Callovian-Oxfordian hecticoceratins from western India: Their biostratigraphic and palaeobiogeographic implications
2014-09-27PinakiRoy
Pinaki Roy
Department of Geology, Durgapur Government College, Durgapur-713214, India
1 lntroduction *
The Oppelidae are the second most diverse ammonite groups after perisphinctids during the Middle-Upper Jurassic.Hecticoceratinae is the most diverse subfamily within oppelids and has a wide palaeobiogeographic and temporal distributions.It is taxonomically well studied in the Mediterranean (Elmi, 1967).This subfamily is known mostly by macroconchs only, although dimorphism has long been recognized (Zeiss, 1956).Elmi (1967)recognized microconchiate groups within Hecticoceratinae but did not establish dimorphic pairs at generic level.In one case, he recognized dimorphism within a genusi.e.,Brightias.st.(microconch)andBrightia(macroconch), and showed separate evolutionary lineages for each.
HecticocerasBonarelli is the type genus of the sub-family Hecticoceratinae.It has stratigraphic importance and is widely used to demarcate subzones and horizons.H.(Chanasia)michalskiLewinsky,H.boginensePetitclerc,H.proximumElmi andH.(Ch.)turgidum(Loczy)are used as zonal indices for the Michalski, Boginense, Proximum and Turgidum horizons respectively in the Subtethyan Province of Europe (Cariou, 1984).In Kutch, Hecticoceratinae is represented by eight genera ranging from the Middle Bathonian to Lower Oxfordian(see Spath, 1927-1933 and present study).The genusHecticocerasis represented by two speciesH.giganteumSpathandH.aff.turgidumLoczy which are now synonymised underH.giganteum, which ranges from the top of the Lower to basal Middle Callovian in Kutch (see Roy and Bardhan, 2007).I have also addressed sexual dimorphism withinHecticoceras giganteumbased on additional topotypes and other materials along with type specimens.The nature of sexual dimorphism appears to be differing from that of Oppelinae andProhecticoceras.The microconch is less strongly ornamented and lack primaries on the body chamber (Royet al., 2007).Thus it comes close toJeanneticerasZeiss, a microconchiate genus described by Elmi (1967)within the Hecticoceratinae.Within the scope of the present work, the taxonomic revision has been done for the Hecticoceratinae of Lower Callovian to Oxfordian of Kutch.Eleven biological species have been described under six genera and sexual antidimorphs are paired within the generaKheraitesSpath,PutealicerasBuckman,SublunulocerasSpath andBrightiaRollier.Kheraites crassefalcatumandK.igobilishave been paired as sexual antidimorphs;Putealiceras bisulcatuma smaller variant with a tricarinate venter has now been paired withPutealiceras trilineatum.Sexual dimorphism has also been established withinP.intermedium[male and female] andP.vijaya[male and female].Pseudobrightia dhosaensisan exclusive Lower Oxfordian genus described earlier by Spath(1927-1933)has now been redesignated asPutealiceras dhosaense.Sublunuloceras discoidesSpath, an Upper Callovian hecticoceratin described by Spath(1927-1933)with wide variation in size, I have some recognized smaller variants with a lappet now, so sexual dimorphism have also been addressed.A new species,Brightia callomonihas been described from the Upper Callovian-Oxfordian of Kutch, which is identical with Spath’s Brghtia sp.ind.and sexual antidimorphs are recognized.With these revised taxonomic data high resolution biostratigraphy has been addressed and palaeobiogeographic patterns have been analyzed using Brooks Parsimony Analysis (Brooks, 1985, 1990; Wiley, 1988a,1988b; Wileyet al., 1991)on the basis of the phylogeny of the subfamily Hecticoceratinae.
2 Geological setting
The Kutch Basin opened up with the initial fragmentation of Gondwana during the Middle Jurassic and sedimentation began soon due to repeated transgression and regression events (Biswas, 1977).The Jurassic rocks of Kutch range in age from Bajocian to Tithonian and cover nearly half of the area of Kutch, including both the mainland and three ‘islands’ separated by an intervening expanse of salt-flat known locally as the Rann (Figure 1).The regional structure consists of three parallel, NW-SE trending anticlines.The Jurassic rocks are best developed in the central anticline (Wynne, 1872; Rajnath, 1932),situated towards the north of the mainland.These anticlines are superimposed by a set of zones of culmination that crop out as topographical domes, as at Jara, Jumara,Keera and Jhura, which contain the world famous Callovian sequence.The domal outcrops are disturbed by Deccan Trap intrusions and their northern flanks are truncated by an east-west trending fault.The major lithostratigraphic units in ascending order are the Patcham, Chari,Katrol and Bhuj Formations (Mitra,et al., 1979; Krishna,1984).Among these, the Patcham and Chari Formations are present as inliers and are surrounded by the younger Katrol Formation.Despite frequent spatiotemporal facies changes, ammonites are abundant in these two older formations (Janaet al., 2005).
The specimens of the present study have been collected from the Jara, Jumara and Keera sections of the Chari Formation.Detailed lithostratigraphic successions of each section, along with the vertical distribution of the major ammonite species are shown in Janaet al., (2005, figures 3-5).Significant sedimentological works and detailed facies analyses are now available (Datta, 1992; Fürsich and Oschmann, 1993; Fürsichet al., 1994).The Chari Formation is represented by a heterolithic facies association consisting of shale, limestone and sandstone deposited in a mid-shelf environment.The Lower Callovian part of this formation is dominated by shale-limestone (packstone/wackestone)alternations while the base of the Middle Callovian consists of siliciclastics.
Hecticoceratin ammonites are distributed throughout the Patcham and Chari Formations spanning the Middle Bathonian to Oxfordian (see Spath, 1927-1933; Kanjilal,1980; and Figure 2, Roy, pers.obs.).
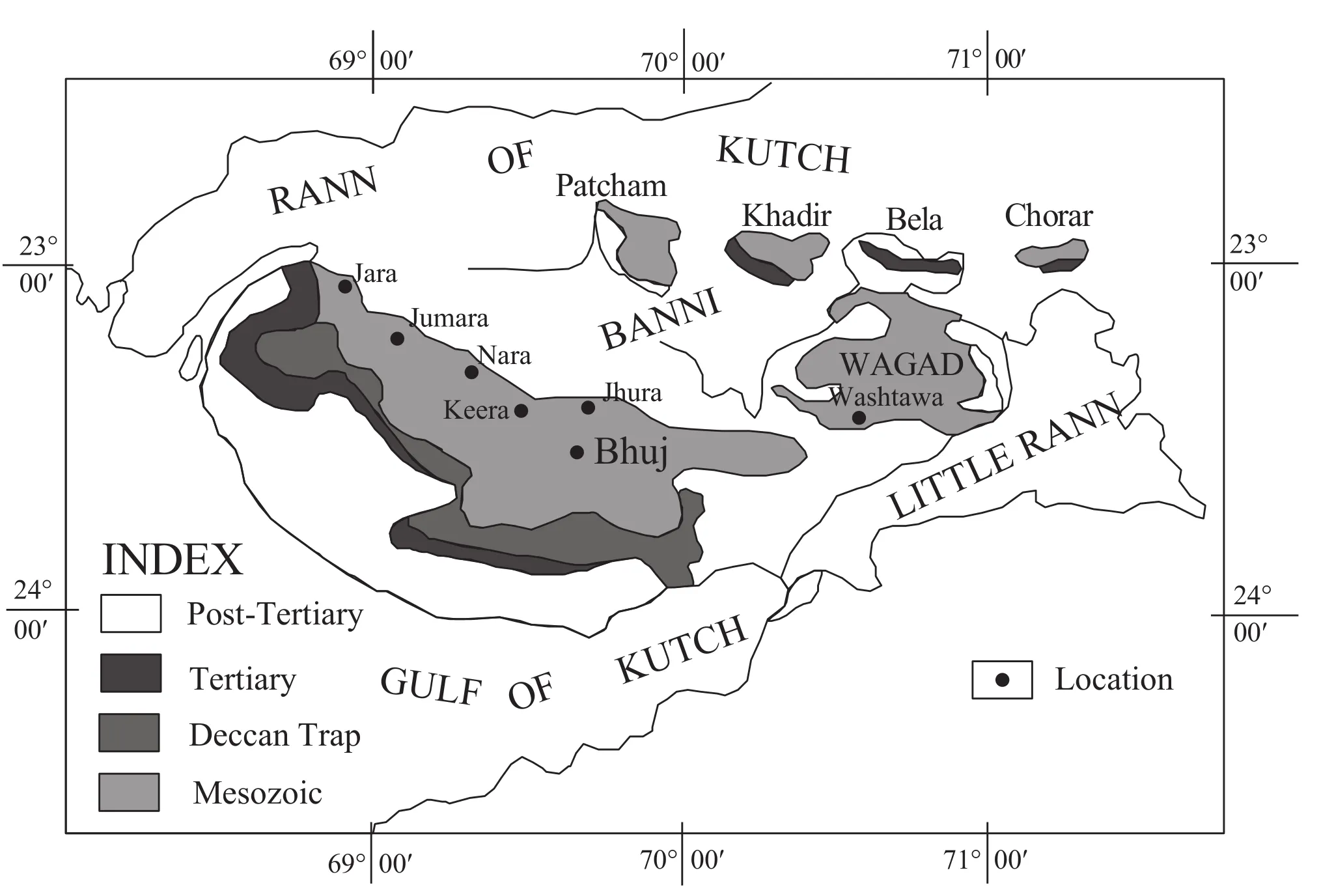
Figure 1 Geological map of Kutch, western India, with the geographic location of the study area.
3 Systematic palaeontology
3.1 Repository
The specimens studied in the present work are reposited in the following institutions:
1)Department of Geology, Durgapur Government College, Durgapur, West Bengal, India.
2)Geological Survey of India, Kolkata.
3.2 Abbreviations and symbols
The following abbreviations and symbols are used in the present study:
D = diameter of shell
H = height of whorl from umbilical margin
U = diameter of umbilicus
W = width of whorl
GSI = Geological Survey of India
JUM = Jadavpur University Museum
[M] = macroconch
[m] = microconch
* = asterisks against items in synonymies and measure-ments to indicate type specimens
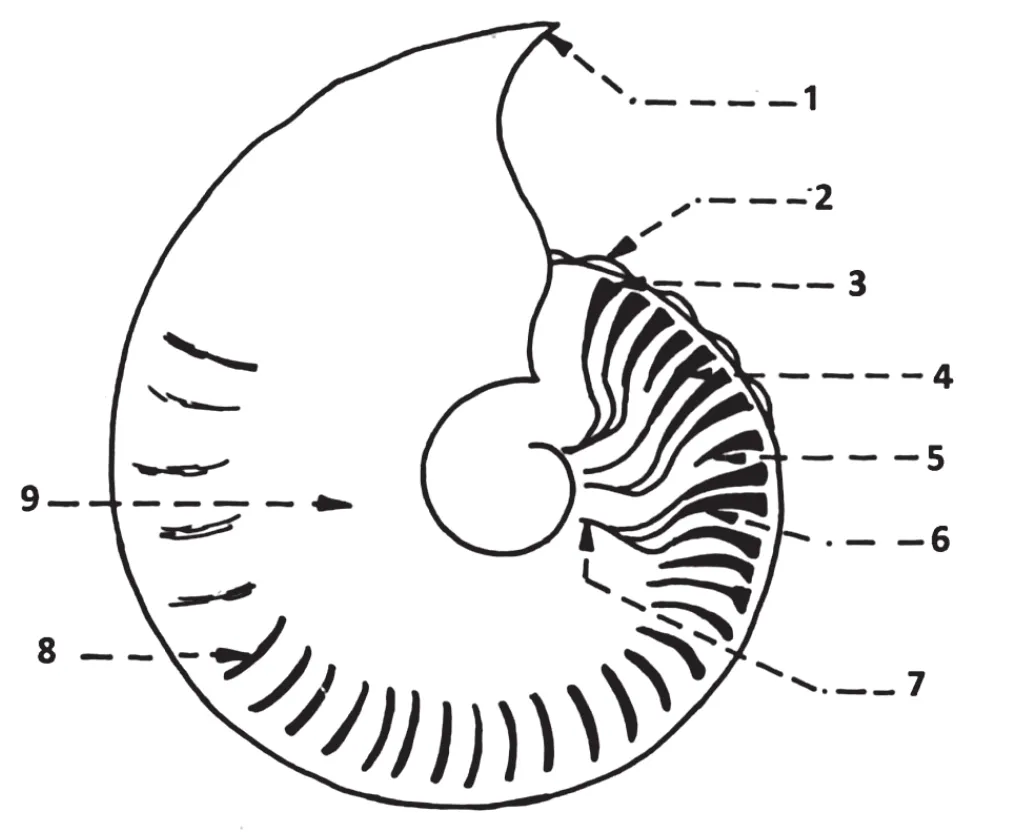
Figure 2 Ornamentation in the Subfamily Hecticoceratinae.1-Peristome margin; 2-Crenulated keel; 3-Ventro-lateral tubercle;4-Solitary rib; 5-Intercalatory rib; 6-Secondary rib; 7-Primary rib; 8-Reduced secondary rib; 9-Smooth inner flank.
ap.= aperture
b.c.= body chamber
E.Ph.= end-phragmocone
i.w.= inner whorl
Ph.= phragmocone
x = cross-marks in the plates indicate the position of adult end-phragmocone.
3.3 Systematic palaeontology
Superfamily Haploceratacea Zittel, 1884
Family Oppelidae Douvillé, 1890
Subfamily Hecticoceratinae Spath, 1925
1)GenusHecticoceras
Type SpeciesHecticoceras hecticum(Reinecke, 1818)
Hecticoceras giganteumSpath, 1927-1933
1875Harpoceras hecticum(Reinecke)Waagen; pars,p.61, pl.12, figures 3-5.
1893Hecticoceras hecticum(Reinecke)Bonarelli;pars, pp.78, 83.
1928Hecticoceras giganteumSpath; p.104, pl.16,figure 5.
1928Hecticocerasaff.turgidumLoczy, Spath; pars,p.105, pl.16, figure 4.
Comments: The genusHecticoceraswas earlier represented by two speciesH.giganteumSpath andH.aff.turgidumLoczy.Roy and Bardhan (2007)synonymized them underH.giganteumwhich ranges from the top of the Lower to basal Middle Callovian in Kutch.They had also addressed sexual dimorphism withinHecticoceras giganteumbased on additional topotypes and other materials along with the type specimens.The nature of sexual dimorphism appears to be different from that of Oppelinae andProhecticoceras.Microconch is less strongly ornamented and lack primaries on the body chamber.Thus it comes close toJeanneticerasZeiss, a microconchiate genus described by Elmi (1967)within Hecticoceratinae.
2)GenusKheraitesSpath, 1927-1933
Type speciesHarpoceras crassefalcatumWaagen, 1875Kheraites ignobilis(J.de.C.Sowerby)
Figure 3A-3L, Figure 4A-4K, Table 1
1840Ammonites ignobile, J.de C.Sowerby, in Grant,pp.297, 329, pl.23, figure 11
1875Harpoceras crassefalcatum,Waagen, p.70,pl.12, figures 6a, 7.
1875Harpoceras ignobile,Waagen (J.de C.Sowerby), p.69, pl.12, figures la, 2a, 2b only.
1884Harpoceras crassefalcatum,Waagen, Teisseyre,p.547.
1885Ludwigia crassefalcata,(Waagen)Haug, p.692
1987Harpoceras crassefalcata,Waagen; Noetling,p.22
1902Ammonites ignobilis,J.de C.Sowerby; Blake,p.35.
1912Harpoceras ignobile(non Sowerby ?)Smith,p.1352.
1913Harpoceras ignobile(non Sowerby ?)Smith,(c), p.423.
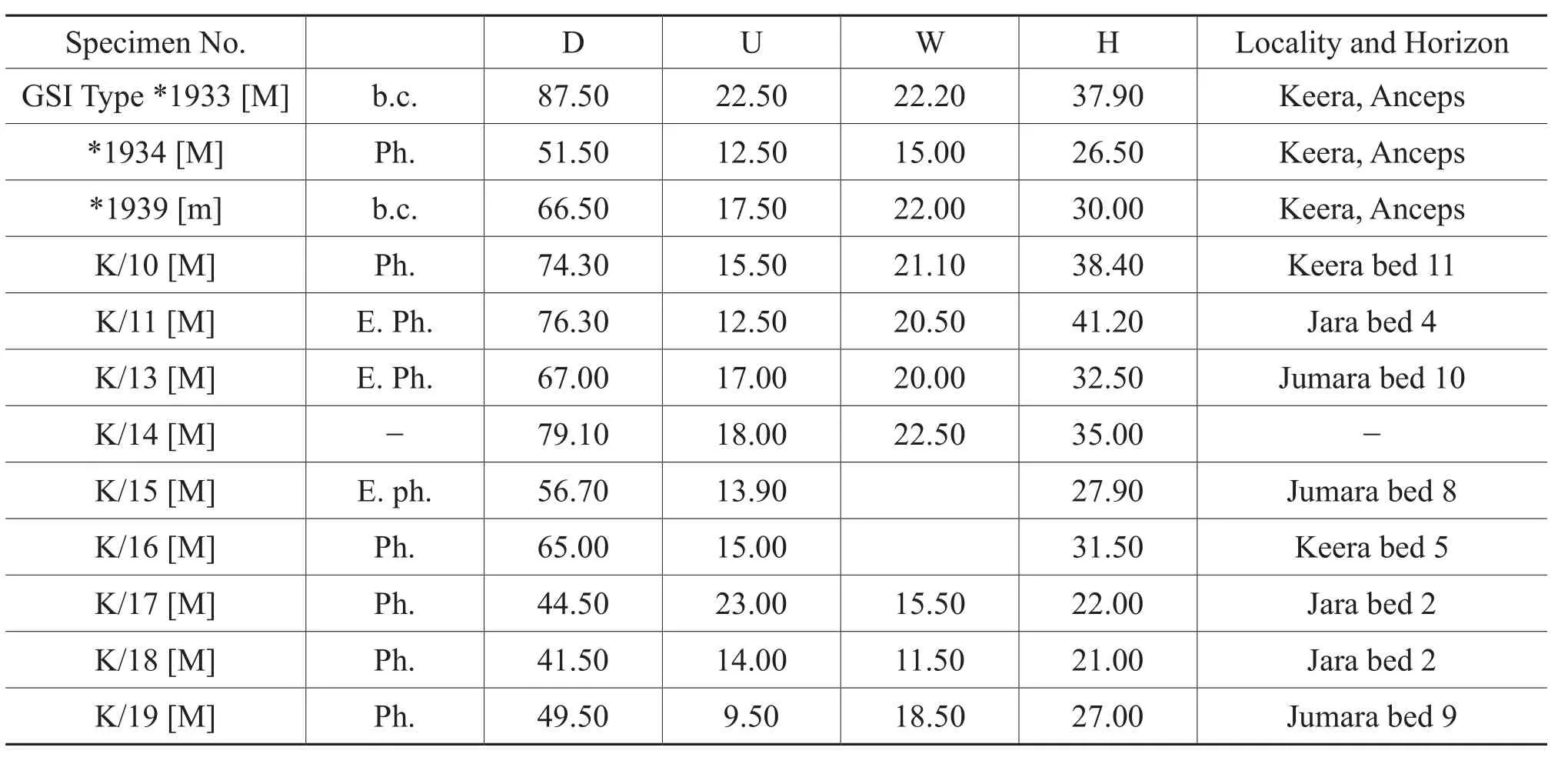
Table 1 Materials and dimensions of Kheraites ignobilis (J.de C.Sowerby)
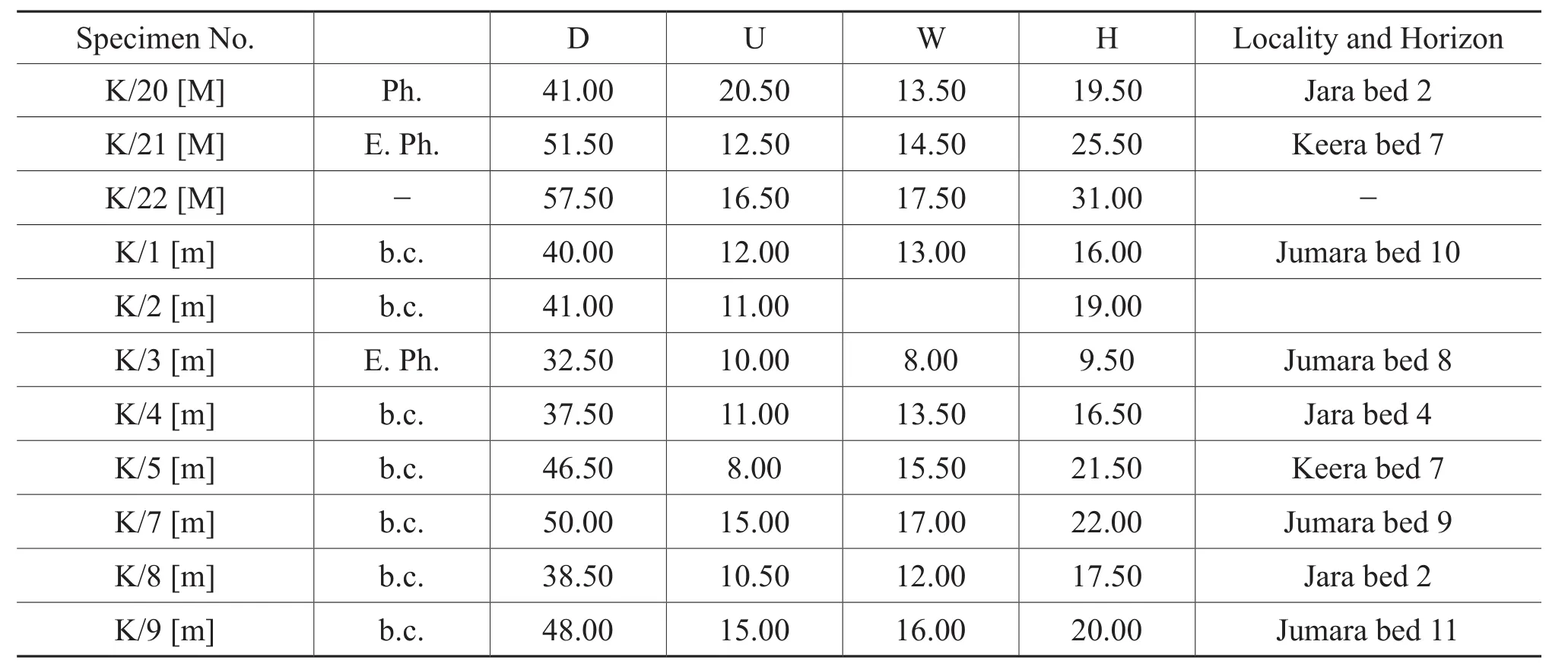
Table 1, continued
1913Harpoceras crassefalcatum,Waagen; Smith,(c), p.423
1925Kheraites crassefalcatum,Spath, (b), p.8.
