Effects of selective logging on tree diversity and some soil characteristics in a tropical forest in southwest Ghana
2014-10-18AlexAsaseBismarkAsiatokorKwasiOforiFrimpong
Alex Asase • Bismark K. Asiatokor • Kwasi Ofori-Frimpong
Introduction
Most of the remaining forests in West Africa have been logged to some degree and about 30% of forests in Central Africa are under logging concessions (Laporte et al. 2007). Logging of tropical forests is a significant source of revenue generation (van Gemerden et al. 2003) in West Africa. It is also a major cause of deforestation in the sub-region (Norris et al. 2010). In Ghana, it is estimated that over half of the country’s 214 forest reserves,enclosing 1.8 million ha forest have been selectively logged(Hawthorne and Abu-Juam 1995). The consequences of logging often remain visible decades later (Hawthorne et al. 2012).
The effects of logging on plant diversity have been well documented and are highly variable due to differences in harvest intensity and logging practice as well as site differences (Kuusipalo et al. 1996; Magnusson et al. 1999; Dickinson et al. 2000;Arets 2005; Park et al. 2005). Logging operations also affect soil characteristics (Pinard et al. 1996; Hutchings et al. 2002). The effects of logging on plant diversity (Hawthorne 1993; Kouamae et al. 2004; Swaine and Agyeman 2008; Onyekwelu et al. 2008)and soils (Abeberese and Kyereh 2006) are well studied in West Africa. However, no studies have addressed the effects of logging disturbances on tree diversity and soil properties. This information could assist strategic planning and management for sustainable timber production and ecological functioning (Hawthorne et al. 2012).
This report informs on the tree diversity and soil characteristics in relation to logging disturbances in a tropical moist forest in the Bia Conservation Area in southwest Ghana, West Africa.The Bia Conservation Area has an interesting and chequered logging history (Anonymous 2001). Previous studies on the effects of logging on the vegetation in the area include the works of Hawthorne (1993) and Oteng-Yeboah (1995). According to Hawthorne (1993) most species appeared to be regenerating well in the log-damaged areas. The study of Oteng-Yeboah (1995)also indicated that despite the previous logging activities in the area there was an abundance of timber trees although the mostdesired timber species such as Entandrophragma spp. and Piptadeniastrum africanum were uncommon (Oteng-Yeboah 1995).The objective of this study was to investigate the effects of selective logging disturbances on tree diversity and soil characteristics 29−35 years after logging. It was also to compare the tree data in the current study with earlier tree data in the study area.
Materials and methods
Study area
The study area in the Bia Conservation Area was located in the Bia district, southwest Ghana at latitude 6°32′ N to 6°37′ N and longitude 3°02′ W to 3°08′ W, and covered 355.6 km2. The study area is made up of Bia National Park in the northern portion and Bia Resource Reserve in the southern portion. The vegetation in the study area falls within the moist semi-deciduous and the moist evergreen vegetation types of Ghana (Hall and Swaine 1981). Geologically, the area is made up of Birrimian rocks that are largely hidden by a variety of forest ochrosols that are moderately acidic (Hawthorne 1993). Rainfall is bimodal with more intense precipitation occurring between May and August, and September and October. There is often a dry spell between November and January. Annual rainfall ranges from 1500 to 1800 mm. Relative humidity is around 75% in the afternoon and 90%at night. Mean monthly temperatures are 24−28°C. Topography is generally flat in the study area (Anonymous 2001).
We sampled two types of forests, unlogged and post-logged in October, 2010. The post-logged forests were in logging gaps(former clear-cuts) which constitute most of the area affected by logging operations in Ghana (Hawthorne et al. 2012). New growth was found in the gaps. Timber was harvested 29 to 35 years earlier under the Ghana selective logging system (Hawthorne 1993; Anonymous 2001). We lack pre-logging data for the sites reported here as post-logging forest stands. Post-logged forests were situated mostly in Bia Resource Reserve and unlogged forests in Bia National Park. We used satellite imagery and field survey to identify the geographic locations of the three forest types.
Methods
In total, we sampled ten of 25 m × 25 m plots. The plot size followed the work of previous studies (van Gemerden et al 2003;Asase and Tetteh 2010). The plots were randomly distributed within each forest type with five in unlogged forest and five in post-logged forest. Within each of the 625 m2plots, all trees of diameter at breast height (DBH) ≥ 10 cm (1.3 m above ground)were individually identified and their DBH measured using a diameter tape. Species identification and nomenclature follow Hawthorne and Jodgkind (2006). Tree species were classified according to their shade tolerance / ecological guilds (Hawthorne 1995). Pioneers are species for which seedlings need sun to establish; non-pioneer light demanders (NPLD) are species that need gaps to develop beyond the sapling stage; and shade bearer guild consists of species that can persist and grow in understory shade at seedling and sapling stage (Hawthorne 1995).
