到 100,说明已经成功地富集了与 CD276 具有高亲和性的噬菌体(表 1)。
在第三轮筛选富集到的噬菌体中,随机挑选 96 个噬菌体克隆进行扩增,对其与 CD276 抗原行 ELISA 测定,筛选到 11 个噬菌体克隆显示出与抗原的高结合性(图 2)。挑选特异性最好的阳性克隆用于进一步的测序鉴定。

表 1 CD276 筛选的富集效应
注:a:富集指数 =(后一轮 O/I)/(前一轮 O/I)
Note: a: Enrichment factor = (Roundn+1Output/Input)/(RoundnOutput/Input)
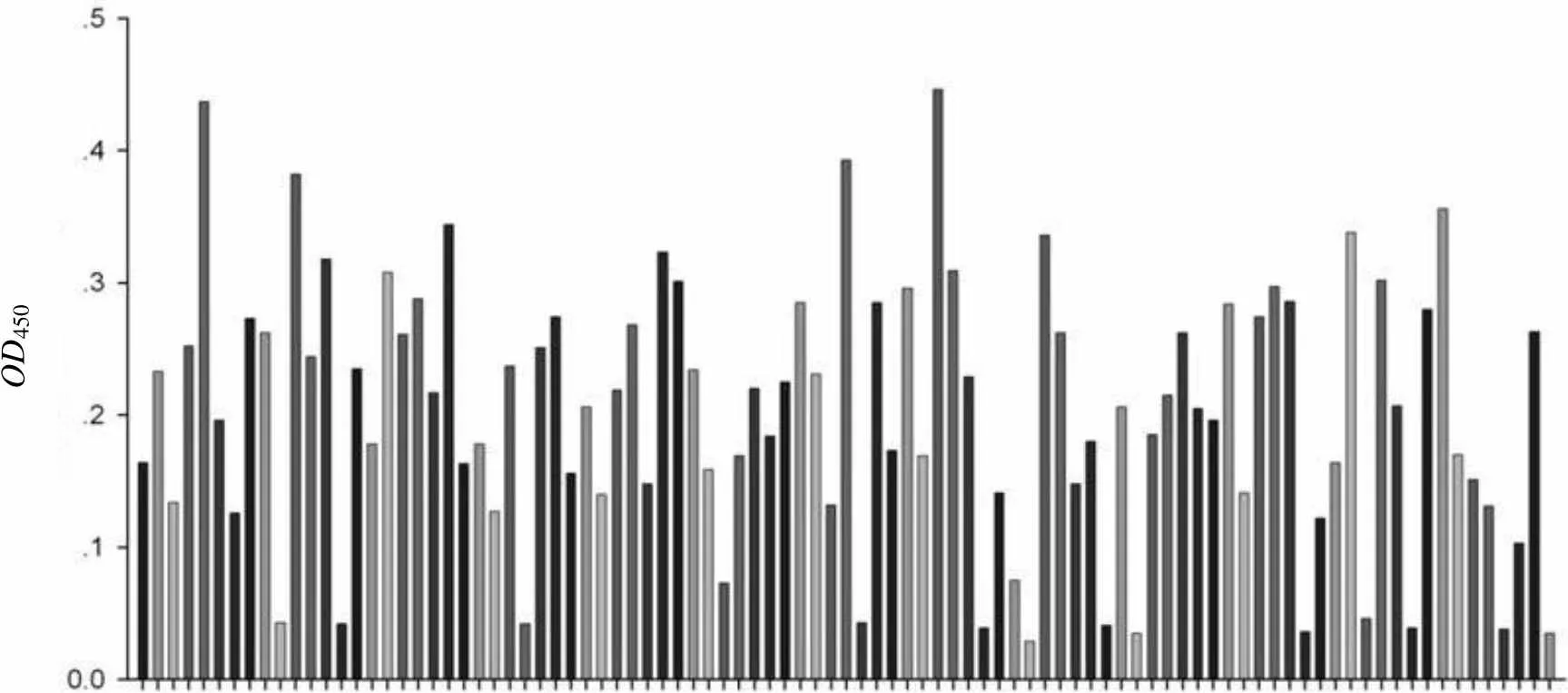
OD450
Figure 2 ELISA screening of anti-CD276 phage clones

kD 1 2 3 28 15A

OD4502.5 2.0 1.5 1.0 0.5 0 10 20 30 40 50 抗体浓度(μg/ml)Antibody concentration (μg/ml)B
1:蛋白分子量标准;2~3:抗 CD276 抗体 Fab
1: Protein marker; 2 -3: anti-CD276 Fab
图 3 抗 CD276 抗体 Fab 的表达纯化结果(A)和 ELISA 分析结果(B)
Figure 3 SDS-PAGE (A) and ELISA (B) of purified anti-CD276 Fab against different antigens
2.4 抗 CD276 抗体 Fab 的表达及结合性分析
测序结果显示所有克隆的序列是相同的,重链属于 V3 家族,轻链属于 VK1 家族。在 HB2151 细菌中可溶性表达 Fab 抗体,经镍离子树脂纯化后,SDS-PAGE 鉴定蛋白的纯度可达 95% 以上(图 3A)。为进一步证实抗体的特异性,CD276 抗原和对照蛋白(IGF1R、gp120 和 BSA)包被 ELISA 板,分析结果显示 Fab 抗体可以特异性地结合 CD276 抗原,不与其他非相关蛋白结合(图 3B)。
3 讨论
噬菌体抗体库技术以 PCR 技术和噬菌体展示技术为基础,通过构建噬菌体抗体库经抗原筛选得到特异性抗体成为获得人源抗体分子的有效途径[11]。抗体库技术利用抗原即可直接从非免疫抗体库中得到全人源特异性抗体,并且能筛选到针对该物种自身抗原的抗体,克服了难以用杂交瘤技术的障碍。被广泛地运用于实验室研究、疾病的诊断与治疗。