1928Kheraites crassefalcatum,Spath, p.107.
1928Kheraites smithi,Spath, p.108, pl.16, figure 9.
1928Kheraites ignobilis,(J.de C.Sowerby)Spath,pl.15, figure 2.
Description [M]: Adult shell medium sized (D = 88 mm, GSI Type no.1933; pl.3, figure A-C)with adult phragmocone diameter ranging from 60 mm to 75 mm.Shell is fairly evolute (U/D = 0.21 to 0.51), compressed(W/H = 0.55 to 0.70); whorl section elliptical with rounded venter.
Early whorls are smooth up to a diameter of 10 mm,evolute with compressed whorl section (W/H = 0.69 -0.70).Later whorls exhibit coarse ribbing at the ventrolateral margin.These secondaries may continue up to the umbilical margin.Primaries become stronger at the early phragmocone (D = 30 mm)and are also weakly prorsiradiate.They bifurcate at one-third whorl height and form bullae at the furcation point.Weakly rursiradiate secondaries originating from these furcation points continue up to the ventro-lateral and form weak forward projections at the termination point.Occasional intercalatories or solitaries may be present in between the secondaries.With ontogeny, primaries become faint or disappear and the furcation point gradually shifts towards the mid flank (see Figure 3A-3C).Secondaries are flattened, coarse and concave towards the aperture and form clavi or projected bullae at the ventro-lateral margin.Spacing between secondaries gradually increases with growth.At the end of the phragmocone the inner flank is completely smooth (Figure 3A-3C), secondaries become more distant also lose strength;effaced clavi or bullae are only present near the ventrolateral margin.
Umbilicus is wide at the early stage of growth (U/D= 0.3)and at the phragmocone stage it become narrower(U/D = 0.22); at the end phragmocone or in body chamber it remains wider (U/D = 0.33)due to uncoiling of the body whorl.
The whorl section remains compressed throughout ontogeny, although the keel was not as prominent over the rounded venter in early ontogeny.In latter phragmocone the venter is weakly fastigate to flat with a raised keel at the siphonal area and on the body chamber the venter becomes rounded with faint trace of keel (Figure 3E).
Description [m]: The shell is small, maximum measured diameter being 66 mm (GSI Type 1939).The adult phragmocone diameter varies from 25 mm to 35 mm.Shell is evolute (U/D = 0.27 - 0.31), with subtrapezoidal whorl section (W/H = 0.72 - 0.84)and ornamented with falcoid strong ribs.
Primary ribs appear at the umbilical margin after an initial smooth stage 8-10 mm; although some variants show umbilical tubercles below 10 mm diameter (Spath’sK.varicosus).Primaries at early whorls are strongly prorsiradiate and form weak tubercles just near the umbilical margin.In inner whorls, these tubercles are at the point of bifurcation;secondaries thus are sharp, rectiradiate to weakly rursiradiate and terminate at the ventro-lateral margin by forming tubercles.In the phragmocone the furcation points become shifted towards the one-third whorl height and tuberculate primaries become elongated bullae.Secondaries becomerelatively blunt, concave towards the aperture and form clavi at the termination point at the ventro-lateral margin.At the adult phragmocone the primaries are strongly prorsiradiate or shifted away from their origin from the umbilical margin and become blunt.Secondaries become distant,blunt and weakly concave towards the aperture and they terminate at the ventro-lateral margin forming clavi which are projected towards the direction of growth.Irregular solitaries are present at the body chamber.
Venter is weakly fastigate to flat with a mid-ventral keel throughout ontogeny.In one variant the venter becomes wavy in the last part of the body chamber; it may be due to the projected termination of the secondaries at the ventrolateral margin or may be due to abrasion of the shell; only a preservational artifact.
Umbilicus is wide (U/D = 0.31)in all stages of growth,with an inclined umbilical wall and no prominent margin.Though, in some variants the umbilicus is relatively narrow at the body chamber (Figure 4C-4F).
Discussion: Waagen (1875)described nine specimens underHarpoceras ignobileJ.de.C.Sowerby and two specimens underH.crassefalcatum.Spath (1927-1933)redescribedH.crassefalcatumasKheraites crassefalcatumand also designatedK.crassefalcatumas the genotype.Spath (1927-1933)split Waagen’sH.ignobileintoK.smithiandK.ignobileon the basis of distant ribbing,greater whorl thickness and loss of keel in larger diameter.Spath (1927-1933)included the larger of Waagen’sH.ignobileintoK.smithi, although a faint keel continues in the body chamber fragment as evident from the study of the type specimen (GSI Type 15989).Study of type specimens in the repository of the Geological Survey of India, Kolkata along with additional collection, reveals that ribbing patterns shows continuous variation during ontogeny.As stated by Waagen (1875)both of these species have similar ornamentation up to 50 mm.All three species mentioned above have an initial smooth stage up to 10-15 mm diameter.K.crassefalcatumas described by Waagen (1875),and later by Spath (1927-1933), is smaller in size (Waagen’s two specimens show the beginning of body chamber at 50 mm and 66 mm diameter respectively)and strong ornamentation continues to the end.On the other handK.ignobilehas similar ornamentation and morphology up to 55 mm diameter as inK.crassefalcatum, although the body chamber (D = 88 mm)ofK.ignobilisshows disappearance of tubercles, rounding of whorls and an incipient keel.All the characteristics of larger diameter are macroconchiate characters; soK.crassefalcatumis the microconchiate counterpart of macroconchiateK.smithi— the larger specimen ofH.ignobileWaagen (1875).Spath(1927-1933)included Waagen’s two smaller specimens with Sowerby’sK.ignobilisdue to sigmoidal ribbing in early phragmocones and continuation of the keel till to the end.On the other hand, Spath (1927-1933)was hesitant to include the above two specimens of Waagen intoK.ignobilisdue to “less straight and blunter ribbing”.The present collection also shows continuous variation between slightly sigmoidal ribbing and less straight ribbing.Allthese suggest thatK.smithiandK.ignobilisare the same species and represent a different ontogeny.So all the three species of Spath are grouped here into a single species,K.ignobilis, and the name is retained due to page priority of Waagen’s (1875)description.
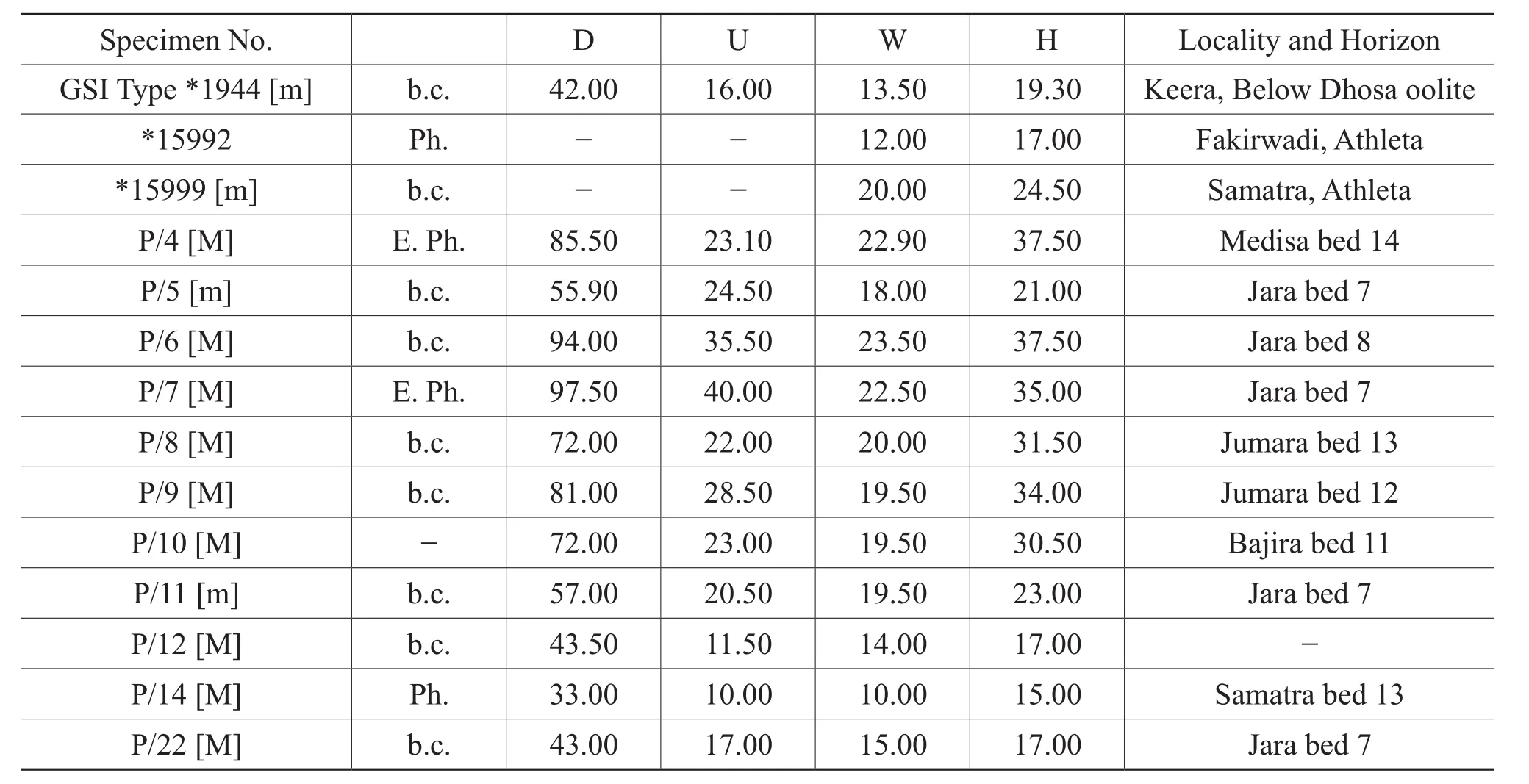
Table 2 Materials and dimensions of Putealiceras trilineatum (Waagen)
K.varicosusdescribed by Spath (1927-1933)is only a small body chamber fragment from the same horizon and locality as the present species.It has umbilical tubercles at an early stage of growth that is more or less similar development as inK.crassefalcatumof Spath (1927-1933);so it has also been included as a microconchiate variant ofK.ignobilis.
Spath’s (1927-1933)K.ferrugineusshows similar distant, course, alternate long and short ribbing as inK.varicosusand it differs by having smooth inner whorls up to 10 mm diameter.On the other hand, it shows similar ornamentation as inK.crassefalcatumbut it has an acute periphery unlike the latter; although the keel is more prominent in the outer whorls as found in the true microconchs ofK.ignoblis.
3)GenusPutealicerasBuckman, 1922
Type speciesAmmonite putealisLeckenby, 1859
i.Putealiceras trilineatum(Waagen)
Figure 5A-5K, Figure 6A-6G, Table 2
1875Harpoceras trilineatumWaagen, p.71, pl.13,figure 2a, 2b.
1885Ludwigia trilineata(Waagen)Haug, p.692.
1887Harpoceras trilineatum, Waagen, Noetling,p.22.
1924Hecticoceras?trilineatum(Waagen); Spath,p.5.
1924Hecticoceras?trilineatum(Waagen); Spath,p.24 (No.121 only)
1925Putealiceras trilineatum(Waagen); Spath, p.9.
1928Putealiceras trilineatum(Waagen); Spath,p.110, pl.12, figures 2a-2c, 10, pl.17, figure 10.
1928Putealiceras bisulcatum, Spath, p.114, pl.11,figure 12a, 12b; pl.16, figure 8a-8f.
1956H.(P.).trilineatum(Waagen), Zeiss, p.77,pl.3, figures 1, 2.
Description [M]: Shell evolute (U/D = 0.27 - 0.41),discoidal, large maximum diameter of partly preserved body whorl is 97.5 mm.Shell smooth up to diameter of 5 mm; afterwards strong rectiradiate primaries appear at the umbilical wall and form thick bullae at the furcation point near the umbilical margin.Secondaries are sharp,closely spaced and weakly falcoid to rectiradiate in nature.Secondaries go to the ventro-lateral margin and form elongated bullae which are projected towards the aperture.In early phragmocone (D = 50 mm)whorl height and width are nearly equal, although the whorl becomes higher than width during growth (W/H = 0.68 at D = 97 mm).In later phragmocone primaries become thicker and form bullae at the furcation point and still rectiradiate.Secondaries at the same stage of growth are still sharp, weakly falcoid to rectiradiate and the furcation point lies near the umbilical margin.Primaries become weaker or incipient in end phragmocone and in body chamber, although, weakly falcoid ribbing is still present and is blunt and distant.Secondaries at the end phragmocone are projected towards the aperture over the ventro-lateral margin.Strength of ribbing diminishes at the body chamber or the ribs become flattened.
In the early stage (D = 10 mm)the venter is rounded or weakly fastigate with the median keel accompanied by two faint auxlliary keels on both sides of the median keel.However, this tricarination may not be distinctly present in all growth stages as well as in all variants;e.g., in the GSI type 15992 the venter is distinctly unicarinate.The venter becomes broad in endphragmocone and the tricarinate continues into the body chamber (Figure 6F-6H), however, this tricarination is not present in the internal mould(Figure 6A-6E).
Umbilicus strongly wide (U/D = 0.3 to 0.41)all throughout ontogeny; umbilical wall rounded without any well defined margin.On adult body chamber the umbilical wall is steeper.
Description [m]: Shell stout, strongly evolute (U/D =0.36 to 0.44)and adult phragmocone diameter ranges from 15 mm to 43 mm.Earliest whorls are smooth up to 5 mm diameter, after that the whorls are closely spaced, strong,rectiradiate primaries appear at the umbilical wall and become thicker at the furcation point near the umbilical margin.Secondaries are also rectiradiate to weakly falcoid in nature and go up to the ventro-lateral margin where they form bullae, which are projected towards the aperture.There are some occasional solitaries and intercalatories present and the number of primaries and secondaries per half whorl is 10 and 14 respectively at 10 mm diameter.Primaries become coarser but still closely spaced and rectiradiate in the later phragmocone; they form elongated bullae at the furcation point.Secondaries are also rectiradiate but projected forward over the ventro-lateral margin and terminate near the auxiliary careena on the venter.The ornamentation continues up to the endphragmocone stage;after that primaries become less distinct and comparativelyblunt and their origin shifts towards the umbilical margin.
Venter broadly rounded with mid ventral keel accompanied by two faint auxiliary keels on either side of it at 10 mm diameter.In adult phragmocone the venter is also tricarinate with elevated mid ventral careena and more prominent auxiliary careena over an almost flat venter.Formation of two careena on either side of the mid ventral keel forms a sulcus along the venter on either sides of the keel.As the microconchs have a more tabulate venter than the macroconchs; the venter seems to be bisulcate.This tricarination continues up to the preserved body chamber.
Umbilicus is always wide (U/D = 0.36 to 0.44); wall rounded with no prominent margin.At the last stage of the adult body chamber, the umbilical wall is steeper, although the margin is still not prominent.
Discussion: Waagen (1875)compared the present species withKheraites crassefalcatum, but the similarities confined only to the outer whorls, where coarse ribbing appears but the keel becomes almost effaced.
P.vijayadescribed by Spath (1927-1933, pl.11, figure 7)is more evolute thanP.trilineatum(pl.12, figure 1)and inner whorls ofP.vijayaare more coarsely ribbed thanP.trilineatum.P.vijayaattains a tabulate periphery at the body chamber whereasP.trilineatumretains a fastigate periphery still at a larger diameter.
Young ofP.intermediumvar.samatrensehas similar straight ribbing as inP.trilineatum, but it has wider umbilicus.
InP.pseudodynastesas described by Spath (1927-1933, pl.14, figure 5), the inner whorls have similar rectiradiate ribbing as inP.trilineatum, and in this respect,Spath, 1927-1933 also intended to includeP.trilineatum;therefore I preferred it to be synonymized under the macroconch ofP.trilineatum.
According to Spath (1927-1933)one of the specimens ofP.bisulcatum(pl.11, figure 12a, 12b)is indistinguishable fromP.trilineatum, exceptP.bisulcatumhas finer,closer and more curved ribs whereasP.trilineatumis more compressed and has more coarsely ribbed inner whorls.But some variants ofP.bisulcatumshow coarse ornamentation and trilineate tabulate periphery.The specimens ofP.bisulcatumdescribed by Spath (1927-1933)are body chamber fragments; I have some small complete specimens which show similar ornamentation at early whorls withP.trilineatumandP.bisulcatumlike ornamentation at late stage of growth.So, I prefer to includeP.bisulcatumof Spath (1927-1933)within the microconch ofP.trilineatum.
Zeiss’s species (1956)from Jason Zone of Blumberg,Germany are relatively evolute and have strong ribbing,but it still has rectiradiate costation like the Kutch species.
ii.Putealiceras intermediumSpath
Figure 6H-6N, Figure 7A-7K, Table 3
1875Harpoceras dynastesWaagen, pars, p.66,pl.13, figures 7, 8 only.
1924Hecticoceras?lairense,Waagen; Spath,p.24 (Nos.122,124-25).
1928Putealiceras intermedium,Spath, p.112,pl.11, figure 9a, 9b, pl.12, figure 9a, 9b,pl.14, figure 4
Description [M]: Shell is large, adult phragmocone diameter ranges from 64 mm to 100 mm.Shell is relatively evolute (U/D = 0.31 to 0.36), compressed (W/H = 0.71 to0.83)and discoidal in shape.Early whorls are smooth up to about 10 mm diameter, then rectiradiate primaries appear near the umbilical wall which are sharp and closely spaced.Soon after the first stage of growth, primaries become a little coarser and are forwardly projected.Primaries are thicker at the point of furcation.Secondaries are rursiradiating and thus have a sickle-shaped appearance.Just before termination, secondaries are weakly projected towards the aperture near the ventro-lateral margin.There are about 9 primaries and 14 secondaries per half whorl at 24 mm diameter.Ornamentation becomes a little coarser at the later phragmocone (D = 46 mm); ribbing is falcoid to falcate; primaries thicker at the furcation point and secondaries form strong projected bullae at the termination point near the ventro-lateral margin.Primaries become strongly prorsiradiate at the endphragmocone and are weak or disappear from the inner lateral area in the body chamber;although secondaries continue at the body chamber they also lose strength with growth.
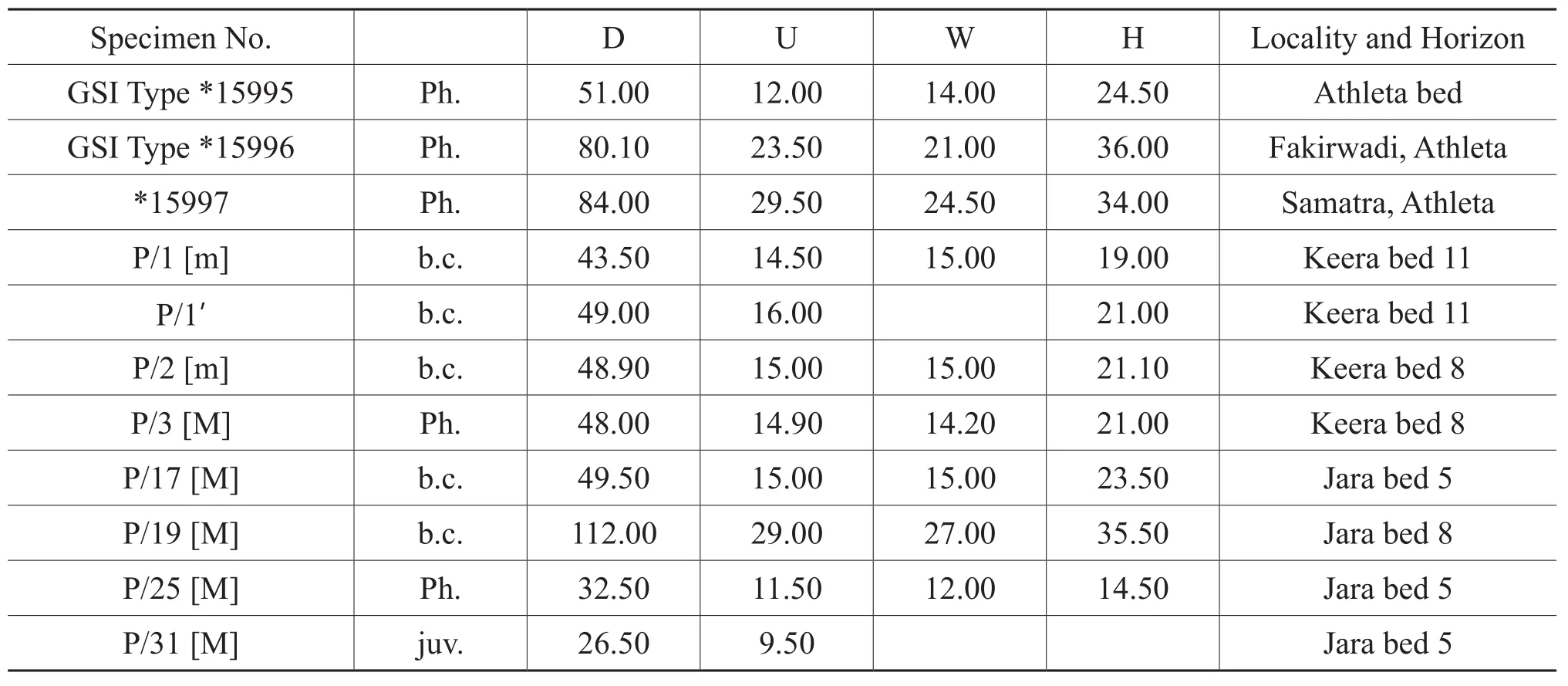
Table 3 Materials and dimensions of Putealiceras intermedium Spath
Venter is rounded to strongly fastigate in early ontogeny and becomes a little broad and unicarinate in the phragmocone.Keel continues up to the body chamber and the venter is broader and weakly fastigate in nature.