Soils were sampled to a depth of 30 cm from two randomly selected spots in a 5 m × 5 m subplot demarcated within each 625 m2plot. The soil samples collected from each subplot were thoroughly mixed and passed through a 2-mm sieve and then analyzed. All soil analyses were duplicated and means were calculated. Soil pH was determined using the distilled water procedure measured at a soil solution ratio of 1:1 (McLean 1982). The semi-micro Kjeldahl digestion method was used to determine N(Bremer 1965). Soil available P was determined using Bray and Kurtz method (Bray and Kurt 1945) and exchangeable K was estimated using flame photometric methods (Bremer 1965). Soil bulk density was determined using the core method (Blake 1965)and percentage moisture was determined following Asase et al.(2012).
Data analysis
Species diversity was evaluated using Shannon-Wiener index(Hill 1973). Importance Value Index (IVI) of trees was calculated as the sum of their relative density, relative frequency and relative dominance (Curtis and McIntosh 1950). Student’s t-test was used to compare means to detect statistical differences in means after testing for normality using the Shapiro.test function in R®statistical software (Crawley 2007). The Kolmogorov-Sminov test was used to identify differences in DBH-class distributions between forest types. Pearson correlation quantified the relationship between tree diversity and soil characteristics. Statistical analyses were executed with R®version 2.7.2. (R Core Development Team 2009).
Results
General tree diversity
We recorded a total of 310 trees of 87 species. Tree species richness ranged from 8−26 per plot with a mean of 18.3. Mean density of trees was 488 and ranged from 160 to 656 trees / ha. The most important trees in unlogged forest were Corynthanthe pachycera (IVI = 42.2), Baphia nitida (IVI = 20.2) and Celtis mildraedii (IVI = 14.7). The most important trees in post-logged forest were C. mildraedii (IVI = 20.4), C. pachyceras (IVI = 18.5)and Dialium aubrevillei (IVI = 17.0). Information on the 50 most abundant species of trees is listed in Appendix 1.
Tree diversity in relation to forest types
A total of 144 trees of 60 species were recorded in unlogged forest. There were 177 individual trees of 77 species in postlogged forest. Species composition varied in terms of shade tolerance / ecological guilds (Fig.1). There were more pioneer species in the post-logged forest stands. Mean Shannon-Weiner index (p <0.05) was higher for the post-logged forest and varied significantly (p <0.05) between forest types (Table1). Meannumber of trees per plot (p >0.05) and basal area per plot(p >0.05) were similar in the forest types. The DBH size class distributions of individual trees in the two types of forests showed an inverted J-shape (Fig.2). Numbers of trees in the different DBH size classes were similar across forest types(Kolmogorov-Smirnov test, p >0.05).

Tables 1. Plant diversity and soil properties (up to 30 cm) in three forest types
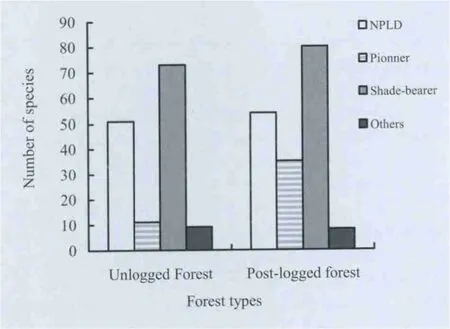
Fig.1. Ecological guilds of tree species in forest types.
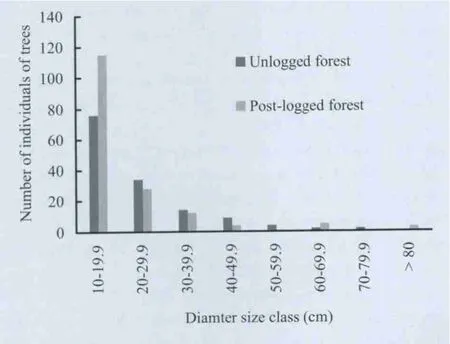
Fig.2. Comparison of DBH size-class distribution of trees (≥ 10 cm DBH) in forest types.