从天然抗体库中成功筛选出相应抗体,关键是保证抗体库中的抗体基因具有足够数量及多样性[12]。本研究收集 10 位健康人外周血分离淋巴细胞,保障抗体基因的全面和多样性,为成功地构建大库容量的抗体库提供了基本的保证。为了尽可能全面地扩增出抗体家族各成员的抗体片段,选择扩增了所有 IgG 和 IgM 的重链家族,尽管在人体内轻链 λ 链的抗体仅占 40%,为了在扩增过程中减少遗漏,同时也扩增了包括 κ 和 λ 在内的所有轻链成员。从理论上讲,Fab 片段的结构与天然抗体的抗原结合部位最为相似,因此选取噬菌体展示的抗体形式。有报道认为天然抗体库 1010的库容就可能包容所有的抗体[13],本研究构建的抗体库容量为5 × 1010,可以保证得到高特异性的人源抗体。
CD276 是新近发现的共刺激分子 B7 家族的一个新成员,在 T 细胞介导的免疫反应中起重要的调节作用,在正常组织、细胞中不表达或极低表达,高表达于多种肿瘤组织中[6]。以CD276 作为靶抗原,对 Fab 噬菌体抗体库进行了 3 轮吸附-洗脱-扩增的淘选富集。通过测定各级库的滴度和洗脱下来的噬菌体的产出率,计算噬菌体抗体库得到了约 10 倍的富集,表明 CD276 对所构建的噬菌体抗体库表面呈现抗体的噬菌体起到了淘选富集作用。经过 96 孔的单克隆 ELISA 分析和可溶性表达,获得高纯度的 Fab 抗体用于亲和性分析。最终鉴定了一个 Fab 抗体可以与 CD276 高亲和性结合,这些结果证实从 Fab 噬菌体抗体库中获得特异性人源抗体的可行性。
本研究通过提取总 RNA 进行 RT-PCR 等生物学技术初步获得了一个大容量的人源抗体库,并且初步筛选得到一个抗肿瘤抗原 CD276 的单克隆抗体,为进一步的研究和应用提供实验基础。
[1] Steinberger P, Majdic O, Derdak SV, et al. Molecular characterization of human 4Ig-B7-H3, a member of B7 family with four Ig-like domains. J Immunol, 2004, 172(4):2352-2359.