Umbilicus is wide in early whorls (U/D = 0.44 at D =25 mm)and continues to be wide in the later phragmocone but becomes narrow in the body chamber (U/D = 0.25 at D = 112 mm).Umbilical wall is rounded at early ontogeny but in adult body whorl it becomes steep with a sharp margin.
Description [m]: Shell relatively evolute (U/D = 0.3),small with discoidal whorl section.The complete body chamber diameter is 49.5 mm.Closely-spaced sharp primaries first appear after the smooth stage up to 5 mm diameter.Primaries are at first rectiradiate to weakly prorsiradiate and become thick at the furcation point at one-third whorl height.Like the primaries, secondaries are also closely spaced, rursiradiate and falcoid in nature.They form bullae weakly projected towards the aperture at the ventro-lateral margin.In phragmocone ribbing is relatively coarser, primaries are strongly prorsiradiate and secondaries become rursiradiate and concave towards the aperture.Secondaries project towards the aperture along the ventrolateral margin.There are 9 primaries and 17 secondaries per half whorl at the last whorl of the complete specimen.
Venter is rounded and unicarinate at the early whorls and comparatively depressed.At 21.5 mm diameter the whorl becomes compressed (W/H = 0.75), with strongly fastigate venter ornamented with an elevated keel.The venter becomes weakly fastigate to flat at the body chamber and keel continuous up to the end.
The umbilicus is wide (U/D = 0.39)at the early stage of growth (D = 21 mm)but relatively closer at the body chamber (D = 49.5 mm, U/D = 0.32).Umbilical wall is rounded in early ontogeny with no prominent margin.At the last part of the body chamber of the complete specimen the umbilical wall is vertical with a sharp margin.
Discussion: Spath (1927-1933)included Waagen’s(1875, pl.13, figure 6a, 6b)two small specimens ofHarpoceras dynastesinto his newly formed speciesPutealiceras intermedium.Spath (1927-1933)differentiated his species with additional specimens from Waagen’s species on the basis of whorl shape, which is more inflated and at the same time more distantly and more coarsely costate thanHarpoceras dynastes.The present species apparently shows similarities in whorl shape withP.trilineatum, but differs by a relatively compressed whorl section and involute coiling.P.intermediumshows great intraspecific variations,e.g., some variants of the present species have a similar inflated whorl section like Spath’s (1927-1933, pl.12, figure 9)P.intermediumvar.robustaand it is also rela-tively larger and evolute likeP.trilineatumbut unlikeP.trilineatumit has distant, coarse, strongly falcoid ribbing.On the other hand, some variants have indistinct ribbing in inner lateral areas as is found inSublunuloceras dynastes;but the present species differs by its comparatively evolute coiling and thickening of primaries at the point of bifurcation.
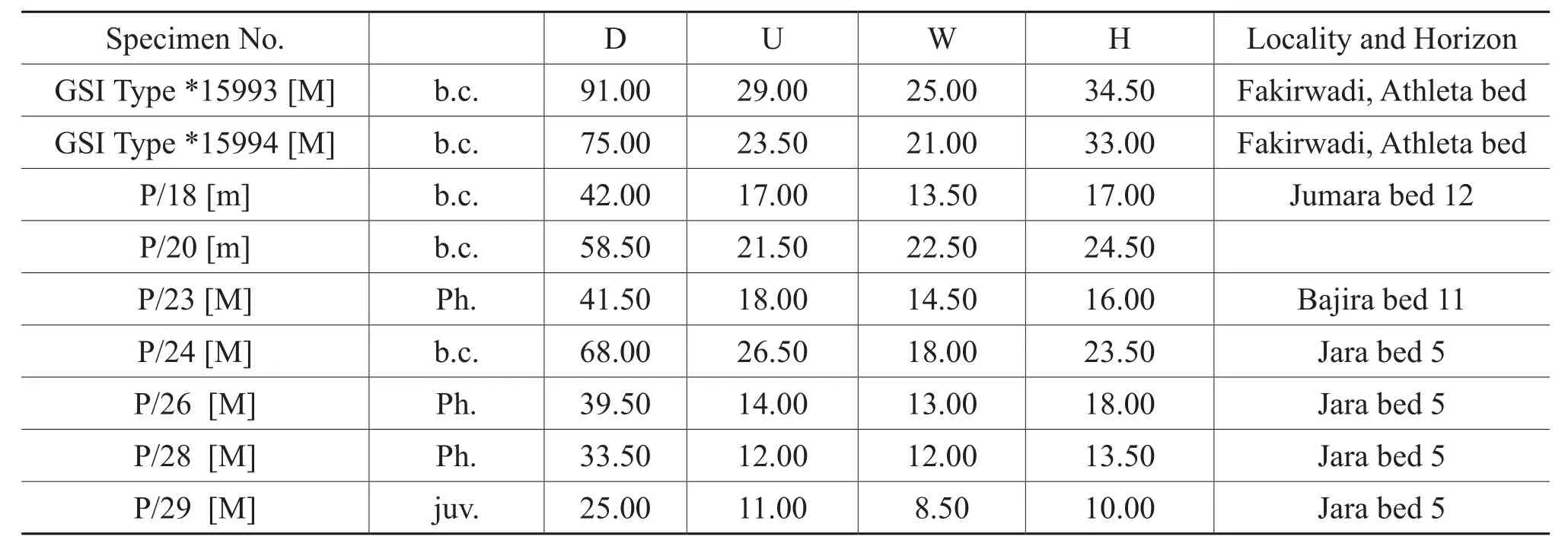
Table 4 Materials and dimensions of Putealiceras vijaya Spath
In bothP.vijayaandP.intermediumthe strength of ribbing diminishes or becomes blunt on the body chamber,so they may look alike, but in the present form the prominent keel continues along the mid venter even in the body whorl, whereas, inP.vijayaventer becomes more or less smooth in the body whorl.
iii.Putealiceras vijayaSpath
Figure 8A-8L, Table 4
1928Putealiceras vijaya, Spath, p.111, pl.11, figure 7, pl.12, figure 1a, 1b, pl.13, figure 9.
Description [M]: Shell large, end phragmocone diameter ranges from 71 mm to 73 mm (Geological Survey of India, Type no.15993 and 15994; Spath, 1927-1933; Pl.11, figure 7; Pl.12, figure 1).Shell is evolute (U/D = 0.32),compressed (W/H = 0.63 to 0.72)with elliptical whorl section.
Early whorls smooth up to about 10 mm diameter, then strong primaries appear near the umbilical margin, they are weakly prorsiradiate, sharp, distant and not always bifurcating in nature.They are falcoid, go up to the ventrolateral margin and form elongated bullae at their termination point.Intercalatories arise at the midflank and go up to the ventro-lateral margin and terminate there by forming bullae like projections.Primaries become more prorsiradiate, coarser and more distant with growth.In the phragmocone (D = 39.8 mm), the ribbing becomes more distant and stouter and the alternate long and short ribs continues,although occasional bifurcations are also present.In late phragmocone the ribbing is relatively straighter, robust in mid lateral and becomes blunt near the ventro-lateral margin.Secondaries are projected along the ventro-lateral margin towards the growth direction.At the later part of endphragmocone or in the body chamber the strength of ribbing diminishes greatly; only distant robust secondaries continue to the end of preserved body chamber.
Venter is rounded in early whorls, although compressed(W/H = 0.85 at D = 25 mm)with a prominent mid-ventral keel.In the endphragmocone or in the body chamber the venter becomes fastigate though the keel is not so prominent on the internal mould and the venter seems to be smooth.The whorl becomes comparatively depressed.
Umbilicus remains wide in all stages of growth.Umbilical wall is rounded with no prominent margin in early phragmocones.In body whorl the wall is gradual.
Description [m]: Shell strongly evolute (U/D = 0.43),robust, small-adult phragmocone diameter is 33.5 mm.
Strong primaries appear at the umbilical wall after an initial smooth stage of 3-4 mm.Ribbing rectiradiate and mostly solitary.Primaries when bifurcate form strong bullae at the furcation point just below the mid flank.Secondaries form thick elongated bullae at their termination point near the ventro-lateral margin.Ribbing becomes coarser and distant in the adult phragmocone; although they are still mostly rectiradiate and solitary.There are regular intercalatories in between solitaries.In adult body chamber, ribbing is falcoid and bifurcation becomes relatively frequent than in the earlier whorls.Primaries form thick, elongated bullae at the point of furcation near the mid-flank.In body chamber primaries appear near the umbilical margin instead of the wall.Secondaries are relatively falcoid and form strong bullate termination near the ventro-lateral margin.Ribbing becomes mostly blunt in the body chamber.
Venter is narrow in early whorls and becomes weakly fastigate to flat in the body chamber, a prominent midventral keel continues up to the end of the preserved body chamber.
Umbilicus is deep but wide and umbilical wall is gradual with no prominent margin.
Discussion: Spath (1927-1933)comparedP.vijayaandHecticoceras thirriaiPetitclerc (1921, pl.21, figure 10)with respect to blunt, straight ribbing, and loss of tuberculation at the later whorls; however, the ribs are not flattened inP.vijayaas in trueHecticoceras.
Spath (1927-1933)also comparedP.vijayawithP.trilineatumas the macroconch of both of them are of nearly the same size and both are evolute.P.vijayahas a more or less depressed whorl throughout whereasP.trilineatumhas an almost squarish whorl section in the inner whorls and becomes compressed in later stage of growth.P.vijayahas distant, coarse, rectiradiate ribbing;P.trilineatumshows closely spaced, weakly falcoid to rectiradiate ornamentation.Like Spath (1927-1933), I also have complete specimens ofP.vijaya, which shows a broad tabulate,smooth periphery at the body chamber;P.trilineatumhas a relatively fastigate, trilineate periphery.
P.intermediumdiffers fromP.vijayaby having a relatively involute and compressed whorl; closely spaced sharp, falcate ribbing with strongly fastigate venter and elliptical whorl section.
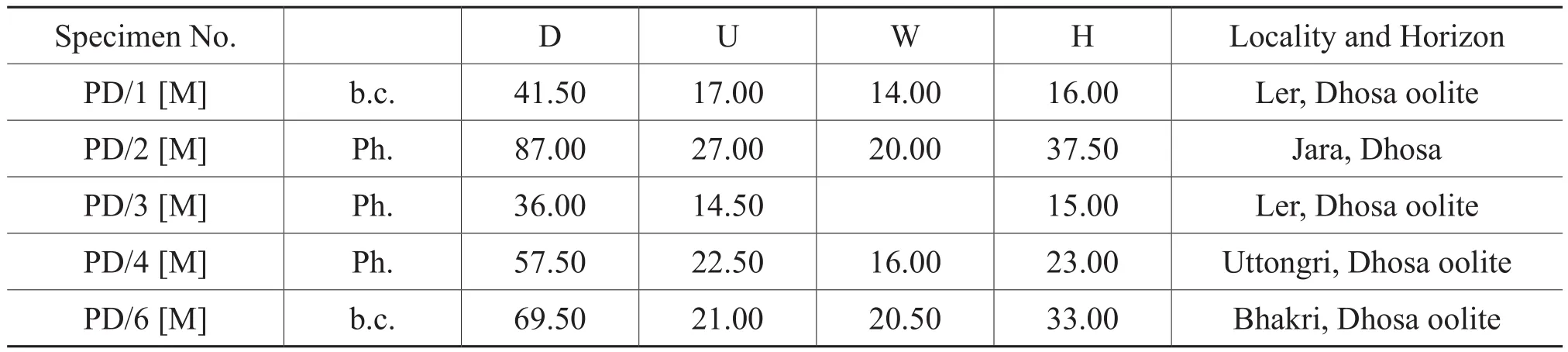
Table 5 Materials and dimensions of Putealiceras dhosaense n.sp.
P.punctatum(non Stahl)documented by R.Douvillé(1914, p.6, pl.1, figure1)shows similar loss of the blunt keel at the outer whorl, however, this species differs from the present form in its evolute inner whorls as well as in the very prominent ribs of the body chamber; the trueP.punctatum(Stahl)has not only less straight ribbing thanP.vijaya, but more compressed and prominent keeled whorls.
iv.Putealiceras dhosaense(Spath)
Figure 9A-9H, Table 5
1875Harpoceras punctatum, Waagen, p.62, pl.13,figure 9a, 9b.
1928Pseudobrightia dhosaense, Spath, p.116,pl.13, figure 10a, 10b.
1928Pseudobrightia subpunctata, Spath, p.116.
Description: Shell widely evolute (U/D = 0.30 to 0.41)compressed (W/H = 0.53 to 0.88), and moderately large with maximum adult phragmocone diameter 49 mm.
Shell is smooth up to 5 mm diameter, and then closely spaced primaries appear near the umbilical margin.Strength of primaries increases with growth.In the early phragmocone primaries become prorsiradiate and bifurcate at one-third whorl height; secondaries are rursiradiate in nature and form bullae at their termination near the ventro-lateral margin.Ribbing becomes more angulirursiradiate in later ontogeny, but secondaries become blunt and are projected towards the aperture at their termination along the ventro-lateral margin.In the body chamber the ornamentation loses strength, although continues up to the preserved end.
Venter is broadly fastigate with a blunt mid ventral keel at the early stage of growth.In the late phragmocone the venter becomes bisulcate in nature and the mid ventral blunt keel continues.
Umbilicus is broad in all stages of growth (U/D = 0.41 at D = 26 mm).In early stage, the umbilical wall is rounded with no prominent margin.In the body chamber the wall is relatively steep with a rounded margin.
Discussion: Spath (1927-1933)described the genusPseudobrightiafor the first time from Kutch and representedP.dhosaensisfrom the Lower Oxfordian (Dhosa Oolite bed)of Fakirwadi as the genotype, on the basis of a single fragmentary and completely septate specimen.The present author has some additional specimens from the same horizon and from different sections.P.dhosaensishas similar tricarinate periphery as inPutealiceras trilineatumandP.bisulcatum.The latter two have already been combined withinP.trilineatumin the present endeavour.Moreover,P.dhosaensisof Spath has stouter and evolute whorls as inPutealiceras.Spath (1927-1933)compared hisP.dhosaensiswith aPutealiceras(figured but not described by Spath, pl.16, figure 10)and it connectsP.dhosaensiswithP.trilineatumandP.bisulcatum.ButP.dhosaensisdiffers fromP.trilineatumandP.bisulcatumby having strongly falcoid and blunt ribs as also indicated by Spath (1927-1933).
Spath (1927-1933)described a new speciesP.subpunctataunder his new genus and he also included Waagen’s(1875)Harpoceras punctatumwithin it.H.punctatumhas high intraspecific variation in costation (Waagen, 1875)and it also varies during ontogeny; weak ribbing appears in early whorls and becomes stronger at the later phragmocone.The ribbing is relatively straighter compared toP.dhosaensisand the ribs do not form tubercles as also seen in some varieties ofPutealiceras trilineatum.Waagen’s(1875)specimen has the beginning of a body chamber preserved and Spath (1927-1933)compared a doubtful body chamber fragment (at D = 90 mm)with Waagen’s species as well as to the holotype ofP.dhosaensisand the transitional form toPutealicerassp.ind.(Spath, 1927-1933, pl.16, figure 10).All of them are similar in their peripheral aspects.So the present author, while revisiting the genusPeudobrightiawith additional specimens, has not found enough distinct morphological features to retain this genus separately fromPutealiceras.Hence, bothP.dhosaenseandP.subpunctatahave been synonymized under the present species.
4)GenusSublunulocerasSpath, 1927-1933
Type speciesHarpoceras lairenseWaagen, 1875
v.Sublunuloceras lairense(Waagen)
Figure 10A-10J
1975Harpoceras lairense, Waagen, p.65, pl.13,figures 3, 4.
1885Ludwigia lairensis(Waagen), Haug, p.691.
1893 (?)Harpoceras lairense, Waagen; Bonarelli,p.103.
1912Harpoceras lairense, Waagen; Smith,pp.1350-1352.
1913Harpoceras lairense, Waagen; Smith (c),p.422.
1928Sublunuloceras lairense, (Waagen), Spath,p.123, pl.11, figures 6a, 6b, 10; pl.14, figure 1; pl.17, figures 5, 6; pl.18, figure 8.
1928Sublunuloceras prelairense, Spath, p.122,pl.11, figure 5a, 5b.
Comments: Spath (1927-1933)described the species on the basis of 15 specimens, including Waagen’s four specimens.In the present study, I do not have any specimen similar in form to Spath’s forms which have rectiradiate ribbing in outer whorl and bending of moderately thick ribs in the body chamber.Spath designated one of Waagen’s forms (GSI Type 1945, here in Figure 12H-12J)as varietyS.lairensevar.planawhich shows peculiar flattening of the ribs, instead of thickening at the point of bifurcation.
Spath described a new speciesS.prelairenseand differentiated it from the present form only because of the presence of an acutely fastigate periphery, incipient keel without tendency of tricarination as shown in Waagen’s form.However, the ribbing shows peculiar bending and outer flattening as that of the present form, GSI Type 16007(Figure 12E-12G)does not show tricarinate venter and it may be regarded as an intraspecific variability.So in the present study, I have synonymized Sapth’sS.prelairensewithin the present species.
vi.Sublunuloceras discoidesSpath
Figure 11A-11H, Figure 12A-12I, Table 6
1928Sublunuloceras discoides, Spath, p.126,pl.12, figure 7a, 7b, pl.13, figure 1a, 1b,pl.15, figure 11.
1951Sublunulocerascf.discoides, Spath, Jeannet,p.66, pl.15, figure 4.
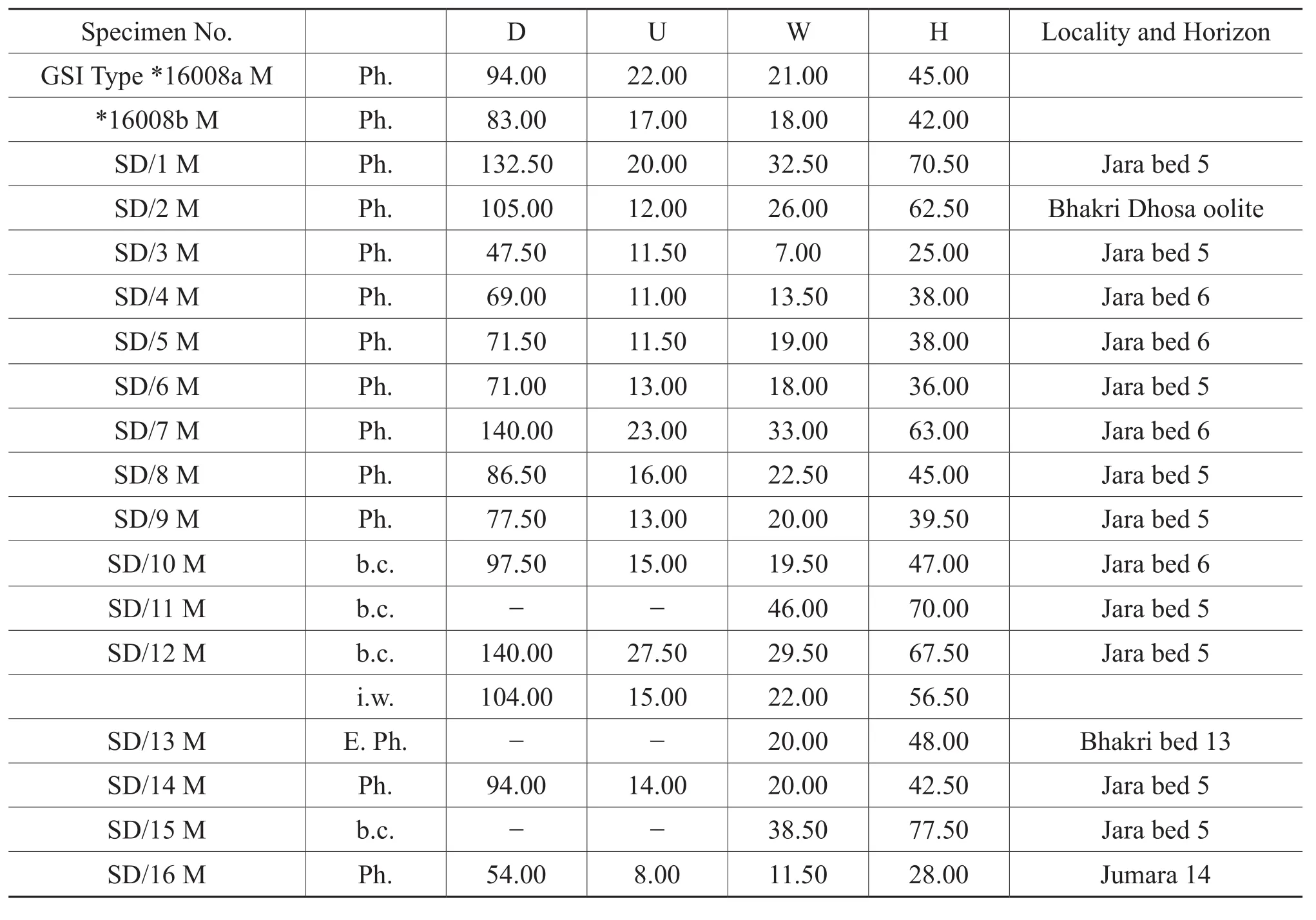
Table 6 Materials and dimensions of Sublunuloceras discoides Spath
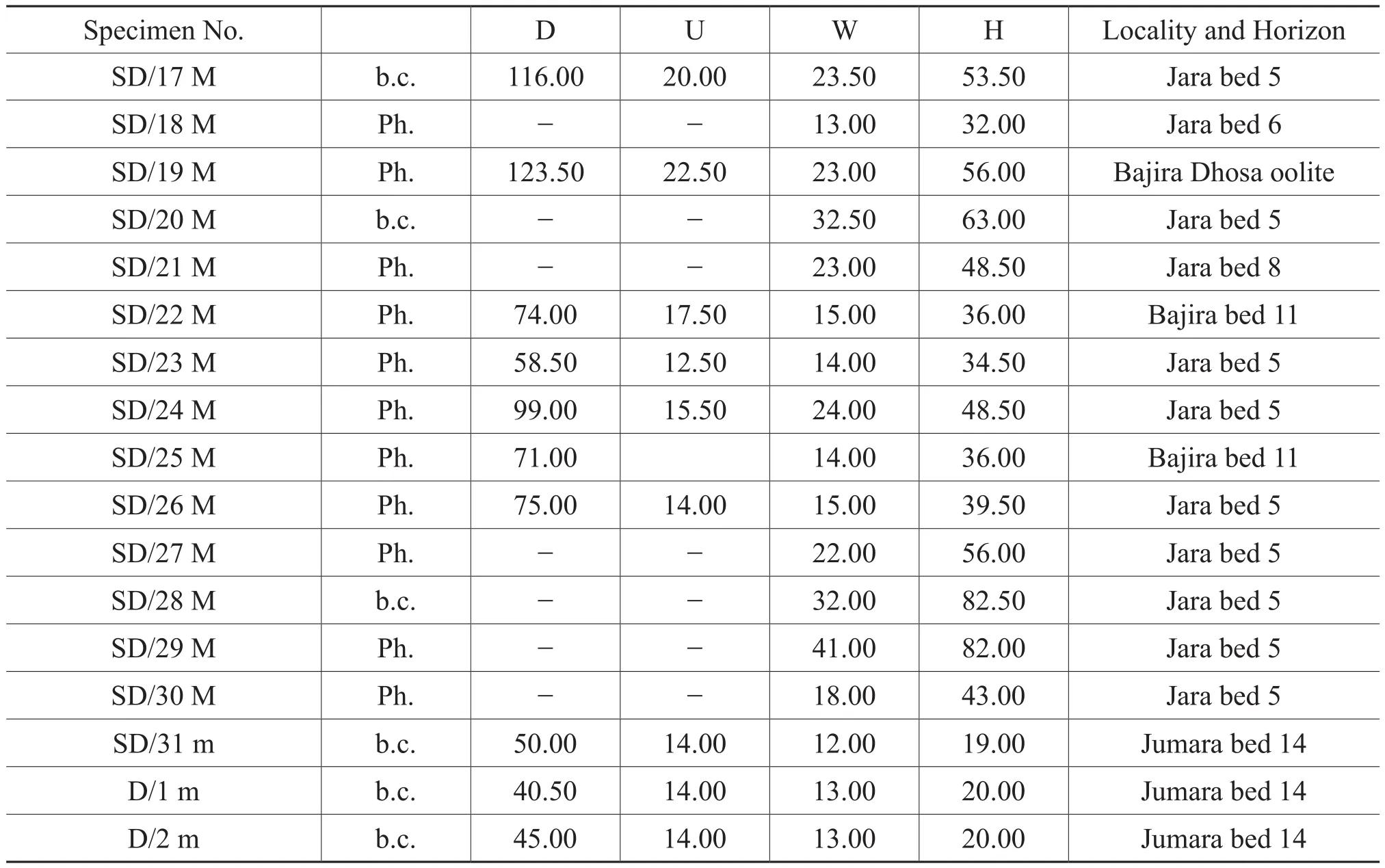
Table 6, continued
1958Sublunuloceras discoides, Spath, Collignon,pl.32, figure 150.