Variations in soil characteristics in forest types
Soil pH ranged from a mean of 4.8 in unlogged forest, to 5.3 for post-logged forest (Table1). The differences in soil pH between the forest types were insignificant (p >0.05). Similarly, there was no difference in soil bulk density between the forest types (p=0.08). The difference in percentage soil moisture content between the unlogged and post-logged forest stands was also significant (p = 0.023).
Soil nutrient status in the unlogged and post-logged forest was similar. No significant difference (p =0.62) in exchangeable K between post-logged and unlogged forest was detected. The difference in the available P between unlogged and post-logged forests was also insignificant (p =0.51). Total N content of soil was similar (p >0.05) in the two forest types.
Correlation analysis revealed significant relationships between basal area and soil pH (r = 0.58, p= 0.037), and basal area and total N concentration (r = 0.05, p ≤0.05). However, tree species richness, Shannon-Weiner diversity and density of trees were not significantly correlated with any of the soil characteristics.
Discussion
The Bia Conservation Area is an important conservation site surrounded by cocoa farmlands and serving as an international biological corridor between Ghana and Cote d’ Ivoire. Notwithstanding previous logging in the study area, we recorded numerous trees of species including C. mildraedii, S. triplochiton, P.angolense and S. glaucescens that are of economic value as timber). Our results on the general number of species and density of trees are very similar to that of Oteng-Yeboah (1995). In his study, a total of 364 trees belonging to 63 species were encountered in ten 25 m × 25 m quadrats similar to the 310 trees of 87 species recorded in this study. The most common species also included Corynthanthe pachycera, and C. mildraedii as recorded in this study.
In an earlier study on forest regeneration after logging in the study area, Hawthorne (1993) concluded that there was a sharp distinction in the vegetation and tree behavior between the adjacent unlogged forest (twilight zone) and areas directly damaged by logging operations. The botanical diversity of post-logged areas was greater than that of unlogged forest areas. Similarly,we recorded the highest tree diversity in post-logged forest areas.This result was expected because previous studies in other areas have indicated that logging generally leads to an increase in plant diversity due to colonization by trees (Swaine and Agyeman 2008) and weedy plants (Huno and Dang 2000).
In this study, the abundance of trees belonging to ecological guilds differed according to the two type of forest similar to thefinding of Hawthorne (1993). For example, pioneer trees were rare as small trees in the unlogged forest and more common in logged areas while NPLD and shade bearer trees were more abundant in unlogged forest areas. The significant difference in tree species ecological guild composition of the forest types was,however, not surprising because pioneer trees are more common in the post-logged forest due to the destruction of the forest canopy.
Forest structure in terms of tree dominance and DBH class distribution were similar between the forest types. The study of Hawthorne (1993) showed that some trees had attained 5 m height 1-2 years after logging although majority of the individuals were of lower stature. It is worth noting that the study area contains the largest and most impressive trees in Ghana whose canopy reaches in excess of 60 m in height (Anonymous 2001).In tropical forests, the basal area of trees >10 cm DBH is generally around 25–45 m2per ha (Huston and Wolverton 2009). Our forest types were structurally similar but differed in terms of tree species diversity and composition. Our study did not, however,include skid trails and loading bays which are more drastically affected by logging activities than are felling gaps. According to Hawthorne (1993) there may be no relationship between the composition of a young regenerating forest and the mature crop of the future. However based on the above comparisons, our findings on tree data from this study are generally very similar that of his study conducted about 17 years ago in the study area.
The soil characteristics in the study are comparable to those reported for other tropical forests (Onyekwelu et al. 2008). Soils in moist tropical forests are typically poor in nutrients and acidic in nature because of leaching by heavy rainfall. Generally, logging leads to compaction of soils (Alongi and de Carvalho 2008)and one of the indicators of soil disturbances is increase in bulk density from soil compaction (Hendrison 1990). We found no significant differences in soil bulk density by forest type, which could indicate that soils in the post-logged forest have recovered from compaction that might have occurred during logging. Logging can affect soil properties including nutrient loss (Alongi and de Carvalho 2008; Adekunle and Olagoke 2010). But we found that soil nutrients were similar in both unlogged and post-logged forest stands. Tropical rain forests have very efficient litter decomposition by which nutrient stocks are replenished. After a number of years of logging activity in the post-logged forest stands the loss of soil nutrients might have been replenished through litter decomposition and nutrient cycling.
This study has shown that selective logging had significant effects on tree diversity 29−35 years after logging but had no impact on the investigated characteristics of top soils. We recommend that future studies should examine the effects of logging disturbances on other soil physical and chemical properties that we did not investigate. Logging impacts on soil biodiversity should also be investigated to improve our knowledge of the consequences of logging on ecosystem function.