[2] Zhang GB. The role of human B7-H3 in inflammatory responses and tumor immune escape of lung cancer. Suzhou: Soochow University, 2012. (in Chinese)
张光波. 人共信号分子B7-H3在炎症反应及肺癌肿瘤免疫逃逸中的作用机制研究. 苏州: 苏州大学, 2012.
[3] Coyle AJ, Gutierrez-Ramps JC. The expanding B7 superfanuly: increasing complexity in costimulatory signals regulating T cell function. Nat Immunol, 2001, 2(3):203-209.
[4] Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Annu Rev Immunol, 1996, 14:233-258.
[5] Chen L. Co-inhibitory molecules of the B7-CD28 family in the control of T-cell immunity. Nat Rev Immunol, 2004, 4(5):336-347.
[6] Podojil JR, Miller SD. Targeting the B7 family of co-stimulatory molecules: successes and challenges. BioDrugs, 2013, 27(1):1-13.
[7] Zhao Q, Chan YW, Lee SS, et al. One-step expression and purification of single-chain variable antibody fragment using an improved hexahistidine tag phagemid vector. Protein Expr Purif, 2009, 68(2): 190-195.
[8] Shui X, Huang J, Li YH, et al. Construction and selection of human Fab antibody phage display library of liver cancer. Hybridoma (Larchmt), 2009, 28(5):341-347.
[9] Shen BF, Chen ZN, Liu MP. Recombinant antibodies. Beijing: Science Press, 2005. (in Chinese)
沈倍奋, 陈志南, 刘民培. 重组抗体. 北京: 科学出版社, 2005.
[10] Zhao Q, Wong PF, Lee SS, et al. Generation of anti-idiotype scFv for pharmacokinetic measurement in lymphoma patients treated with chimera anti-CD22 antibody SM03. PLoS One, 2014, 9(5):e96697.
[11] Tohidkia MR, Barar J, Asadi F, et al. Molecular considerations for development of phage antibody libraries. J Drug Target, 2012, 20(3): 195-208.
[12] Beerli RR, Rader C. Mining human antibody repertoires. MAbs, 2010, 2(4). [Epub ahead of print]
[13] Adams GP, Schier R. Generating improved single-chain Fv molecules for tumor targeting. J Immunol Methods, 1999, 231(1-2):249-260.
Construction of a large human Fab phage display antibodies library and screening of phage antibodies against CD276
LIU Jie, CHEN Jun-heng, XU Tian-shu, WANG Li-ping, ZHAO Qi
To construct a large human Fab antibodies library and to screen and identify the anti-CD276 phage antibodies from the library.
Total RNA was extracted from peripheral blood lymphocytes of 10 healthy donors, and the Fab antibody genes were amplified by RT-PCR. The amplification products were sequentially cloned into phage vector pCANTAB5E to construct a human Fab phage antibodies library. Antibodies against CD276 were screened using immobilized antigen. After three rounds of screening, 96 randomly selected clones were identified by phage-ELISA to select specific ones with high affinity for CD276.
A large human Fab phage-display library consisting of 5 × 1010members was successfully constructed. From the phage library, we obtained eleven positive clones which had specificity and binding reactivity towards CD276. By sequencing, all of the 11 clones had the same sequences, and their specificity was confirmed by ELISA.
We successfully constructed a large human Fab phage antibodies library and isolated the specific human anti-CD276 Fab antibodies, which provided an experimental foundation in future study and application.
Immunoglobulin Fab fragments; Phage display antibody library; CD276
ZHAO Qi, Email: qi.zhao@siat.ac.cn
10.3969/cmba.j.issn.1673-713X.2014.04.005
诺和诺德-中科院专项研究经费(NNCAS-2013-9)
赵琦,Email:qi.zhao@siat.ac.cn
2014-06-13
Author Affiliations: Shenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenzhen 518055, China (LIU Jie, CHEN Jun-heng, XU Tian-shu, ZHAO Qi); College of Life Science, Jilin University, Changchun 130012, China (CHEN Jun-heng, XU Tian-shu, WANG Li-ping)