1980.Sublunuloceras discoides, Spath, Kanjilal,p.34, figure 1a, 1b.
Description [M]: Shell discoidal, strongly involute(U/D = 0.15 - 0.18); maximum end phragmocone diameter varies from 113 mm to 145 mm.Shell is strongly compressed (W/H = 0.28 to 0.66)with elliptical whorl section.
Incipient falcoid ribbing first appears after an initial smooth stage of about 20-25 mm diameter.Primaries emerging from the umbilical margin go prorsiradiately towards the midflank and bifurcate.Secondaries thus originate and go rursiradiately towards the ventro-lateral margin.Primaries become blunt or disappear from the inner flank at a diameter of 40-50 mm.Secondaries are relatively strong, sharp and continue up to the ventro-lateral margin at this stage of growth, but they also become incipient in the latter phragmocone.Primaries completely disappear from the inner flank at the latter phragmocone (60-80 mm diameter), where only flattened, distant secondaries continue to the outer flank.At the adult phragmocone, the shell becomes almost smooth with distantly spaced, flattened, incipient secondaries.Two fragmentary specimens with incompletely preserved body chamber fragments show almost no ornamentation at the final whorl.
Venter is narrow with a sharp mid-ventral keel at the early whorls as well as in latter phragmocones.In the adult phragmocone and also in the body chamber the venter becomes broad and the keel becomes faint.
Umbilicus is narrow, deep in early phragmocone stage with an inclined wall and rounded margin.The umbilical wall becomes steep, but the margin remains rounded in late phragmocone as well as in the body chamber.
Description [m]: Shell is relatively evolute (U/D =0.28-0.31), extremely compressed (W/H = 0.63 - 0.68)with maximum adult size varying from 40 mm to 50 mm.
Initial whorl smooth up to 9 mm diameter then strong rectiradiate primaries appear near the umbilical margin,which go up to mid flank and bifurcate.Secondaries continue up to the ventro-lateral margin where they terminate forming weak bullae.Primary ribbing gradually becomes prorsiradiate and stronger in inner whorls and bifurcates below the mid flank.Secondaries continue rursiradiately up to the ventro-lateral margin and terminate there by forming elongated bullae.In the latter phragmocone (D= 25 mm)primaries become stouter, more prorsiradiating and bifurcate at the mid flank.Closely spaced secondaries are rursiradiate and concave towards the aperture.They terminate at the ventro-lateral margin forming clavi or elongated bullae.Ornamentation continues up to the endof adult body chamber; although in one variant ribbing becomes indistinct in the internal mould at the last part of the body whorl.
Venter narrow and keel is not prominent in inner whorls.In the adult phragmocone the venter becomes wider with a prominent mid ventral keel; the sharpness of the keel diminishes at the body whorl and the venter becomes smooth and broad at the last part.
The umbilicus is wide from the beginning and becomes wider with ontogeny.Both umbilical margin and slope are gradual.
Whorl section is elliptical in inner whorls as well as in the phragmocone and becomes subtrapezoidal in the body chamber.
Apertural modification presents; spatulate projection of apertural portion suggests presence of lappet.
Discussion: The present species has two sexual antidimorphs.Typically smooth outer whorls and discoidal whorl shape are the two distinguishing morphological features that separate the present species from other related forms.Inner whorls of some variants ofS.discoideshave a similar wider umbilicus as inS.aff.nodosulcatum(Lahusen)(Spath, 1927-1933, pl.15, figure 1a, 1b), but it has coarser as well as more angulirursiradiate ribbing and an acute periphery.
S.dynastes(Waagen)shows a smaller umbilicus as in macroconchiateS.discoides; but in the later stage of growth there appear many distinguishing features such as strongly ribbed outer whorls with bluntly fastigate periphery; moreover,S.dynastesis confined to Upper Callovian,whereas,S.discoidescontinues into the Middle Kimmeridgian (Kanjilal, 1980).
S.lairense(Waagen)of Spath (1927-1933)is more evolute and coarsely ornamented in all stages of growth as compared to the present species.On the other hand,S.lairense(as stated by Spath, 1927-1933)shows typical weakening of primaries at the point of bifurcation, which is not shown by any otherSublunulocerasof Kutch.
Jeannet’s (1951)species from Lower Oxfordian of Herznach is similarly involute and has a sharp periphery and weakening of ribs in outer whorls like the present form, so it has been synonymized here.Kanjilal’s (1980)forms are relatively inflated although it is similar to the present form in weakening of ribs in later phragmocone.
vii.Sublunuloceras dynastes(Waagen)
Figure 13A-13M, Table 7
1875Harpoceras dynastesWaagen, pars, p.66,pl.13, figure 6 only.
1885Ludwiqia dynastes(Waagen); Haug, p.91.
1893 (?)Harpoceras dynastes, Waagen; Bonarelli,p.103.
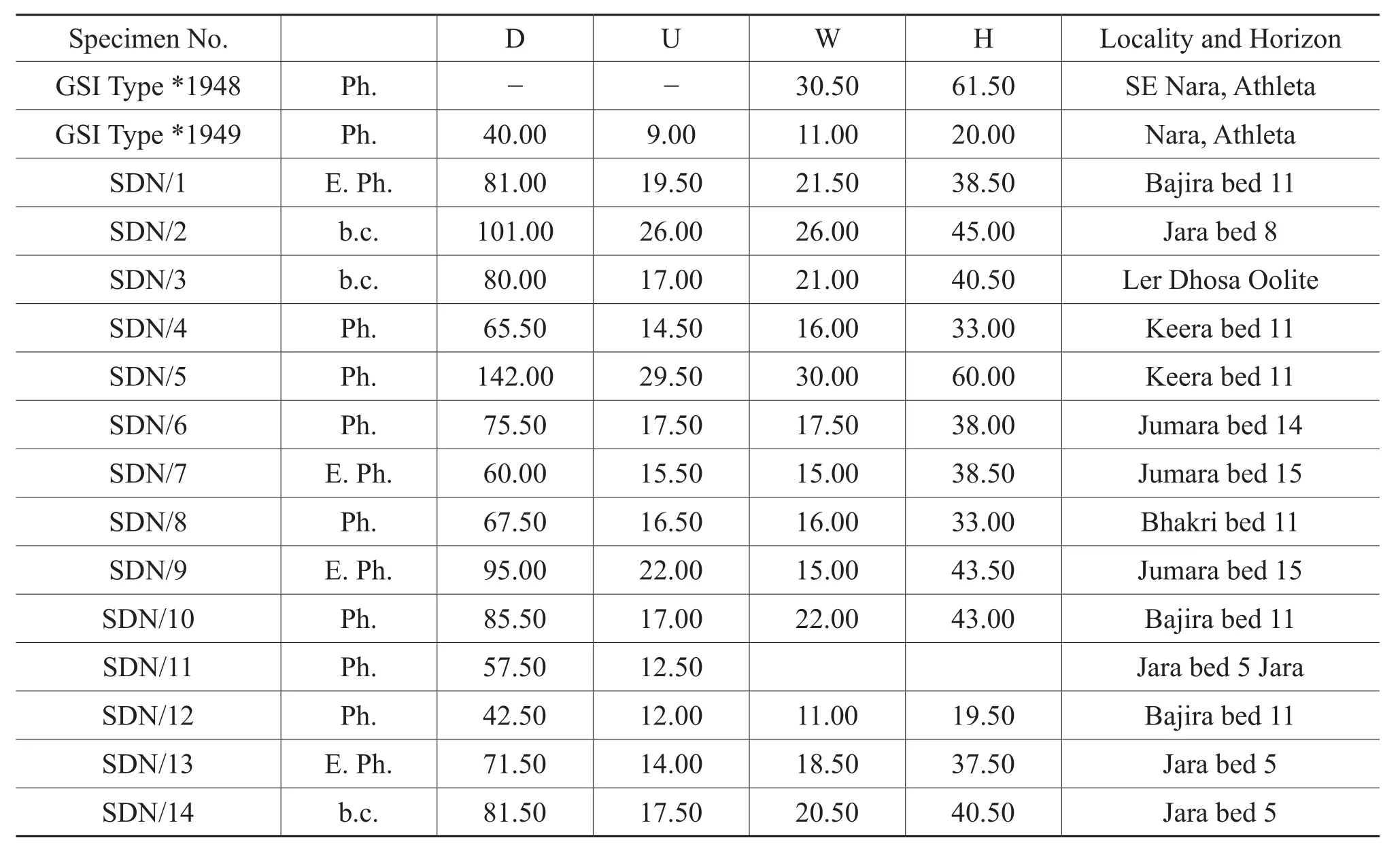
Table 7 Materials and dimensions of Sublunuloceras dynastes (Waagen)
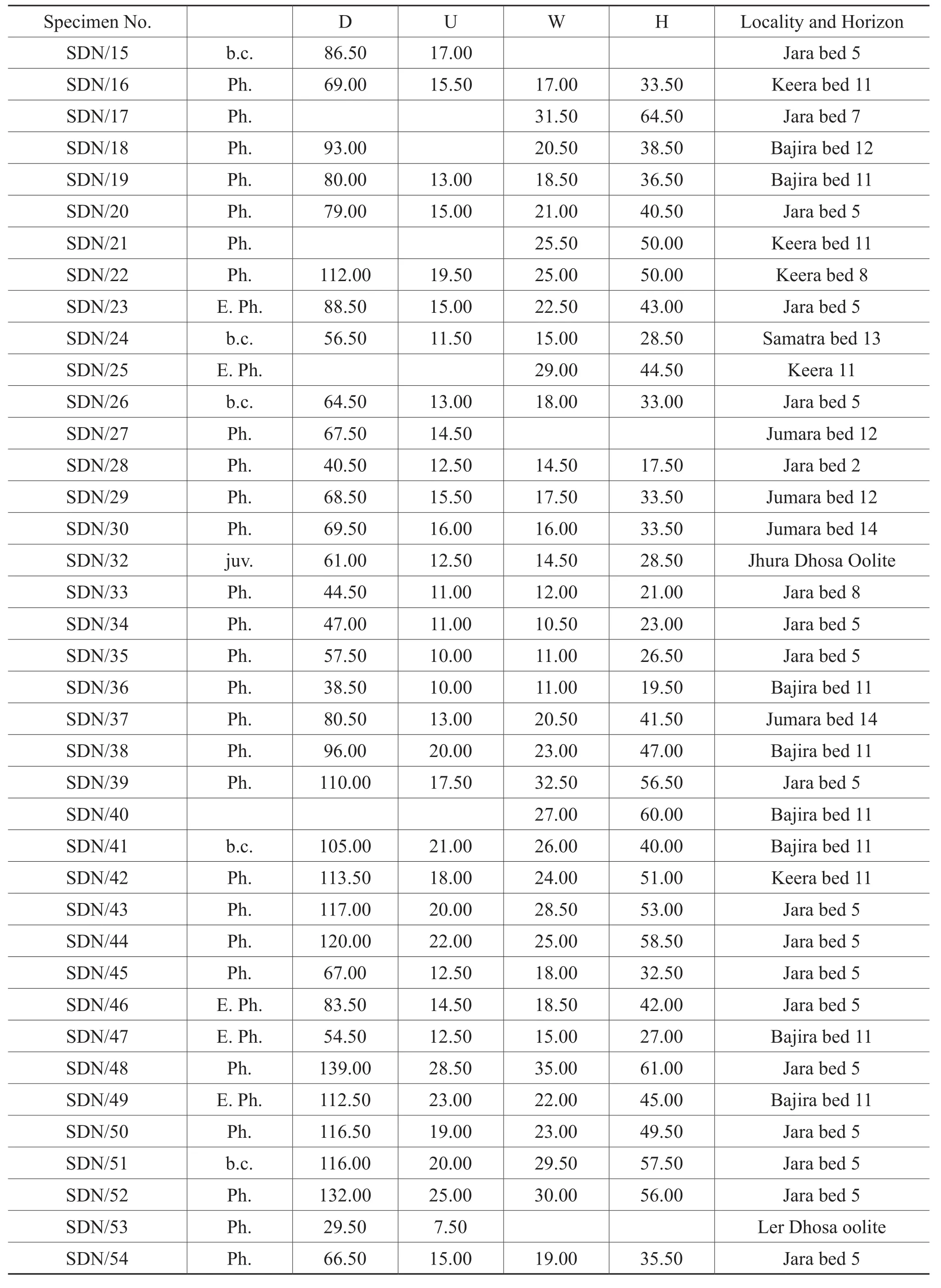
Table 7, continued

Table 7, continued
1912Harpoceras dynastes, Waagen; Smith (a),p.714; (b), pp.1350-1351.
1913Harpoceras dynastes, Waagen; Smith (c),p.422.
1915Harpoceras dynastes, Waagen; Smith, p.795 1928Sublunuloceras dynastes(Waagen), Spath,p.125, pl.11, figures 2a, 2b, 3.
1928Sublunulocerasaff.nodosulcatum(Lahusen),Spath, p.124, pl.15, figure 1a, 1b.
Description: Shell large maximum adult phragmocone diameter varies between 80 mm and 100 mm, and the preserved adult body chamber may reach up to 142 mm.Shell is discoidal evolute (U/D = 0.28 - 0.37)and compressed (W/H = 0.49 - 0.83)with an elliptical whorl section.Shell is smooth up to 10 mm diameter, and then closely spaced weak falcoid ribs appear near the umbilical margin.Primary ribs at this stage are straight to slightly prorsiradiate and bifurcate at the mid flank.Secondary ribs are sickle-shaped and rursiradiately continue up to the ventro-lateral margin; they terminate there by forming elongated bullae.There are about ten primary ribs and 19 secondary ribs per half whorl.At a still larger diameter(D = 38 mm)the shell becomes relatively involute (U/D= 0.26)and ribbing is stronger than in earlier whorls.Primaries are more prorsiradiate and secondaries remain rursiradiate in nature and form strong elongated bullae at the termination point near the ventro-lateral margin.The strength of ribbing increases with ontogeny, although primaries are obsolete or blunt in the inner lateral.In adult phragmocone ribbing becomes weakly falcoid to rectiradiate; primaries almost obsolete or disappear from the inner flank, although in some variants distant, weak primaries still continue; secondaries also become blunt and show weak clavate termination at the ventro-lateral margin.Only blunt secondaries continue in body chamber;they also are distant and form weak bullae at the ventrolateral margin.Body chamber starts with egressions of whorl.
Venter is rounded in early ontogeny with an incipient mid-ventral keel.Venter becomes fastigate with prominent keel in later phragmocone but in body chamber the venter becomes wide and keel becomes indistinct.
Umbilicus is wide at the early stage (U/D = 0.3)and becomes narrower in the middle stage of growth (U/D =0.26).In body chamber the umbilicus is again wider.Umbilical wall is always steep with rounded margin.
Discussion:Sublunulocerasaff.nodosulcatum(Lahusen)described by Spath (1927-1933, pl.15, figure 1a, 1b),has moderately wide umbilicus (U/D = 0.23)in its inner whorls and falcate to falcoid ribbing-those morphological features are close to similar sizedS.dynastes.InS.aff.nodosulcatumcostation appears at about 10 mm diameter as in the present species, but are more falcate in nature.Both species have more or less inflated outer whorls with similarly strong ribbing that continues up to the adult body whorl.S.dynastesdiffers fromS.nodosulcatumby its more fastigate venter and loss of keel at the body chamber.Although some of the variants of the present species show a keeled venter at least up to the preserved end of body chamber, so it may be a variable character for this present species.For those above mentioned reasons,S.aff.nodosulcatumof Kutch has been synonymized with the present species.
S.discoidesSpath is clearly distinguished from the present species by its discoidal shape, sharpened periphery and loss of costation at larger diameters.
The present species differs fromS.lairense(Waagen)by its non-tuberculate costation and bluntly fastigate periphery.
5)GenusLunulocerasBonarelli, 1893
Type speciesNautilus lunulaReinecke, 1818
i.Lunuloceras lunula(Zieten)
Figure 14A-14H, Table 8
1840Ammonites corrugates; J.de.C.Sowerby; in Grant, pl.23, figure 12.
1850Ammonites orientalis,d’Orbigny, p.331.
1875Harpoceras lunula(Zieten), Waagen, p.63,pl.12, figure 1a, 1b.
1891Ludwigiasp.ind.I, v.d.Borne, p.9.
1893Hecticoceras(Lunuloceras)lunula(Reinecke)Bonarelli, pars.p.99.
1902Ammonites corrugates; J.de.C.Sowerby;Blake, p.35.
1928Lunuloceras orientale(d’Orbigny), Spath,p.118, pl.13, figure 9.
1928Lunuloceras nisoides, Spath, pl.13, figure 8b;pl.14, figure 11, pl.15, figure 5.
1912 (?)Harpoceras lunula(non Reinecke), Smith,p.1352
1956H.(L.)lunula(Reinecke), Zeiss, p.33, pl.1,figure 5.
Description: Shell is medium sized (maximum diameter of the shell is 62 mm), planulate, evolute (U/D = 0.33)and compressed (W/H = 0.64), with adult phragmocone diameter from 40 mm to 57 mm.
The shell is smooth up to 3-4 mm diameter then faint nearly rectiradiate primaries appear near the umbilical margin.Primaries at this stage are mostly sharp and numerous but indistinct.However, in some variants the inner flank remains smooth up to a considerable diameter; only faint primaries appear at 25 mm diameter.Secondaries at this stage are rursiradiate and falcoid in nature.At later phragmocone primaries become a little stronger than the earlier whorls; they disappear at the adult phragmocone and the inner flank remains smooth on the body whorl.Secondaries continue with the same strength up to the adult phragmocone and they remain concave towards the aperture.They form elongated bullae at the ventro-lateral margin and terminate.However, at later part of the endphragmocone and on the body chamber the secondaries are gradually restricted only at the upper flank.
Venter narrow and weakly fastigate in early whorls with trapezoidal whorl section (W/H = 0.52).It becomes gradually wider with a more fastigate nature during ontogeny.The venter has a hollow floored keel at early stage and continues up to the phragmocone but loses strength at the end-phragmocone and on the body whorl.
Umbilicus is fairly wide (U/D = 0.31)and has steep umbilical slope and gradual margin at early whorls.The margin becomes sharper during ontogeny.
Discussion: Spath (1927-1933)redescribed Waagen’s (1875)Harpoceras lunulaasLunuloceras orientale(d’Orbigny)and he showed the reason thatL.lunulas.st.(Reinecke)are rare in occurrence and Waagen (1875)had described the species based on a single specimen.Moreover, he also stated that otherLunulocerasspecies which were earlier identified as Reinecke’s trueL.lunula,most of them were later found to be different when revisited.When the author compares the present species with Spath’s (1927-1933)L.orientaleas well as Waagen’s(1875)H.lunulawith additional systematic collections,he has found that all of the specimens have certain common features like highly compressed whorls, smooth inner whorls and indistinct keel.Though ribbing pattern varies to some extent, most differences may be accounted for by intra-specific variation.Therefore, the author prefers to synonymize both Waagen’s (1875)species and Spath’s(1927-1933)species asL.lunula.