Acknowledgements
We are very grateful to the Wildlife Services Division of the Ghana Forestry Commission (FC) and the managers of the Bia Conservation Area especially Messer’s Alex N. Akwoviah, Richard Gyamfi Boakye, Fredrick Amankwa and Richard Ofori-Amafo (Park Manager) for permission and support during field work. Thanks to Messrs Patrick K. Ekpe and John Yaw Amponsah of the Ghana Herbarium at the Department of Botany, University of Ghana for their help in the identification of plants.Thanks to the Ecological Laboratory at the University of Ghana for laboratory analysis of soil samples.
Abeberese IK, Kyereh B. 2005. Regeneration of timber species following selective logging in a most-semi-deciduous forest in Ghana. Ghana Journal of Forestry, 17&18: 20−35.
Abeberese IK, Kyereh B. 2006. A post-logging assessment of some soil physical properties in a Ghanaian moist semi-deciduous forest. Ghana Journal of Forestry, 19&20: 54−62.
Adekunle VAJ, Olagoke AO. 2010. 'The impacts of timber harvesting on residual trees and seedlings in a tropical rain forest ecosystem, southwestern Nigeria. International Journal of Biodiversity Science, Ecosystem Services and Management, 6: 131−138.
Alongi DM, de Carvalho NA, 2008. The effects of small-scale logging on stand characteristics and soil biogeochemistry in mangrove forests of Timor Leste. Forest Ecology and Management, 255: 1359−1366.
Anonymous. 2001. Bia Management Area Plan.141pp.
Arets EJM. 2005. Long term responses of populations and communities of trees to selective logging in tropical rain forests in Guyana. Tropenbos-Guyana Series 13. Tropenbos-Guyana Programme, George-town, Guyana.
Asase A, Asiatokor BA, Ekpe P. 2012. Linkages between tree diversity and carbon stocks in unlogged and logged West-African tropical forests. International Journal of Biodiversity Science, Ecosystem Services and Management, 8(3): 217−230.
Asase A, Tetteh DK. 2010. The role of complex agroforestry systems in the conservation of forest tree diversity and structure in southeastern Ghana.Agroforestry Systems, 79: 355–368.
Blake GR. 1965. Bulk Density. In: C.A. Black (ed.), Methods of Soil Analysis:Part 1. Physical and Mineralogical Properties including Statistics of Measurement and sampling. Wisconsin, USA: Agron. Am. Soc. Agron.,Inc., Publisher Madison, pp. 374−377.
Bray RH, Kurtz LT. 1945. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci, 59: 39−45.
Bremer JM, 1965. Total Nitrogen. In: C.A. Black (ed.), Methods of Soil Analysis (part 2). Chemical and Microbiological Properties. Wisconsin,USA: Agron. Am. Soc. Agron., Inc., Publisher Madison, pp 1149−1178.
Crawley MC. 2007. The R book. West Sussex (UK) and New York: Wiley &Sons Ltd., p. 942.
Curtis JT, McIntosh RP. 1950. The interrelation of certain analytic and synthetic phytosociological characters. Ecology, 31: 434−455.
Dickinson MB, Wigham DF, Hermann SM. 2000. Tree regeneration in felling and natural treefall disturbance in a semi deciduous tropical forest in Mexico. Forest Ecology and Management, 134: 137−151.
Hall JB, Swaine MD. 1981. Distribution and Ecology of Vascular Plants in Tropical Rainforest: Forest Vegetation in Ghana. The Hague: Dr. W. Junk,p.383.
Hawthorne WD, Abu-Juam M. 1995 Forest Protection in Ghana: With Particular Reference to Vegetation and Plant Species. Gland (Switzerland) and Cambridge (UK): IUCN, p.203.
Hawthorne WD, Jongkind CCH. 2006. Woody plants of western African forests. A guide to the forest trees, shrubs and lianas from Senegal to Ghana. Kew (UK): Kew Publishing, Royal Botanic Gardens, p.1023.
Hawthorne WD, Sheil D, Agyeman VK, Abu-Juam M, Marshall CAM. 2012.Logging scars in Ghanaian high forest: towards improved models for sustainable production. Forest Ecology and Management, 271: 27−36.
Hawthorne WD. 1993. Forest regeneration after logging; findings of a study in the Bia South Game Production Reserve, Ghana. ODA Forestry Series No. 3 ODA, London, UK, p.52.