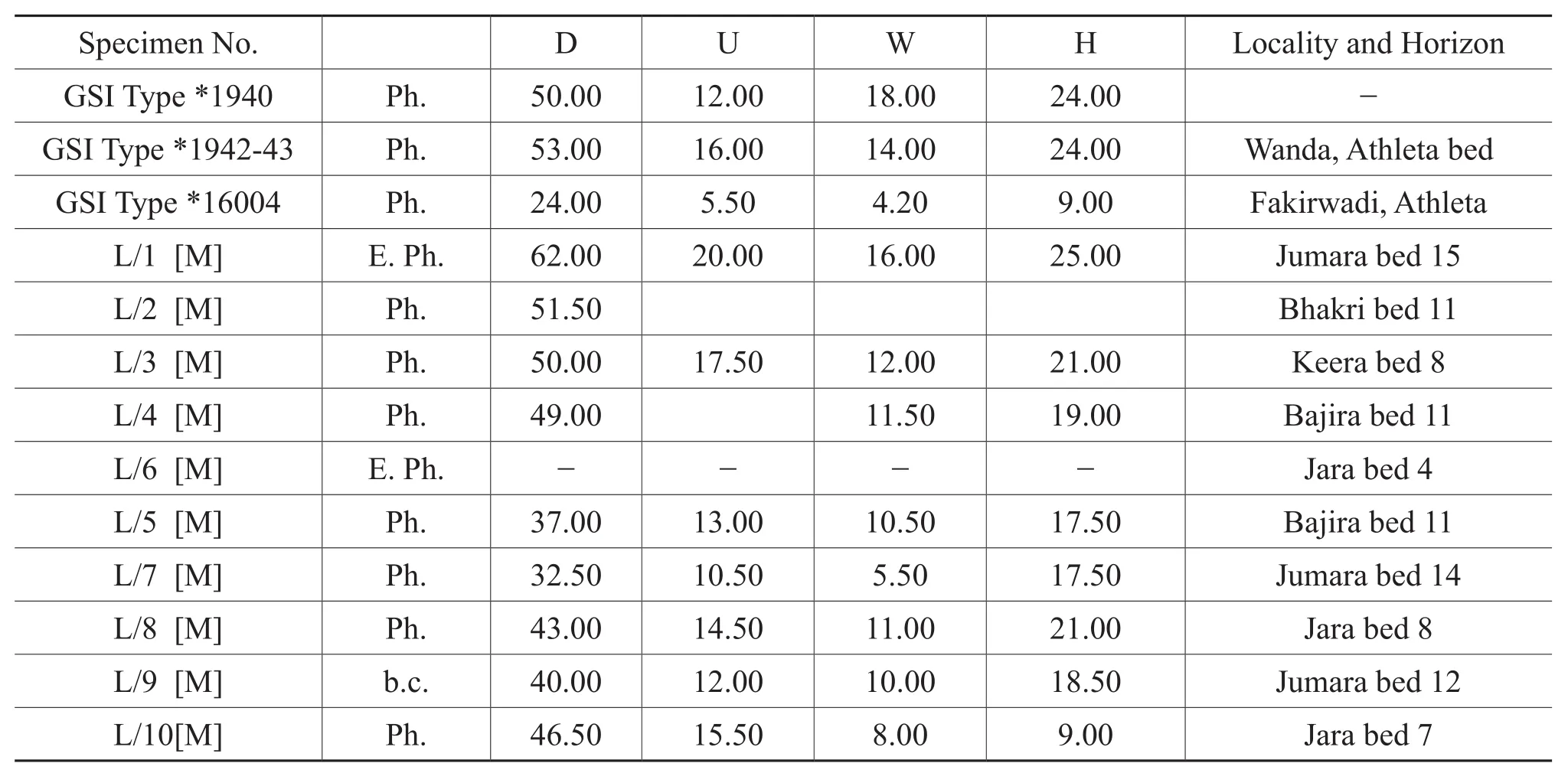
Table 8 Materials and dimensions of Lunuloceras lunula (Zieten)
Spath’s (1927-1933)L.nisoidesfrom the Athleta beds of an unlocalized section has similarly smooth inner whorls, and falcoid to angulirursiradiate ribbing toL.lunulawith half of the last whorl belonging to body chamber.By comparing the morphological features and morphometrics, the author prefers to synonymize it within the present species.
Spath’s (1927-1933)L.orientalevar.plana(here included withinL.lunula)has similar non-tuberculate ribs,compressed whorl and narrowly fastigate venter likeSublunuloceras lairense(Waagen); but unlike the present species it has bifurcating ribs.
The present species, although a bit larger, has identical ornamentation like Zeiss’s (1956)species from Blumberg,Germany.
6)GenusBrightiaRollier, 1922
Type speciesHecticoceras nodosumBonarelli, 1893
ii.Brightia callomonin.sp.
Figure 15A-15J, Table 9
1928Brightiasp.ind.Spath, p.121, pl.18, figure 7.
Etymology: The species is named after the great palaeontologist Dr.J.H.Callomon.
Description [M]: Shell is medium sized (maximum preserved diameter 74 mm), fairly evolute (U/D = 0.31 to 0.47)and compressed (W/H = 0.5 to 0.69)throughout ontogeny.
Shell is smooth up to about 6 mm diameter; then rectiradiate to slightly prorsiradiate primaries appear near the umbilical wall.Weak dense secondaries also appear simultaneously.Strength of ribbing increases with growth,although primaries are still stronger than secondaries.At the later phragmocone stage primaries become slightly tuberculate and more prorsiradiating and relatively distant than earlier whorls.At this stage of growth (D = 30 mm)the rursiradiate secondaries also become stronger and there are 2-3 secondaries that originate from tuberculate primaries at one-third whorl height.The largest fragmentary specimen is septate (D = 74 mm)and shows distant,robust, prorsiradiate primaries which are bifurcating at one-third whorl height and rursiradiate secondaries continue to ventro-lateral margin.The secondaries form weak bullate termination at ventro-lateral margin.This nature of secondaries continues all throughout ontogeny.There are about 10-11 primaries and 20 to 25 sencondaries per half whorl at this stage of growth.
Umbilicus is widely open throughout ontogeny with no distinct umbilical wall or margin in the early stages, although, a distinct inclined umbilical wall with a rounded margin is present in the adult phragmocone.
Venter is rounded to narrowly fastigate in early stage of growth with a weak non-floored keel at the mid venter.On the later phragmocone, the venter becomes weakly fastigate and the keel is also prominent.At the body chamber, the venter is still widely fastigate but the keel becomes faint.
Description [m]: Shell is small (maximum preserved diameter is 31.5 mm), strongly evolute (U/D = 0.52), compressed (W/H = 0.78), with last half whorl occupied by the body chamber (adult phragmocone diameter = 28 mm).
Prominent primary ribs appear as early as 5 mm diameter.After they appear, they soon become tuberculate just near the umbilical margin.Strongly tuberculate primaries are projected forward and furcated into 2-3 weak secondaries which are rursiradiate in nature.At the later phragmocone those primaries become strongly elongatedbullae which are also prorsiradiating and furcated into 2-3 strongly rursiradiate secondaries, thus giving rise to an angulirursiradiate pattern.The whorl shows weak uncoiling after the adult phragmocone but distant bullate primaries still continue up to the end of the whorl preserved.
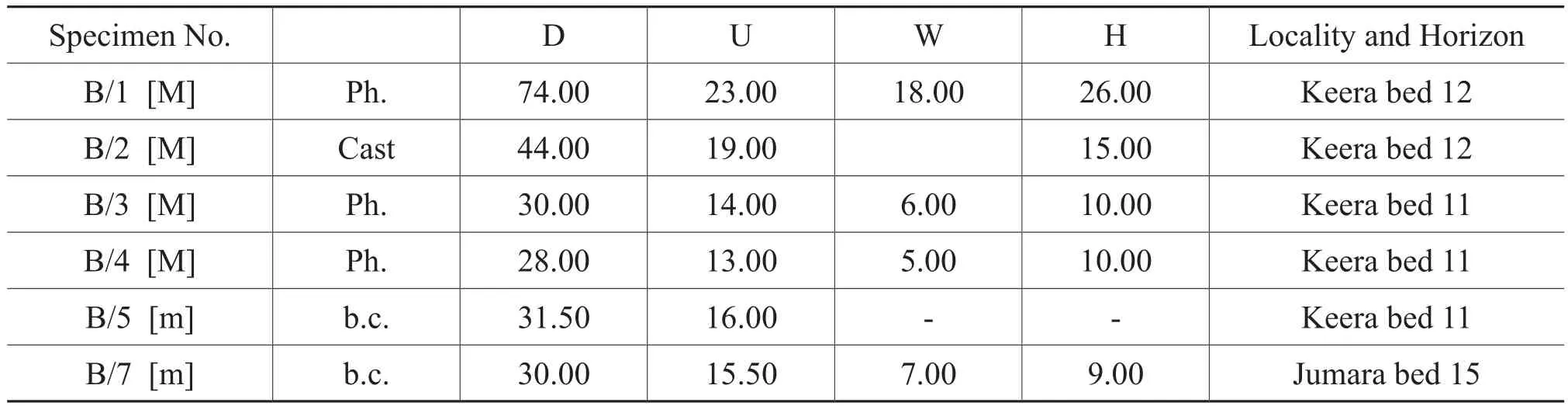
Table 9 Materials and dimensions of Brightia callomoni n.sp.
The umbilicus is widely (U/D = 0.5)open with rounded margin and gentle umbilical slope.The slope becomes a little steeper at later whorls.
Figure 3 Kheraites ignobilis (Waagen)[M].A-C-Lateral(A), apertural (B)and ventral (C)views of phragmocone specimen K/17 from bed 2 of Jara; D-E-Lateral (D)and ventral (E)views of phragmocone specimen K/16 from bed 5 of Keera, note strong falcoid ribbing and smoothing of the inner flank at later phragmocone; F-H-Lateral (F), apertural (G)and ventral (H)views of endphragmocone specimen K/11 from bed 4 of Jara,note keeled venter; I-K-Lateral (I), apertural (J)and ventral (K)views of phragmocone specimen K/19 from bed 9 of Jumara; LLateral view of endphragmocone specimen K/21 from bed 7 of Keera, note smooth inner flank.White bar adjacent to each figure measures 2 cm.
Venter is narrowly fastigate with a mid-ventral keel at the early stage.However, it gradually becomes widely fastigate during ontogeny, although the keel becomes faint at the adult body chamber.
Discussion: Spath (1927-1933)describedBrightiasp.ind.with a fragmentary 35 mm-long whorl and he compared this form with the Persian formB.metomphalaidentified by Parona and Bonarelli (1897).While revisiting the Kutch form along with the additional collections the author has found that the EuropeanB.metomphaladescribedby Jeannet (1951)is more bluntly ribbed and has more inflated whorl section at a larger diameter.The ribbing pattern of EuropeanB.salvadoridescribed by Jeannet (1951)is close to the present species but it is relatively involute and is more densely costate than the Kutch species.Because of these above reasons, the Kutch specimens have been described as a new species.
Figure 4 Kheraites ignobilis (Waagen)[m].A-B-Lateral (A),apertural (B)views of a nearly complete specimen K/7 from bed 9 of Jumara, note strong falcoid ribbing; C-E-Lateral (C), ventral(D)and apertural (E)views of specimen K/5 with preserved body whorl from bed 7 of Keera, note strong falcoid ribbing continues at the adult body chamber; F-Lateral view of specimen K/1, with last half whorl body chamber, from bed 10 of Jumara, note strong rectiradiate ribbing forming bullae at the ventro-lateral margin;G-I-Lateral (G), ventral (H)and apertural (I)views of specimen K/9 with partly preserved body whorl from bed 10 of Jumara;J-Lateral view of endphragmocone specimen K/3 from bed 8 of Jumara, note smooth inner flank; K-Lateral view of a body chamber specimen (K/2)from bed 7 of Keera, note strong falcoid ribbing.White bar adjacent to each figure measures 2 cm, x means the beginning of body chamber.

Figure 5 Putealiceras trilineatum (Waagen)[M].A-C-Lateral (A), ventral (B)and apertural (C)views of a body whorl fragment (specimen P/6)from bed 8 of Jara, note falcoid ribbing forming clavi at the ventro-lateral margin; D-Lateral view of specimen P/14 from bed 13 of Samatra; E-Cast of venter of the preceding whorl of specimen P/6, note tricarinate venter; FH-Lateral (F), apertural (G)and ventral (H)views of a body chamber fragment (specimen P/8)from bed 13 of Jumara, note tricarinate venter (H); I-K-Lateral (F), apertural (G)and ventral(H)views of GSI Type specimen 1944 with body chamber preserved from bed 11 of Keera.White bar = 2 cm.
Hecticoceras(Brightia)tenuinodosumdescribed by Zeiss (1956)from Jason Zone of Blumberg has a smooth inner flank or a few incipient tubercles present irregularly;moreover, it has smooth inner whorls unlike the present species.

Figure 6 Putealiceras trilineatum (Waagen)[M].A-C-Lateral (A), apertural (B)and ventral (C)views of a nearly complete specimen P/5 from bed 7 of Jara, note rectiradiate ribbing and tricarinate venter; D-E-Lateral (D)and ventral (E)views of body whorl fragment (specimen P/11), note falcoid ribbing and tricarinate venter; F-G-Lateral (F)and ventral (G)views of incomplete specimen P/13 from bed 5 of Jara.Putealiceras intermedium Spath [M].H-J-Lateral (H), apertural (I)and ventral (J)views of incomplete GSI Type specimen 15995 (holotype)from Upper Callovian (Athleta bed of Spath); K-Lateral view of a juvenile specimen P/31 from bed 5 of Jumara.L-N-Lateral (I), apertural(J)and ventral (K)views of GSI Type specimen 15997 with septate whorl from Upper Callovian (Athleta bed of Spath)of Samatra.White bar = 2 cm.x means the beginning of body chamber.
H.(B.)aff.diforme(De Tsytovitch)from the Athleta beds of Blumberg reported by Zeiss (1956)has outer whorls almost identical with the present form, but the inner whorls are completely smooth unlike the Kutch species.
3.3.1Biostratigraphic implications of Hecticoceratin genera
Hecticoceras giganteumspans the semilaevis horizonof the Formosus Zone to perisphinctoides horizon of the Anceps Zone (Table 11).In semilaevis horizonH.giganteumco-occurs withNothocephalitessemilaevis(Waagen)the zonal species, along withMacrocephalitesformosus(J.De.C.Sowerby),Kheraiceras cosmopolitum(Parona and Bonerlli),Kamptokephaliteslamellosus(J.De.C.Sowerby),Dolikephalites subcompressus(Waagen),ReineckeiatyranniformisSpath,Reineckeiaanceps(Reinecke),Collotia oxyptycha(Neumayr)and others (Spath, 1927-1933; Cariou and Krishna, 1988; Datta, 1992; Bardhanet al., 2002; Janaet al., 2005).In theAncepsSubzone of the basal Middle CallovianH.giganteumis associated withEucycloceras opis((J.De.C.Sowerby),Reineckeia tyranniformis,R.ancepsandChoffatiaspp.This level marks the disappearance ofMacrocephalitesspp.(Table 11).WhileKheraicerasspp.H.giganteumgoes up to the perisphinctoides horizon of the Opis Subzone and is found to be associated withIdyocycloceras perisphinctoidesSpath,the zonal index,E.opis,PhlycticerasandReineckeiaspp.continue (Janaet al., 2005).

Figure 7 Putealiceras intermedium (Spath)[M].A-C-Lateral (A), apertural (B)and ventral (C)views of incomplete GSI Type specimen 15996 from the Upper Callovian (Divesian, Athleta bed of Spath), note rectiradiate ribbing becomes falcoid in the later phragmocone; D-E-Lateral (D)and ventral (E)views of complete specimen P/1 from bed 11 of Keera, note falcoid ribbing and smoothing of inner flank in the body chamber; FG-Lateral (F)and ventral (G)views of complete specimen P/2 from bed 8 of Keera; H-I-Lateral (H)and ventral (I)views of endphragmocone specimen P/3 from bed 8 of Keera; J-K-Lateral (J)and ventral (K)views of incomplete specimen P/25 from bed 5 of Jara.White bar = 2 cm, x means the beginning of body chamber.

Figure 8 Putealiceras vijaya Spath [M].A-B-Lateral (A)and ventral (B)views of completely septate specimen P/26 from bed 5 of Jara; C-Lateral view of young specimen P/23 from bed 11 of Bajira; D-F-Apertural (D), lateral (E)and ventral (F)views of complete specimen P/18 from bed 12 of Jumra, note strong, falcate ribs forming elongated bullae at the mid flank;G-I-Lateral (G), ventral (H)and apertural (I)views of a body chamber fragment P/24 from bed 5 of Jara; J-L-Lateral (J),apertural (K)and ventral (L)views of complete specimen P/20 from bed 5 of Jara.White bar = 2 cm, x means the beginning of body chamber.
Janaet al.(2005)have attempted a correlation of the Kutch biozonation with that of the SubmediterraneanProvince (Table 10).The top of theGracilisZone of France has been correlated with the Semilaevis Subzone of Kutch.They found faunal homogeneity within many horizons between these two regions.Macrocephalites gracilisshows strong resemblance toNothocephalites semilaevis(see also, Westermann and Callomon, 1988, p.16).The lower stratigraphic level ofH.giganteumtherefore corresponds toH.(J.)girodiBonarelli andH.boginensePetitclerc of boginense horizon of theGracilisZone of France.In fact the Kutch species is morphologically very similar to those European species (Roy and Bardhan, 2007).The highest occurrence ofH.giganteumis confined to the perisphinctoides horizon where one of the associated taxa isR.tyranniformisand Janaet al.(2005)matched this horizon with the blyensis horizon of France, which belongs to theTyranniformisSubzone.Recently Jain (1997)reportedPhlycticerasgr.pustulatum(Reinecke)at a similar level from Jumara in Kutch.It co-occurs withR.tyranniformis,Eucycloceras pilgrimiSpath(=E.opis; see Janaet al.,2005).Hecticocerass.st.is short ranging in France.It is confined only to the topmost Lower Callovian (boginense to kiliani horizons)(Cariou, 1984).However, in KutchHecticocerasappeared simultaneously in the top of the lower Callovian but lived longer and continued up to the lower part of theAncepsZone.In GermanyHecticocerass.st.also continued into a higher stratigraphic level and is found from the Middle to Upper Callovian (Schlegelmilch, 1985; Martill and Hudson, 1994).

Figure 9 Putealiceras dhosaense (Spath).A-C-Lateral (A),ventral (B)and apertural (C)views of incomplete specimen PD/4 from Dhosa oolite bed of Jumara; D-F-Lateral (D), ventral (E)and apertural (F)views of a fragmentary body chamber specimen PD/1 from Dhosa oolite bed of Ler; G-H-Lateral (G)and ventral (H)views of specimen P/6 with partly preserved body chamber fragment from Dhosa oolite bed of Bhakri.White bar = 2 cm.
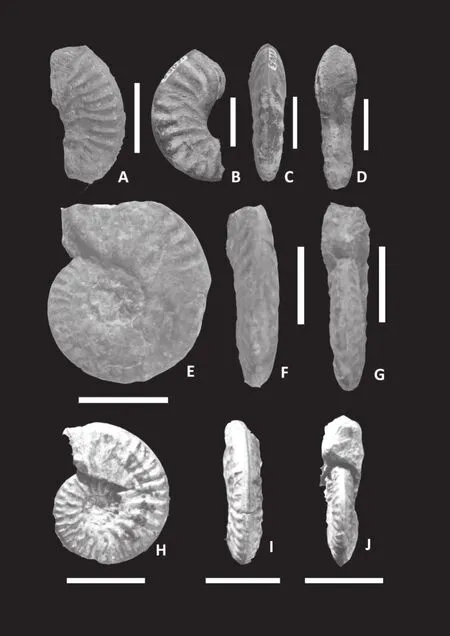
Figure 10 Sublunuloceras lairense (Waagen).A-Lateral view of GSI Type specimen 16006; B-D-Lateral (B), ventral(C)and apertural (D)views of a fragmentary body chamber of GSI Type specimen 16005 from Upper Callovian; E-G-Lateral(E), ventral (F)and apertural (G)views of incomplete GSI Type specimen 16007 from Fakirwadi, Athleta bed of Spath; H-JLateral (H), ventral (I)and apertural (J)views of incomplete GSI Type specimen 1945 from Ler, Athleta bed of Spath.White bar= 2 cm.
The EuropeanAthletaZone is equivalent toPonderosumZone of Kutch;Peltoceras athletais quite similarin morphology toPeltoceras ponderosumas depicted by Datta (1992).The Trezeense horizon of theAthletaZone in Subtethyan Province is marked byHecticoceras(Orbinyiceras)trezeensewhich is associated withH.(Sublunuloceras)aff.dynastesandH.(Putealiceras)triliineatum.In Kutch,H.(Sublunuloceras)dynastesandH.(Put.)trilineatumthough appear in theSubgrossouvria aberranshorizon of theReissiZone, become prolific in the lower part of thePonderosumZone.Putealiceras intermediumandSublunuloceras dynastesoccur throughout thePonderosumZone in Kutch, although in the Subtethyan Province they co-occur again in theCollotia collotiformishorizon of theAthletaZone.Thus, the zonal assemblage reinforced the stratigraphic correlation between Kutch and Subtethyan Province, especially for the Upper CallovianAthletaZone.
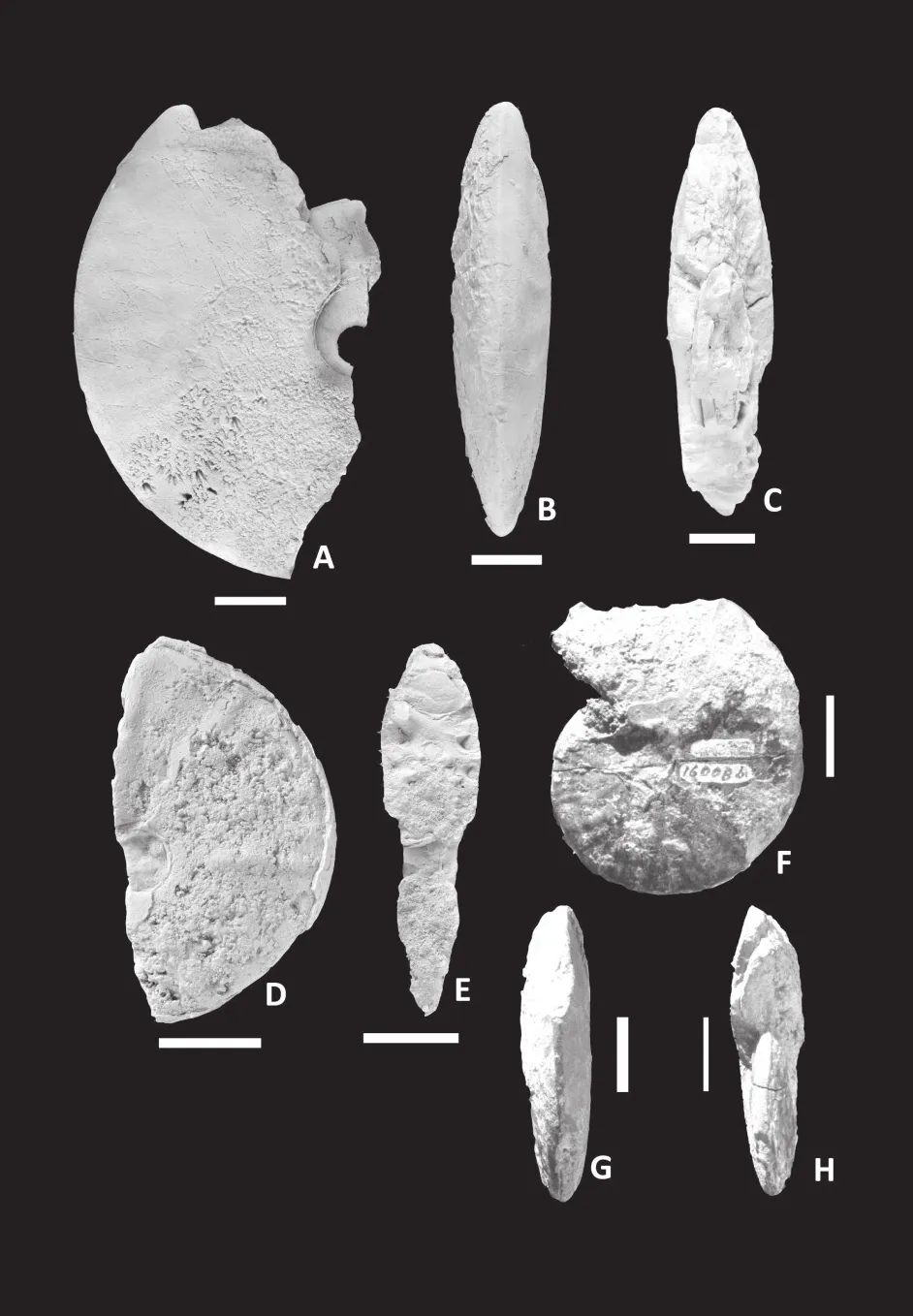
Figure 11 Sublunuloceras discoides Spath [M].A-C-Lateral (A), ventral (B)and apertural (C)views of partly preserved body chamber specimen SD/12 from bed 5 of Jara, note irregular blunt secondary ribs on outer flank; D-E-Lateral (D)and apertural (E)views of incomplete specimen SD/6 from bed 5 of Jara,note weakening of ribs in outer whorl and incipient blunt secondary ribs; F-H-Lateral (F), ventral (G)and apertural (H)views of GSI Type specimen 16008b (completely septate Holotype).White bar = 2 cm.
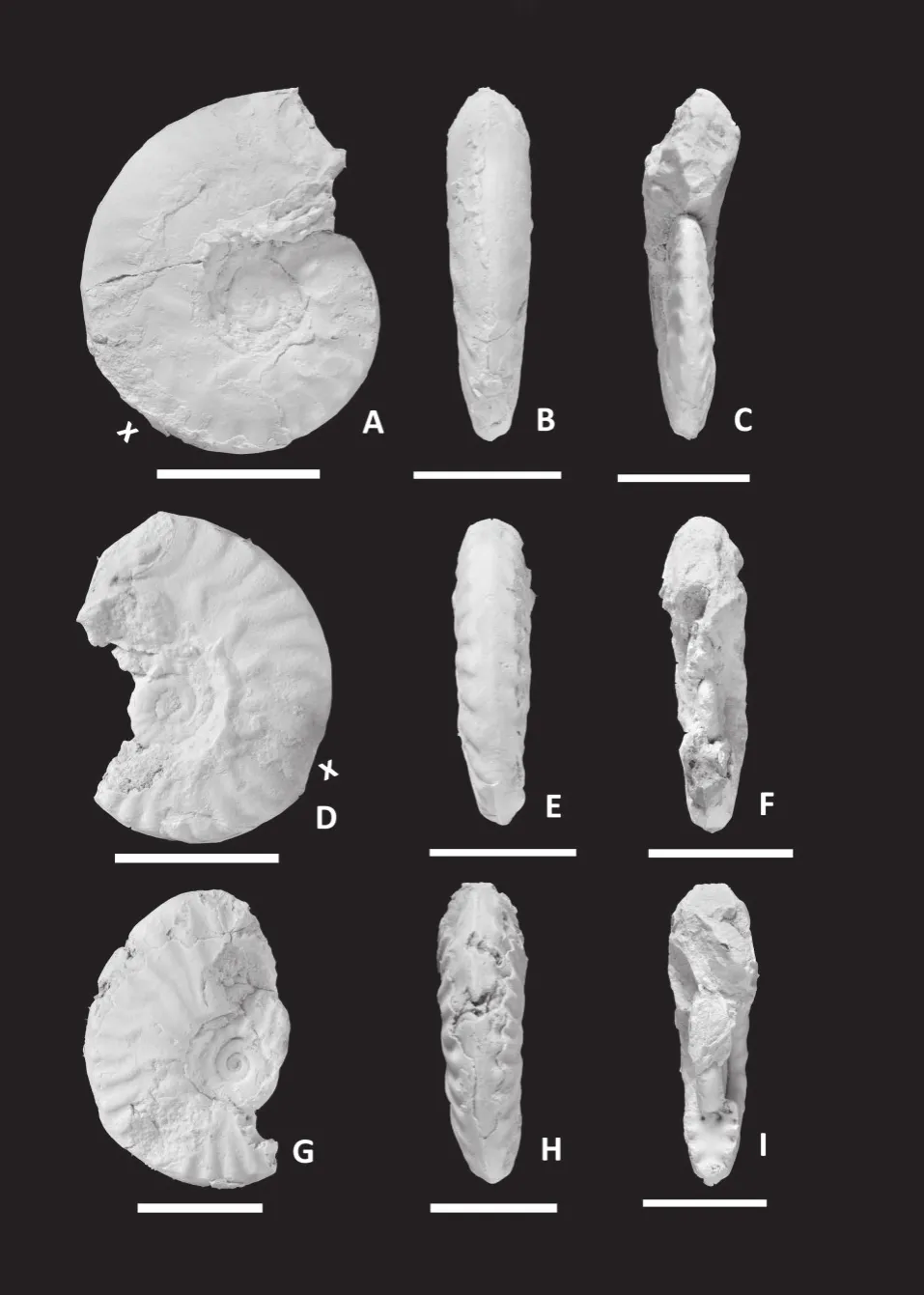
Figure 12 Sublunuloceras discoides Spath [m].A-C-lateral(A), ventral (B)and apertural (C)views of complete specimen SD/31 from bed 14 of Jumara, note presence of lappet and nearly smooth last part of body whorl; D-F-lateral (D), ventral (E)and apertural (F)views of complete specimen D/1 from bed 14 of Jumara, note presence of spatula-like lappet; G-I-lateral (G), ventral (H)and apertural (I)views of partly preserved body chamber specimen D/2 from bed 14 of Jumara.White bar = 2 cm, x means the beginning of body whorl.
3.3.2Palaeobiogeography of the family oppelidae during the Callovian-Oxfordian with special emphasis on Hecticoceratinae
During the Callovian oppelids are mainly represented by the hecticoceratins; there are only two genera:ProhecticocerasandHecticocerass.st.; both are cosmopolitan.Both are most diverse in the Mediterranean and Submediterranean compared to other provinces of the world including Kutch.In Submediterranean France the subfamily Hecticoceratinae is represented by three species ofHecticocerass.st.fourJeanneticerasspp., threeZietenicerasspp., nineChanasiaspp.and oneLunulocerasspp.(Cariou, 1984).Whereas in Kutch after the revision of Spath’s(1927-1933)forms I have encountered only two species of trueHecticocerasand oneKheraitesspp., three species ofPutealiceras, two species ofSublunuloceras, oneLunulocerasspp.and oneBrightiaspp.In Germany hecticoceratins are less diverse and are represented byH.hecticum(Reinecke)and twoJeanneticerasspp.(Schlegelmilch,1985).
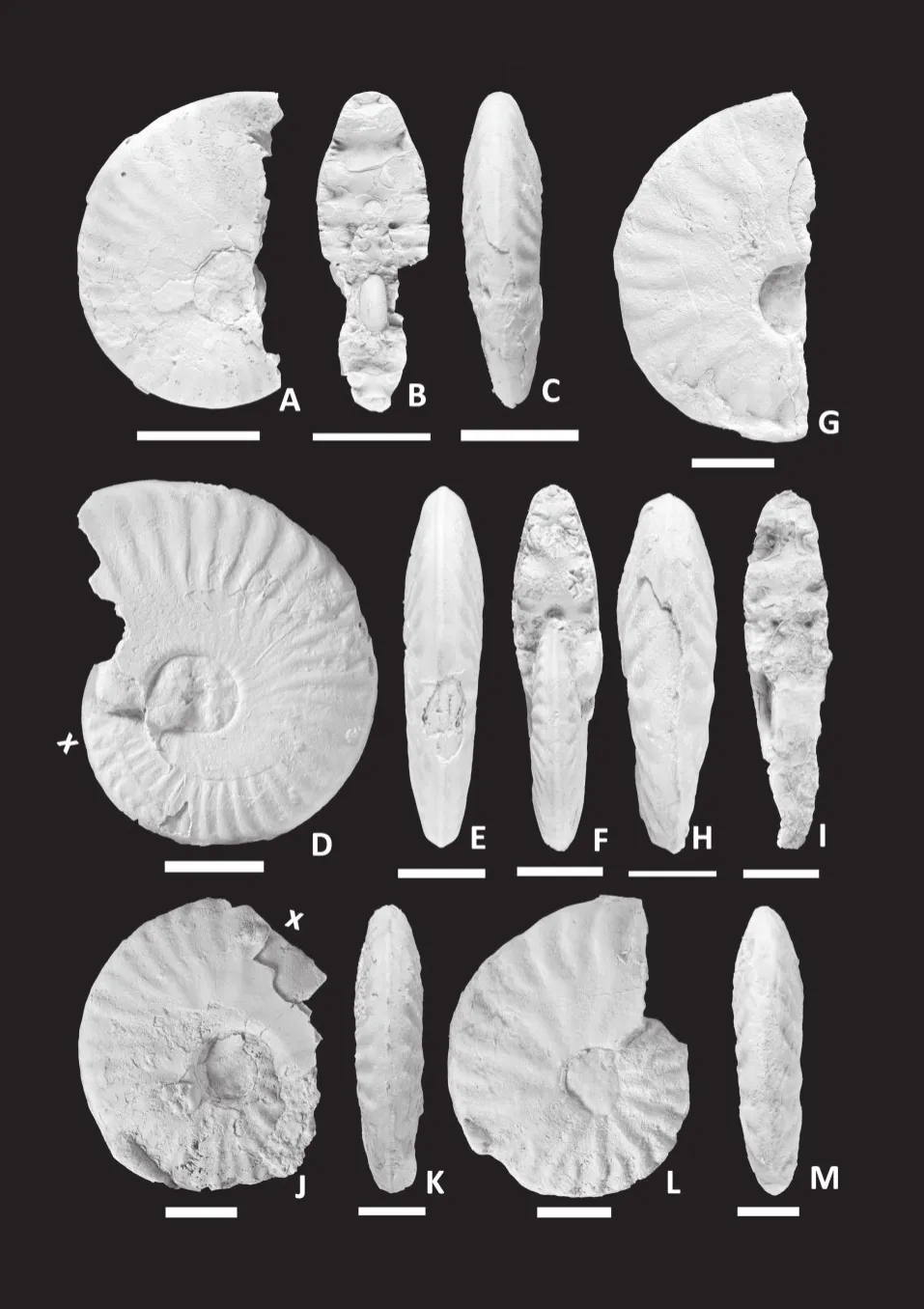
Figure 13 Sublunuloceras dynastes [Waagen].A-C-Lateral(A), ventral (B)and apertural (C)views of completely septate specimen SDN/61 from bed 11 of Bajira; D-F-Lateral (D), ventral (E)and apertural (F)views of incomplete specimen SDN/6 from bed 14 of Jumara, note trace of uncoiling suggests beginning of body chamber; G-I-Lateral (G), ventral (H)and apertural (I)views of completely septate specimen SDN/10 from bed 5 of Jara; J-K-Lateral (J)and ventral (K)views of partly preserved body chamber specimen SDN/1 from bed 11 of Bajira;L-M-Lateral (L)and ventral (K)views of specimen SDN/14 from bed 5 of Jara.White bar = 2 cm, x means the beginning of body whorl.

Figure 14 Lunuloceras lunula (Zieten).A-C-Lateral (A),ventral (B)and apertural (C)views of an adult phragmocone specimen L/1 from Dhosa oolite bed of Jumara; D-F-Lateral(D), ventral (E)and apertural (F)views of incomplete specimen L/10 from bed 7 of of Jara; G-Lateral view of a partly preserved body chamber specimen L/9 from bed 12 of Jumara; H-Lateral view of a nearly complete specimen L/3 from bed 12 of Jumara,note closely spaced secondary ribs on the outer flank.White bar= 2 cm, x means the beginning of body whorl.
Spath (1927-1933)described about seven species of“Alcidia” that need close scrutiny and might have been placed inOxyceritesand he also described oneParalcidiasp.from the Lower Callovian of Kutch.TheseOxyceritesgroup of genera forming a subclade remain unchanged and species-poor in the Lower Callovian but still cosmopolitan in distribution.In the Mediterranean and Submediterranean they are represented by fourOxyceritesspp.but rarely continued in the upper part of the Lower Callovian (Elmi,1967; Cariou, 1984).On the other hand, Parent (2006)has reportedOxyceritesoxynotusfrom the Middle Callovian in the East Pacific.
The hecticoceratin subclade shows a great diversification during the Middle Callovian, and Kutch is not an exception(Figure 16).In the Mediterranean and Submediterranean Province the subfamily Hecticoceratinae is represented by 82 species under 13 genera (Jeannet, 1951; Zeiss, 1956;Elmi, 1967; Cariou, 1984; Schlegelmilch, 1985)and Kutch shows homogeneity in generic distribution; the only exceptions areJeanneticerasandZieteniceraswhich are absent in the Indo-Madagascan Province.Spath (1927-1933)described 19 species under six genera from the Middle Callovian of Kutch.The present author, while revisiting Spath’s form in the light of sexual dimorphism and intraspecific variability distinguished only six genera and 11 biological species of which seven species have been dimorphically paired.In the Upper Callovian the subfamily Hecticoceratinae is consistent with the same generic diversity but the genera becomes species-poor.TrueHecticocerasare totally absent in all faunal provinces; at the same time a new genusi.e.Sublunulocerasappeared.In Kutch also the generic compositions of hecticoceratins remain the same as in the Mediterranean and Submediterranean andSublunulocerasis abundant along withPutealicerasandBrightia.Parent (2006)has recently describedH.(Sublunuloceras)cf.lairensealong withfour other hecticoceratins from the Upper Callovian of the East Pacific Province.The present work also included Spath’s (1927-1933)Sublunuloceras lairensebut the present species assemblage does not show homogeneity with South America and as such the provincial distinctiveness has been reflected in the composition of the hecticoceratin community.As in the Mediterranean-Submediterranean and East Pacific Province, the Indo-Madagascan Province also shows a sudden fall in diversity of hecticoceratins during the later part of the Upper Callovian, and onlySublunulocerasandBrightiacontinue into the Oxfordian.

Figure 15 Brightia callomoni n.sp.A-F and H-I = [M], G and J = [m].A-Lateral specimen B/2 from Dhosa oolite bed of Keera; B-C-Lateral (B)and apertural (C)views of incomplete specimen B/1 from Dhosa oolite bed of Keera; D-E-Lateral (D)and apertural (E)views of incomplete specimen B/3 from bed 11 of Keera; F-Lateral view of complete specimen B/4 from bed 11 of Keera; G-Lateral view of nearly complete specimen B/5 from bed 11 of Keera; H-I-Lateral (H)and ventral (E)views of incomplete specimen B/6 from bed 11 of Keera; J-Lateral view of complete specimen B/7 from Dhosa oolite bed of Jumara.White bar = 2 cm, x means the beginning of body whorl.
4 Palaeobiogeographic pattern analy‑sis with respect to Hecticoceratin distribution during the Callovian-Oxfordian
Phylogenetic patterns of hecticoceratin clades were used to deduce biogeographic patterns during the Callovian-Oxfordian.Biogeographic patterns in hecticoceratin phylogeny was evaluated and synthesized using a modified version of Brooks Parsimony Analysis (BPA)(Brooks, 1985, 1990; Wiley, 1988a, 1988b; Wileyet al.,1991).Several authors have described ways to apply cladistic approaches to biogeographic analysis, but the most prevalent approach is Brooks Parsimony Analysis.BPA is the method that currently offers the best prospects for application to the extant and fossil biota.In BPA the biogeographic patterns inherent in phylogenies are deduced by coding geographic distributions of ancestral nodes and terminal taxa as character states in a data matrix, where the“taxa” are the biogeographic regions of interest.This data matrix is then subjected to parsimony analysis.An ancestral biogeographic region is used as an outgroup, with all taxa treated as primitively absent.
In the present endeavour, BPA was applied in the two separate studies with biogeographic states from hecticoceratin phylogenies with an established cladograma (Figure 17; Roy, 2010)coded into two separate data matrices following Liberman and Eldredge (1996).First, BPA was applied in a vicariance analysis, in which only the biogeographic states of nodes; terminals that fit the vicariant pat-tern of progressive range contraction and diversification were coded into a data matrix.All characters related to transformations between ancestral and descendant nodes,and between ancestral nodes and descendant terminal taxa,involving expansion or change of range,i.e., some forms of dispersal, were omitted from this data matrix.
Second, a dispersal analysis was conducted with a data matrix constructed by considering those biogeographic states of nodes and terminals that fit a pattern of dispersal.For each analysis, data matrices were subjected to a parsimony analysis using an exhaustive search on PAUP(Swofford, 1990).Several steps are needed for coding the data matrices.First, the biogeographic states of terminal taxa are mapped onto the phylogenies as given in the cladogram (Figure 18).Then the nodes are coded by following Fitch optimization (Fitch, 1971), which doesn’t assume a priori the direction of biogeographic change and is best suited since the patterns of area relationships are not known beforehand.Data matrices were coded with presence in a biogeographic region treated as derived and denoted by a 1 or 2, and absence in a biogeographic region treated as primitive and denoted by a 0 rather than a ?(see Wiley, 1988a, 1988b for elaboration), following the assumption that in this case the fossil record reasonably reflects the evolutionary history of the hecticoceratin taxa being studied and absence represents true absence.This is a reasonable assumption considering the good state of preservation of these fossil faunas, as well as the fact that they occur in areas that have been intensely sampled by palaeontologists.
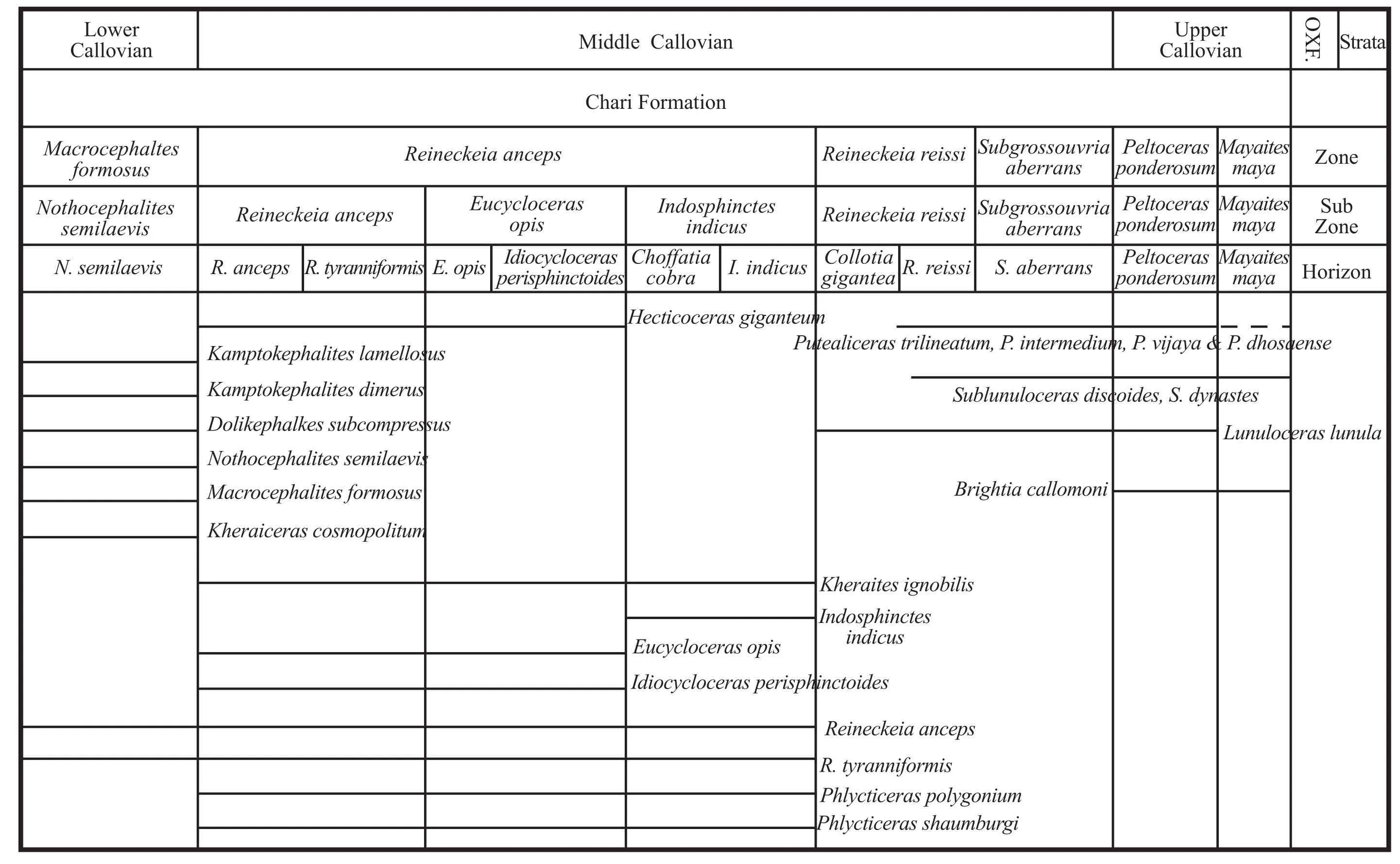
Table 11 Stratigraphic distribution of hecticoceratin species along with other important ammonite species is shown against the regional biostratigraphic zonation of Kutch.Widths of the biozones are not to scale (Source for other ammonites distribution: Jain, 1997; Bardhan et al., 2002; Jana et al., 2005, Kayal, 2009 and the present study).