Hendrison J. 1990. Damage-controlled-logging in managed rain forest in Suriname. Wageningen, The Netherlands: Agricultural University, p.204.
Honu YAK, Dang QL. 2000. Responses of tree seedlings to the removal of Chromolaena odorata Linn. in a degraded forest in Ghana. Forest Ecology and Management, 137: 75−82.
Huston MA, Wolverton S. 2009. The global distribution of net primary production: resolving the paradox. Ecological Monographs, 79: 343−377.
Hutchings TR, Moffat AJ, French CJ. 2002. Soil compaction under timber harvesting machinery: a preliminary report on the role of brash mats in its prevention. Soil Use and Management, 18: 34−38.
Jans L, Poorter L, Van Rompaey SAR, Bongers F. 1993. Gaps and forest zones in tropical moist forest in Ivory Coast. Biotropica, 25: 258−269.
Kouamae FN, Bongers F, Poorter L, Traore D. 2004. Climbers and logging in the Foret Clasee du Haut-Sassandra, Cote-d’Ivoire. Forest Ecology and Management, 194: 259−268.
Kuusipalo J, Jafarsidik Y, Ådjers G, Tuomela K. 1996. Population dynamics of tree seedlings in a mixed dipterocarp rainforest before and after selective logging and crown liberation. Forest Ecology and Management, 81: 85−94.
Laporte NT, Stabach JA, Grosch R, Lin TS, Goetz SJ. 2007. Expansion of industrial logging in Central Africa. Science, 316: 1451.
Magnusson WE, De Lima OP, Reis FQ, Higuchi N, Ramos JF. 1999. Logging activity and tree regeneration in an Amazonian forest. Forest Ecology and Management, 113: 67−74.
McLean EO. 1982. Soil pH and lime requirements. In: Page, A.L, Miller,A.L.R. and Keeney, D.R. (eds.), Methods of soil analysis (Part 2); Chemical and microbiological properties (Second edition). Madison, WI: Agronomy Society of America & Soil Science Society of America, Inc, pp.199−244.
Norris K, Asase A, Collen B, Gockowksi J, Mason J, Phalan B, Wade A.2010. Biodiversity in a forest-agriculture mosaic–The changing face of West African Rainforests. Biological Conservation, 143: 2341–2350.
Onyekwelu JC, Mosandl R, Stimm B. 2008. Tree species diversity and soil status of primary and degraded tropical rainforest ecosystems in southwestern Nigeria. Journal of Tropical Forest Science, 20: 193–204.
Oteng-Yeboah AA. 1995. Biodiversity assessment of forest tree species in Bia National Park Ghana. In: Boyle, J.J.B, Boontawee, B. (eds), Measuring and monitoring biodiversity in tropical and temperate forests. Proceedings of an IUFRO Symposium held at Chiang Mai, Thailand. Centre for International Forestry Research, Bongor, Indonesia, pp.247−256.
Park A, Justiniano J, Fredericksen TS. 2005. Natural regeneration and environmental relationships of tree species in logging gaps in a Bolivian tropical forest. Forest Ecology and Management, 217: 147−157.
Pinard M, Howlett B, Davidson D. 1996. Site conditions limit pioneer tree recruitment after logging of dipterocarp forests in Sabah, Malaysia. Bitotropica, 28: 2−12.
Swaine MD, Agyeman VK. 2008. Enhanced tree recruitment following logging in two forest reserves in Ghana. Biotropica, 40: 370−374.
van Germerden BS, Shu GN, Olff H. 2003. Recovery of conservation values in central African rainforest after logging and shifting cultivation. Biodiversity and Conservation, 12: 1553−1570.

Appendix 1. Density, frequency, dominance and Importance Value Index (IVI) of the 50 most abundant trees in three forest types in Bia Conservation Area, southwest Ghana
杂志排行

Journal of Forestry Research的其它文章
- Development and evaluation of local communities incentive programs for improving the traditional forest management: A case study of Northern Zagros forests, Iran
- Fungal inoculation induces agarwood in young Aquilaria malaccensis trees in the nursery
- Spatial pattern analysis for quantification of landscape structure of Tadoba-Andhari Tiger Reserve, Central India
- Effects of harvesting intensities and techniques on re-growth dynamics and quality of Terminalia bellerica fruits in central India
- The impact of land afforestation on carbon stocks surrounding Tehran,Iran
- Herbaceous species diversity in relation to fire severity in Zagros oak forests, Iran