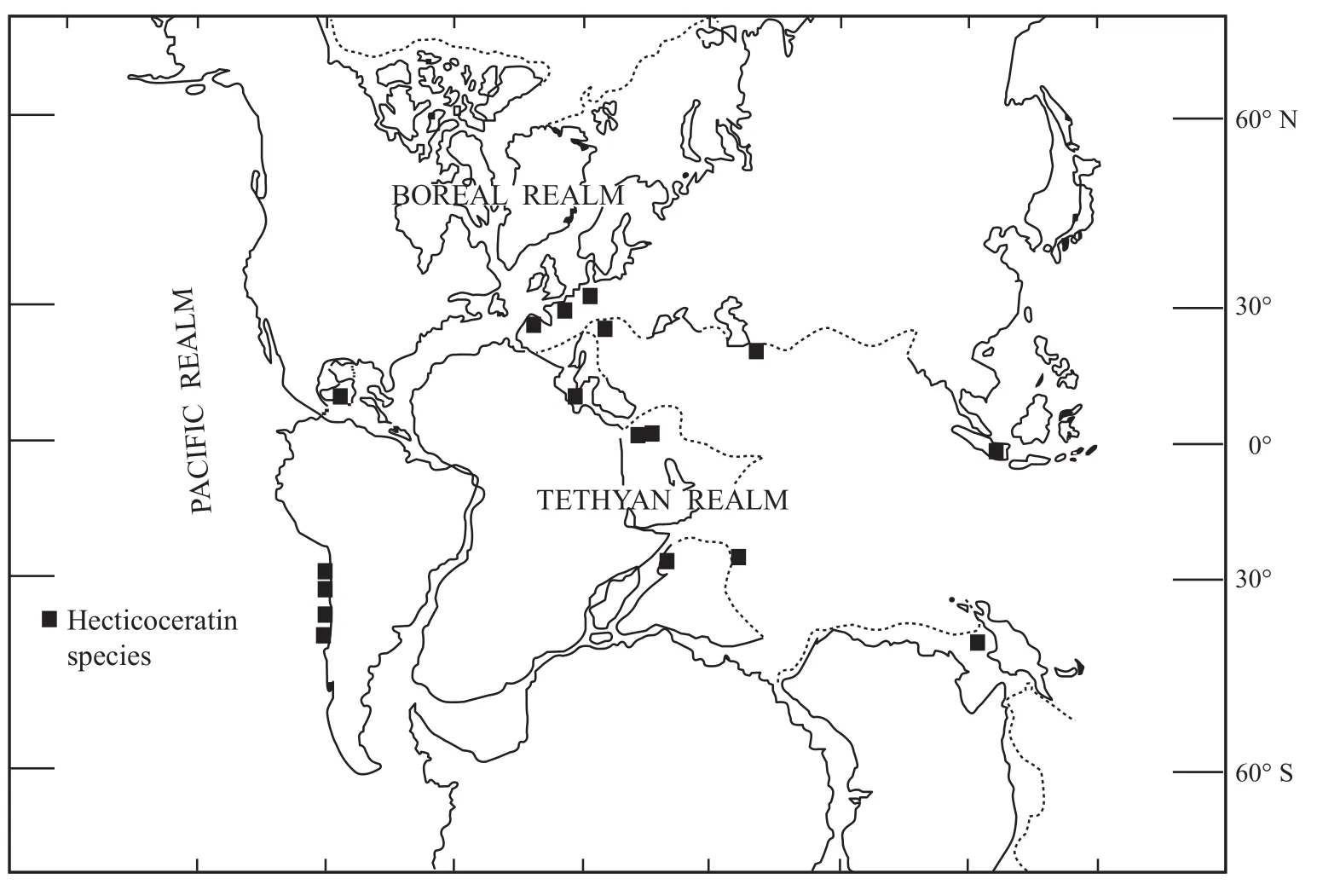
Figure 16 Distribution of subfamily Hecticoceratinae during the Callovian (modified after Hallam, 1975).
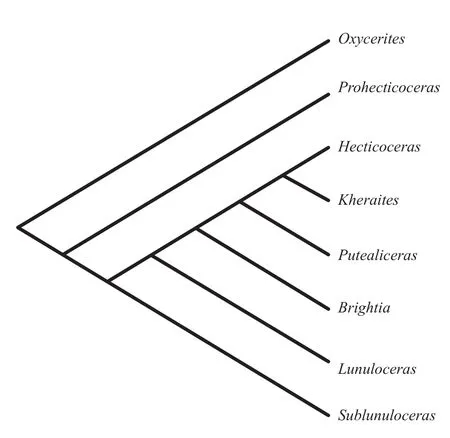
Figure 17 The most parsimonious cladogram showing the phylogenetic relationship of the subfamily Hecticoceratinae (after Roy, 2010).
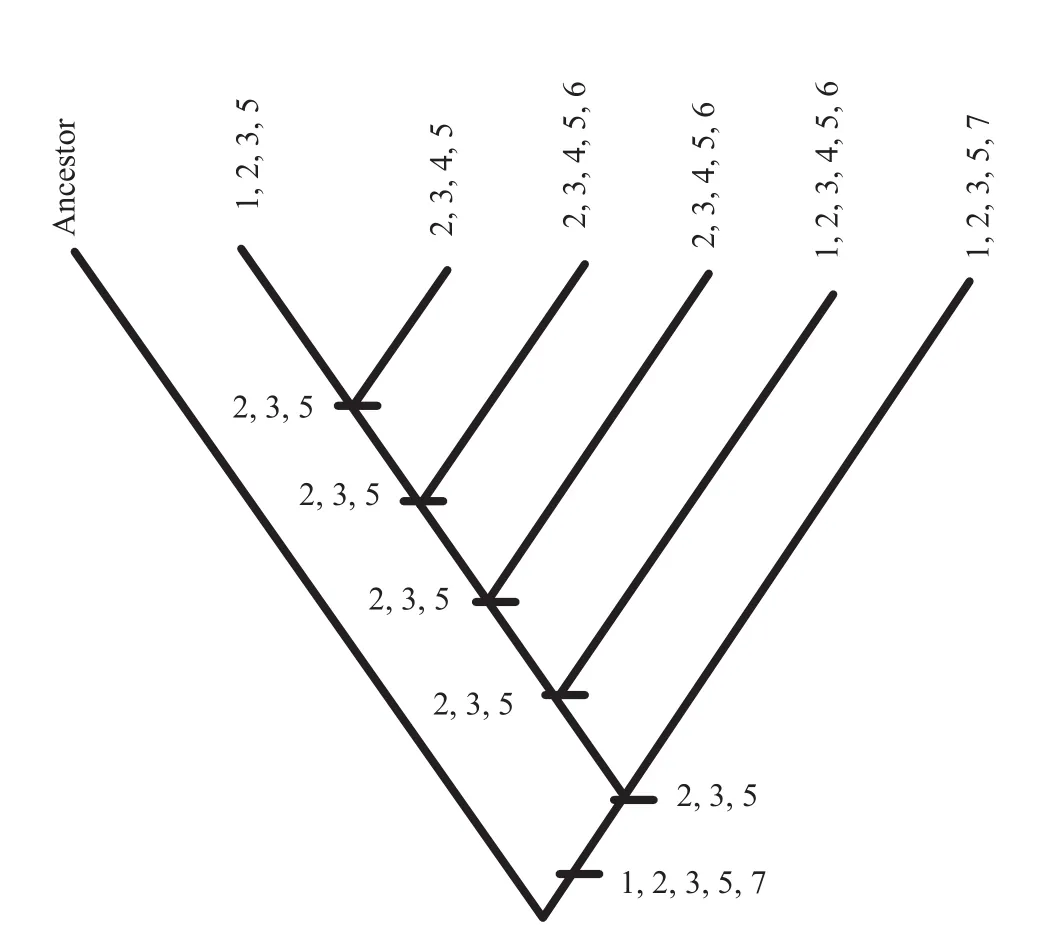
Figure 18 The phylogeny of the subfamily Hecticoceratinae from Roy (2010)with biogeographic states substituted for terminal taxa and mapped onto the ancestral nodes calculated using Fitch optimization (Fitch, 1971).1-France; 2-Switzerland;3-Germany; 4-Israel; 5-India; 6-Iran; 7-South America.
For vicariance analysis, the procedure for coding the data matrix, as proposed by Liberman and Eldredge (1996)was followed in accordance with typical BPA (Brooks,1985; Wiley 1988a, 1988b; but not Brooks, 1990).The biogeographic states of nodes and terminal taxa entered into a data matrix; transitions between adjacent ancestraldescendant nodes and between adjacent ancestral nodes and terminal taxa that involved progressive range contraction or vicariance were treated as ordered multi-state characters following Wiley (1988a, 1988b)and Wileyet al.(1991)(Table 12).For illustrations see Figure 18, where the biogeographic states were mapped onto the phylogenyof Hecticoceratinae.
In dispersal analysis, transitions between adjacent nodes and between ancestral nodes and terminal taxa that involved change or expansion of the range of a lineage(that is, dispersal)were treated as ordered multi-state characters, and these transitions, along with all the other nodes and terminal taxa, were coded into a data matrix (Table 13).This analysis is based on the principle that repeated patterns of range expansion into the same area provide evidence for geo-dispersal (Liberman and Eldredge, 1996).The codings were done in a manner analogous to that in the vicariance analysis.
The data matrix emerged from vicariance analysis when being put into the parsimony analysis through PAUP 4b10 (Swofford, 1990), the seven most parsimonious trees were produced with tree lengths of 16, c.i.0.75 and r.i.0.82 (Figure 19).The g1statistic describing tree-length frequency distributions was -0.8487, a value significantly different from those derived from random data at the 0.01 level of confidence (Hillis, 1991), implying a strong biogeographic signal in the data from our vicariance analysis.
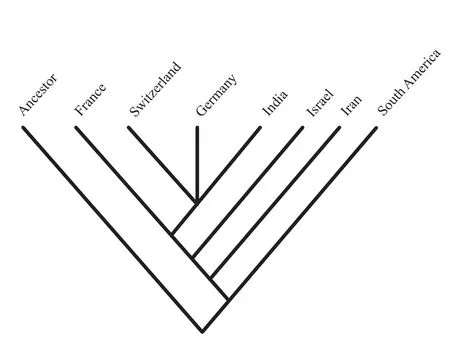
Figure 19 The most parsimonious cladogram showing the area relationship predicted by vicariance analysis.
The area cladogram produced by vicariance analysis shows that basins of the Sub-Mediterranean Province like Switzerland and Germany are closely related to the Kutch Basin of Indo-Madagascan Province, and they form a basal polytomy.The joint area they make up is in turn sister to the France Basin.The Indo-Madagascan Province along with the Sub-Mediterranean Province together forms a group that is sister to the South American Basin of the Andean Province.On the other hand the basins of Israel and Iran emerged as distinct basins.A bootstrap analysis was conducted to assess the relative support for each of the nodes that appear in the most parsimonious tree.Whether or not all the requirements for rigorous statistical application of bootstrap analysis are met (Felsenstein, 1985), it can serve as a qualitative demonstration of relative degree of support for specific cladogram topologies.One thousand replications were employed in a simple step-wise heuristic search that retained groups compatible with the 50%majority-rule consensus tree.The nodes of the tree duplicated in the bootstrap analysis and their confidence values are (Switzerland/Germany/India)= 1.0, and (France)(Switzerland/Germany/India)(Israel/Iran)= 0.52.These bootstrap values indicate that Sub-Mediterranean and Indo-Madagascan area relationships are strongly supported.
The matrix coded for the dispersal analysis, when subjected to parsimony analysis, yields ten most parsimonious trees with a length of 19 steps, c.i.0.84 and r.i.0.88 (Farris, 1989; Figure 20).The g1statistic describing tree length frequency distributions was -0.8826, a value significantly different from those derived from random data at the 0.01 level of confidence (Hillis, 1991), implying a strong biogeographic signal in the data from our dispersal analysis.
The area cladogram produced by dispersal analysis shows that basins of the Sub-Mediterranean Province like Switzerland and Germany are closely related with the Kutch Basin of the Indo-Madagascan Province, and they form a basal polytomy as in the case of vicariance analysis.Here, the joint area they make up is in turn a sister to the Mediterranean basins of Israel and Iran where Israel and Iran are more closely related.The area cladogram depicts that the France basin is a far closer relative of otherSub-Mediterranean, Mediterranean and Indo-Madagascan Provinces.The South American Basin of the Andean Province is totally different in the dispersal analysis.A bootstrap analysis was also performed to assess the relative support for each of the nodes that appear in the most parsimonious tree.Whether or not all the requirements for rigorous statistical application of bootstrap analysis are met, it can serve as a qualitative demonstration of relative degree of support for specific cladogram topologies.One thousand replications were employed in a simple step-wise heuristic search that retained groups compatible with the 50% majority-rule consensus tree.The nodes of the tree duplicated in the bootstrap analysis and their confidence values are similar to vicariance analysis, so the bootstrap values also indicate that the Sub-Mediterranean and Indo-Madagascan area relationships are strongly supported.
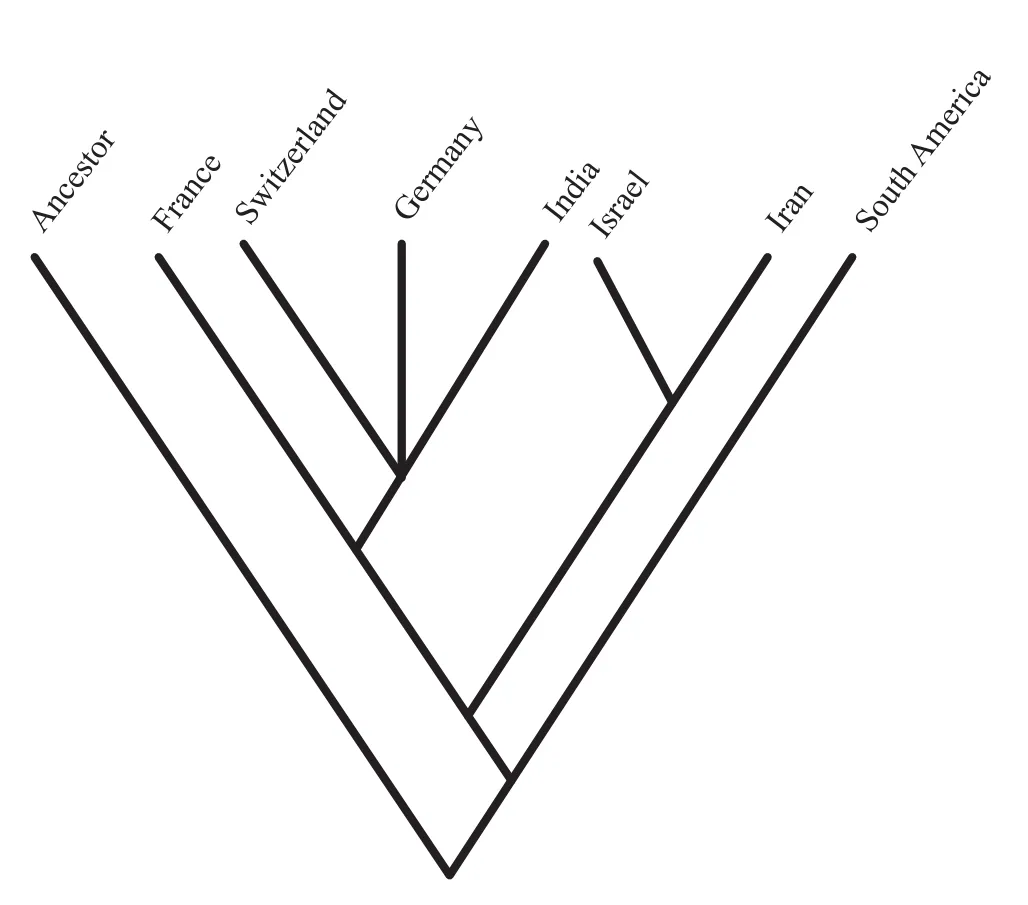
Figure 20 The most parsimonious cladogram showing the area relationship predicted by dispersal analysis.
5 Discussion
Phylogeny of hecticoceratinae clade has been taken into account to evaluate biogeographic patterns of Callovian-Oxfordian hecticoceratins.Parsimony analysis of vicariant palaeobiogeographic data indicate that the Sub-Mediterranean Province is the strongly supported biogeographic region for hecticoceratin diversity during the Callovian.Basins in France were sisters to the Sub-Mediterranean,although less diverse at least during the Middle and Upper Callovian.Hecticoceratin phylogeny shows that the genusProhecticoceraswas the oldest genus of the Hecticoceratinae subfamily (Arkellet al., 1957, Elmi, 1967, Royet al.,2007).Prohecticocerasoriginated during the early Bathonian and the propensity of origination location indicates as Andean Province (South-western Mexico and Northern Chile)which was also regarded as the origin point of the Macrocephalitinae and Reineckeiidae (pers.com.; Callomon and Grǎdinaru, 2005).Prohecticoceraswas near cosmopolitan genus (Hillebrandt, 1970; Sandovalet al., 1990;Hillebrandtet al., 1992).The diversity ofProhecticocerascontinued to the Upper Bathonian and many species have been found throughout Europe as well as Chile, Argentina(Riccardiet al., 1989, Sandovalet al., 1990)and Indonesia (Westermann and Callomon, 1988)and also in India(Royet al., 2007).Both in southern Mexico and northern Chile, the upper Lower CallovianHecticocerass.str.andHecticoceras(Chanasia)(Riccardiet al., 1988, 1989)are present but Middle and Upper Callovian hecticoceratins are scarce except the Upper CallovianHecticoceras(Sublunuloceras)(Parent, 2006).The late Bajocian-early Bathonian Basins of France were well connected through the Hispanic Corridor with that of the Andean Province and faunal exchange was frequent (Moyneet al., 2004).So, in the area cladogram France is a sister to the other Sub-Mediterranean Basins of Germany and Switzerland but during the Middle-Upper Callovian it became distinct with respect to hecticoceratin species.Here is the case where BPA shed some light on the vicariant biogeographic relationships between France and the Andean Province at least with respect to the hecticoceratin lineage.The Kutch Basin of Indo-Madagascan Province is closely related to the Basins of Germany and Switzerland, although this faunal similarity came into being during the Middle Callovian.
Following Liberman and Eldredge (1996)I would also like to refer to the pattern of multiple range expansion of taxa in several clades as geo-dispersal.Geo-dispersal is the phenomenon where dispersal does not occur by crossing of barriers but by the removal of barriers and expansion of range of organisms and here areas rather than fragmenting, would combine into larger areas.Examination of biogeographic patterns in the hecticoceratin phylogeny and in the area cladograms from the dispersal, and vicariance analyses make it possible to determine if geo-dispersal is an important phenomenon in the phylogenetic history of these taxa.In several of the componenets in the area cladogram identical patterns were produced by dispersal and vicariance analyses.In addition, the consensus area cladogram produced by both of the above analyses shows an almost similar pattern (Figure 21).Congruence in the biogeographic patterns indicated by the dispersal and vicariance analyses has interesting implications.For example, patterns from the vicariance analysis predict that the Sub-Mediterranean Province and Indo-Madagascan Prov-ince share a common recent history to the exclusion of the Basins of France (Figure 19).This indicates that there was a progressive isolation of the French Basins from the rest of the Mediterranean Province.The area cladogram from the analysis of dispersal also predicts that, in terms of their dispersal history, the Basins of Germany and Switzerland along with the Kutch Basin share a sister relationship to the exclusion of the Basins of France and all other regions(Figure 20).This implies that the closest dispersal relationship and the most recent shared dispersal history are between the Sub-Mediterranean and Indo-Madagascan and next most recent shared dispersal history is among the Mediterranean, Sub-Mediterranean and Indo-Madagascan provinces.Congruence between patterns of dispersal and patterns of vicariance indicates that in terms of the hecticoceratin clades studied the most recent dispersal history corresponds to the most recent vicariant history.Therefore, the last barriers to come up were the first to fall,and under changing geological conditions, the last areas to differentiate would be the first to merge.This can be explained by relative sea-level rise known to occur during the Bajocian-early Bathonian; relative sea-level fall in late Bathonian followed by a long term rise which started in early Callovian and continued in Oxfordian (Hallam,1978, 2001; Haqet al., 1987).
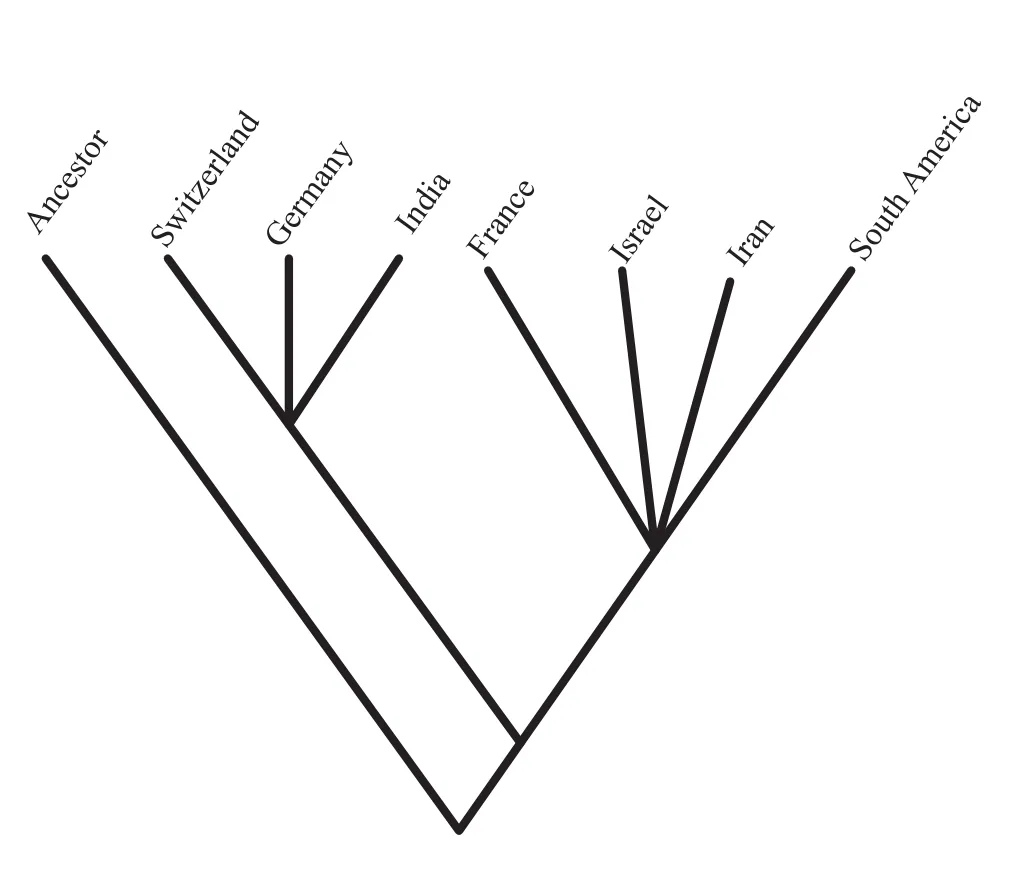
Figure 21 Strict consensus cladogram showing the area relationship predicted by both vicariance and dispersal analysis.
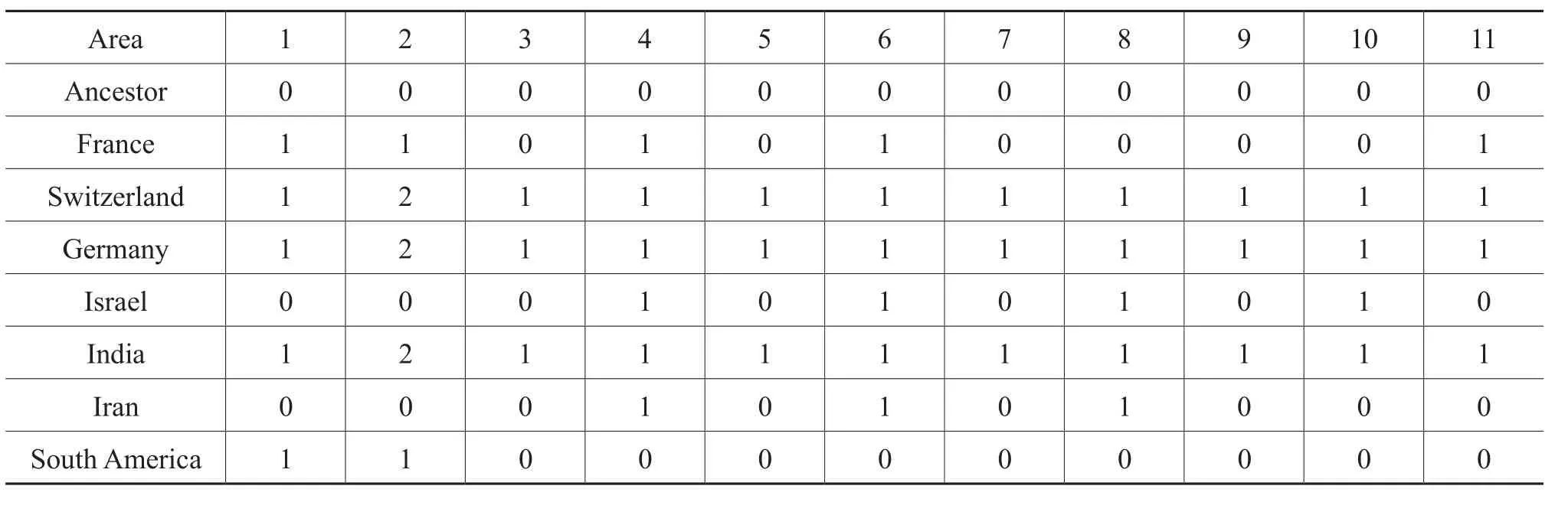
Table 12 Biogeographic character states for the modified Brooks Parsimony Analysis (BPA)data matrix coded for vicariance analysis.Ancestor refers to ancestral biogeographic conditions.
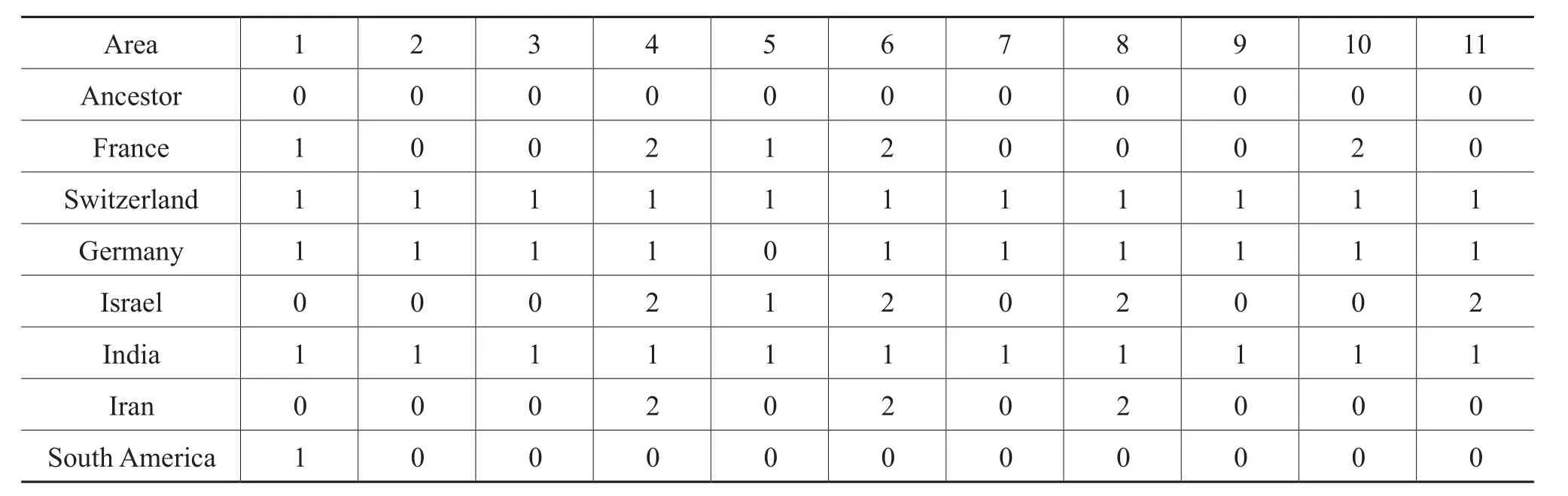
Table 13 Biogeographic character states for the modified Brooks Parsimony Analysis (BPA)data matrix coded for dispersal analysis.Ancestor refers to ancestral biogeographic conditions.
Thus, it appears from the stratigraphic and palaeobiogeographic distribution pattern, as well as BPA, that immediately after the formation of the Kutch Basin, during the early Bathonian relative sea-level rise, immigrant faunas of different provinces expanded their range to the Kutch sea.EuropeanProcerites,Choffatiaalong withProhecticocerasinvaded the Kutch Basin immediately after macrocephalitin taxa reached the Kutch Basin from the West Pacific Faunal Province (Royet al., 2007).Again during the late Bathonian-early Callovian sea level fall the Kutch basin became isolated and the hecticoceratin ammonites developed species level endemicity.From the Middle Callovian onwards the Kutch Basin again experienced a relative rise in sea-level (Biswas, 1977, 1991)and a simultaneous range expansion of hecticoceratin genera especially from the Mediterranean and Sub-Mediterranean provinces.So, species level endemicity in hecticoceratingenera became less distinct during that stage, and it strongly suggests geo-dispersal as a dominant biogeographic pattern during the middle Callovian-Oxfordian.
Acknowledgements
It is a great pleasure for me to express my most sincere gratitude to Professor Subhendu Bardhan for his constant thoughtful supervision and encouragement for this research work.
I am deeply indebted to J.H.Callomon and G.Schweigert for their invaluable suggestions and criticism as well as providing valuable literature.I would like to thank Prof.B.S.Liberman for his valuable suggestions, however, the responsibility for the opinion expressed in the present paper lies entirely with the present author.Thanks are also given to Prof.Ian D.Somerville and Prof.Adam D.Woods for their constructive and important corrections about this manuscript which improved its language.I acknowledge the Director of Repository Section, Geological Survey of India for giving us permission to study the type materials.The help rendered by the local administration of Kutch and the local people of Jumara, Jara and Keera are also acknowledged.The research work was completed under UGC Minor Research Project Scheme [F.PSW-008/11-12(ERO)].
Arkell, W.J., Kummel, B., Wright, C.W., 1957.Mesozoic Ammonoidea.In: Moore, R.C., Teichert, C., (eds).Treatise on Invertebrate Paleontology.Part L.Mollusca.The Geological Society of America and University of Kansas Press, Boulder, Colorado and Lawrence, Kansas, 81-129.
Bardhan, S., Sardar, S., Jana, S.K., 2002.The Middle JurassicKheraicerasSpath 1924 from the Indian Subcontinent.In:Summesberger, H., Histon, K., Daurer, A., (eds).Cephalopods — Present and Past.Abhandlungen der Geologischen Bundesanstalt, A, 57:265-277.
Biswas, S.K., 1977.Mesozoic rock-stratigraphy of Kutch, Gujarat.Quarterly Journal of the Geological, Mineralogical, and Metallurgical Society of India, 49: 1-51.
Biswas, S.K., 1991.Stratigraphy and sedimentary evolution of the Mesozoic basin of Kutch, western India.In: Tandon, S.K., Pant,C.C., Casshyap, S.M., (eds).Sedimentary Basins of India: Tectonic Context.Gyanodoy Prakashan, Nainital, 74-103.
Brooks, D.R., 1985.Historical ecology: A new approach to studying the evolution of ecological association.Annals of the Missouri Botanical Gardens, 72: 660-680.
Brooks, D.R., 1990.Parsimony analysis in historical biogeography and coevolution: Methodological and theoretical update.Systematic Zoology, 39: 14-30.
Callomon, J.H., Grǎdinaru, E., 2005.From the thesaurus of the museum collections.I.Liassic ammonites from Munteana (Svinita Zone, southern Carpathians, Romania).Acta Palaeontologica Romaniae, 5 (2005): 49-65.
Cariou, E., 1984.Biostratigraphic Subdivisions of the Callovian Stage in the Subtethyan province of Ammonites, correlations with the Subboreal zonal scale.In: Michelsen, O., Zeiss, A., (eds).International Symposium on Jurassic Stratigraphy, (I.U.G.S.), 1,Ehrlangen, 315-326.
Cariou, E., Krishna, J., 1988.The Tethyan Reineckeiinae of Kachchh and Jaisalmer (West India): Systematic, Biostratigraphic and Biogeographic implications.Palaeontographica, AbteilungA, 203:149-170.
Datta, K., 1992.Facies, Fauna and Sequence: An integrated approach in the Jurassic Patcham and Chari Formation, Kutch, India.[Ph.D.Thesis].India, Kolkata: Jadavpur University, 167.
Douvillé, R., 1914.Etude sur les Oppelidés de Dives et de Villers--sur-mer.Mém.Soc.Géol.France, 48: 26, fig.33, pl, 1-2 (4-5).
Elmi, S., 1967.Le Lias supérieur et le Jurassique Moyen de L’Ardèche.Documents des Laboratories de Géologie de la Faculté des Sciences de Lyon, 845.
Farris, J.S., 1989.The retention index and the rescaled consistency index.Cladistics, 5: 417-19.
Felsenstein, J., 1985.Confidence limits on phylogenies: An approach using bootstrap.Evolution, 39: 783-791.
Fitch, W.M., 1971.Towards defining the course of evolution: Minimum change for a specific tree topology.Systematic Zoology,20: 406-416.
Fürisich, F.T., Oschmann, W., 1993.Shell beds as tools in basin analysis: The Jurassic of Kachchh, western India.Journal of Geological Society, London, 150: 169-185.
Fürisich, F.T., Pandey, D.K., Callomon, J.H., Oschmann, W., Jaitly,A.K., 1994.Contribution to the Jurassic of Kachchh, western India.II.Bathonian Stratigraphy and Depositional Environment of the Sadhara Dome, Patchchham Island.Beringeria, 12: 95-125.
Hallam, A., 1975.Jurassic Environments.Cambridge: Cambridge University Press.
Hallam, A., 1978.Eustatic cycles in the Jurassic.Palaeogeography,Palaeoclimatology, Palaeoecology, 23: 1-32.
Hallam, A., 2001.A review of the broad pattern of Jurassic sea-level changes and their possible causes in the light of current knowledge.Palaeogeography, Palaeoclimatology, Palaeoecology,167:23-37.
Haq, B.U., Hardenbol, J., Vail, P.R., 1987.Chronology and fluctuating sea-levels since the Triassic.Science, 235: 1156-1166.
Hillebrandt, A.von, Smith, P., Westermann, G.E.G., Callomon,J.H., 1992.Ammonite zones of the circum-Pacific region.In:Westermann, G.E.G., (ed).The Jurassic of the Circum-Pacific.Cambridge: Cambridge University Press, 247-272.
Hillebrandt, A.von, 1970.Zur biostratigraphie und ammoniten-fauna des siidamerkanischen Jura (insbes.Chile).Neues Jahrbuch fur Geologie und Pafaontologie, Abhandhmgen, 136: 166-211.
Hillis, D.M.1991., Discriminating between phylogenetic signal and random noise in DNA sequences.In: Miyamoto, M., Cracraft, J.,(eds).Analysis of DNA Sequences.New York: Oxford University Press, 278-294.
Jain, S., 1997.On the earliest occurrence of genusPhlycticerasHyatt in Kachchh, Western India.Journal of Geological Society of India, 49: 75-80.
Jana, S.K., Bardhan, S., Halder, K., 2005.Eucycloceratin ammonites from the Callovian Chari Formation, Kutch, India.Palaeontology, 48(4): 883-924.
Jeannet, A., 1951.Stratigraphie und Palaeontologie des oolitischen Eisenerzlagers von Herznach und seiner Umgebung.Beitr.Geol.Schweiz,geotech.ser., XIII life., 5 band, 240 p., taf.1-107, 544 text-fig.
Kanjilal, S., 1980.On the stratigraphic range ofSublunulocerasSpath (Ammoniodea Jurassica).Neues Jahrbuch für Geologie und Paläontologie, Mh., 1: 33-36.
Kayal, A., 2009.Palaeobiogeography, systematics and evolution of the Middle Jurassic family Reineckeidae Hyatt and other ancillary ammonite taxa from Kutch, western India.[Ph.D.thesis].India, Kolkata: Jadavpur University, 200.
Krishna, J., 1984.Current status of the Jurassic stratigraphy of Kachchh, western India.In: Michelsen, O., Zeiss, A., (eds).International Symposium on Jurassic Stratigraphy, Erlangen, 3:731-742.
Liberman, B.S., Eldredge, N., 1996.Trilobite biogeography in the Middle Devonian: Geological processes and analytical methods.Paleobiology, 22(1): 66-79.
Martill, D.M., Hudson, J.D., 1994.Fossilien aus Ornatenton und Oxford Clay.Ein Bestimmungsatlas (translated into German by W.Riegraf).Korb (Goldschneck), 224.
Mitra, K.C., Bardhan, S., Bhattacharya, D., 1979.A study of Mesozoic stratigraphy of Kutch, Gujarat with a special reference of rock-stratigraphy and bio-stratigraphy of Keera dome.Bulletin of Indian Geologists’ Association, 12: 129-143.
Moyne, S., Neige, P., Marchand, D., Thierry, J., 2004.Répartition mondiale d’ammonites au Jurassique moyen (Aalénien supérieur à Bathonien moyen): relations entre biodiversité et paléogéographie.Bulletin de la Societé géologique de France, 175: 513-523.
Parent, H., 2006.Oxfordian and Late Callovian ammonite faunas and biostratigraphy of Neuquén-Mendoza and Tarapacá basins(Jurassic, Ammonoidea, Western South-America).Bolétin del Instituto Fisiografia y Geologia, 76(1-2): 1-70.
Parona, C.F., Bonarelli, G., 1897.Sur a faune du Callovien inférieur (Chanasien)de Savoie.Mém.Acad.Sc.Belles-Lettres Arts Savoie, Chambéry, sér.4, t.6: 35-211, fig.a-e, pl.11.
Rajnath, 1932.A contribution to the stratigraphy of Kutch.The Quarterly Journal of the Geological, Mineralogical and Metallurgical Society of India, 4: 161-174.
Reinecke, I.C.M., 1818.Maris protogaei Nautilos et Argonautas,vulgo Cornua Ammonis in Argo Coburgico et vicino reperiundos, L.C.A.Ahlii imp., Coburg, 90.
Riccardi, A.C., Westermann, G.E.G., Elmi, S., 1988.The Bathonian-Callovian ammonite zones of the Argentine-Chilean Andes.In: Rocha, R.B., Soares, A.F., (eds).Proceedings of the 2ndInternational Symposium on Jurassic Stratigraphy, 1987.Instituto Nacional de Investiçào Cientifica, Lisoba, 347-358.
Riccardi, A.C., Westermann, G.E.G., Elmi, S., 1989.The Middle Jurassic Bathonian-Callovian ammonite zones of the Argentine-Chilean Andes.Geobios, 22: 553-597.
Roy, P., Bardhan S., Mitra, A., Jana, S.K., 2007.New Bathonian(Middle Jurassic)ammonite assemblages from Kutch, India.Journal of Asian Earth Sciences, 30: 629-651.
Roy, P., Bardhan, S., 2007.Sexual dimorphism inHecticoceras giganteumSpath (Ammonoidea)from the Callovian (Middle Jurassic)of Kutch, India.Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, 245(3): 263-272.
Roy, P., 2010.Stratigraphy, phylogenetic systematics and evolutionary palaeobiology of the family Oppelidae Douvillé (Ammonoidea)and related ancillary taxa from the Jurassic of Kutch, western India.[Ph.D.thesis].India, Kolkata: Jadavpur University, 190.
Sandoval, J., Westermann, G.E.G., Marshall, M.C., 1990.Ammonite fauna, stratigraphy and ecology of the Bathonian-Callovian(Jurassic)Tecocoyunca Group, South Mexico.Palaeontographica, 210(4-6): 93-149.
Schlegelmilch, R., 1985.Die ammoniten des süddeutschen Doggers.Stuttgart: Gustav Fischer Verlag, 1-284.
Spath, L.F., 1927-1933.Revision of the Jurassic cephalopod fauna of Kachh (Cutch).Memoirs of the Geological Survey of India.Palaeontologia Indicia, New Series 9: 1-945.
Swofford, D.L., 1990.PAUP: Phylogenetic analysis using parsimony, version 4.0 b 6.Smithsonian Institute (Computer Software).
Waagen, W., 1875.Jurassic Fauna of Kutch, the Cephalopoda.Palaeontologia Indica, Geological Survey of India, Series 9, Memoir 1: 1-247.
Westermann, W., Callomon, J.H., 1988.The macrocephalitinae and associated Bathonian and Early Callovian (Jurassic)Ammonoids of the Sula Islands and New Guinea.Palaeontographica,Abteilung A, 203: 1-90.
Wiley, E.O., 1988a.Vicariance biogeography.Annual Reviews of Ecology and Systematics, 19: 513-542.
Wiley, E.O., 1988b.Parsimony analysis and vicariance biogeography.Systematic Zoology, 37: 271-290.
Wiley, E.O., Siegel-Causey, D., Brooks, D.R., Funk, V.A., 1991.The compleat cladist.Lawrence: University of Kansas Museum of Natural History Special Publication 19, University of Kansas.
Wynne, A.B., 1872.Memoire on the geology of Kutch to accompany a map compiled by Wynne A.B.and Fedden F., during the seasons 1867-1868 and 1868-1869.Memoirs of the Geological Survey of India, 9(1).
Zeiss, A., 1956.HecticocerasundReineckeiaim Mittel- und Ober-Callovien von Blumberg (Südbaden).Verlag der Bayerischen Academie der Wissenschaften in Kommission bei der C.H.Beck’schen Verlagsbuchhandlung.München.
杂志排行

Journal of Palaeogeography的其它文章
- Peritidal carbonate cycles induced by carbonate productivity variations: A conceptual model for an isolated Early Triassic greenhouse platform in South China
- Palaeoenvironmental and biostratigraphic implications of microbial mat-related structures:Examples from the modern Gulf of Cambay and the Precambrian Vindhyan Basin, India
- Jurassic sedimentary evolution of southern Junggar Basin: Implication for palaeoclimate changes in northern Xinjiang Uygur Autonomous Region, China
- Restrictions to the application of ‘diagnostic’criteria for recognizing ancient seismites
- Dating recent sediments from the subaqueous Yangtze Delta and adjacent continental shelf, China
- Palinspastic reconstruction and geological evolution of Permian residual marine basins bordering China and Mongolia