高原深水湖泊水温日成层对溶解氧、酸碱度、总磷浓度和藻类密度的影响:以云南阳宗海为例*
2014-09-25袁琳娜杨常亮李晓铭李世玉申时立李智圆刘仍兵
袁琳娜,杨常亮,李晓铭,李世玉,申时立,李智圆,刘仍兵,刘 楷
(1:云南大学生命科学学院,昆明650091)
(2:云南大学工程技术研究院,昆明650091)
(3:昆明市环境监测中心,昆明650028)
水温是湖泊水环境变化的重要驱动因子,水的所有物理化学特性几乎都与水温有关[1].有研究表明[2],大多数温带及亚热带的深水湖泊都会出现水温分层,湖泊水温分层使湖水上下层的交换受抑制,导致湖泊水质分层.湖泊中水温的情况及其分层现象将直接或间接的影响到湖水环境中的各种物理、化学和生物过程[3-5].特别是在深水湖泊中,湖泊水体的垂向温度分布特点和变化规律对水体水质有较大影响,物质的交换能力和湖泊的许多生态动力学过程都受到水温的影响,加之水温本身就是深水湖泊水库水质的最重参数.因此,研究深水湖的水温分层对了解污染物浓度分布和维护湖泊生态系统平衡有重要的作用.
水体理化指标是影响水质的重要因素,比如湖泊水华可通过叶绿素等进行反应和预警.叶绿素的水平反映了浮游植物生物量的多少[6].水温高低及其分层情况会直接或间接的影响到湖泊水环境中的这些理化和生物过程[3,7].张玉超等[8]对浅水湖泊太湖的研究表明其存在日成层现象,太湖DO及pH与水温分层相关.戴凌全等[9]探讨了水库水温预测的经验法和数学模型法,发现水温分层会导致DO、pH、COD等理化参数的分层.吴志旭等[10]研究了新安江水库湖泊区水体的垂向物理、化学参数以及浮游植物群落,结果表明夏季该水库湖泊区垂向pH值、溶解氧及浊度的变化同叶绿素a浓度呈现高度一致.赵巧华等[11]通过气象数据用模型对洱海水温垂直分布进行模拟,发现夏季洱海能维持持续的分层现象,秋季有明显的日分层现象.目前关于浅水湖泊的水温与水体理化参数关系的研究较多较系统[12-13].然而,目前对深水湖泊分层的研究案例较少,大部分为数学模拟研究.对于昼夜温差很大的高原深水湖泊,从上午到下午,水温层次是否发生变化以及呈现什么样的变化规律?湖泊水体中浮游植物、藻类密度、总磷浓度、溶解氧和酸碱度这些理化及生物因子是否随水温的分层也呈现分层现象,这种分层现象是否也随水温层次的变化而呈现出同步的变化呢?在不同的水温层里,这些理化指标是否呈现出不同的变化规律?这是亟待回答的问题,这方面的实测研究案例仍然缺乏.
云南九大湖泊之一阳宗海,为深水湖泊,地处云贵高原.该流域属北亚热带气候,受季风影响明显,气温日较差较大,干湿季节分明[14].选择这样一个日温差大的高原深水湖泊正好满足了我们要研究的上述问题,符合研究需求.阳宗海承担着饮用水源、农业灌溉水源和工业用水水源三大功能,保护标准为《地表水环境质量标准GB/T 3838----2002》Ⅱ类标准.在1992年前阳宗海水质符合地面水质Ⅰ、Ⅱ类标准,但随着该流域人口的增加和工农业的发展,流域水环境逐渐被污染,湖泊变为中营养状态,水质恶化.在1997----1998年间出现“水华”现象,水质降为Ⅳ类,部分水域水质降低到Ⅴ类[15].之后采取了诸如取缔大规模水产养殖等政策措施,取得一些效果,到2004年水质恢复到Ⅱ类水,但2005年以后,流域内逐年增加的散养家禽和家畜的粪便量超过了流域系统的自净能力,使湖泊再次富营养化[16],2011年度,阳宗海湖体水质达Ⅲ类水标准;而阳宗海2012年1----9月水质状况与去年同期相比,水质类别由Ⅲ类下降为Ⅳ类.因此,选择这样一个正在富营养化的高原深水湖泊具有现实意义.
1 材料与方法
1.1 研究区概况
阳宗海(24°51'~24°58'N,102°5'~103°2'E;图 1 ),地跨云南省澄江、呈贡、宜良三县交界,属天然断陷淡水湖泊.呈南北向伸展,南北平均长约12.7 km,东西平均宽约3 km,湖岸线长约34 km,正常运行高水位1769.9 m时,水面面积31.6 km2,相应蓄水量为6.17×108m3,最大水深30 m,平均水深20 m,多年平均水资源量7380×104m3.流域人口327万人,流域以农业和旅游业为经济主体[16-17].
1.2 样点设置
根据阳宗海的基本形状特征,选择了有代表性的6个样点,A、B、C点分布在阳宗海的北、中、南部,A~F点深度分别为 2 2.5、22.5、18.5、14、15、13 m,样点布设如图 1 .
1.3 采样及分析方法
于2012年8月7日(晴天)进行第1次现场采样,11:00到16:00每隔1 h采集A~F点的样品,样品记为A1~F1;于2012年8月30日(晴天)进行第2次现场采样,13:00到16:00每隔1 h采集A~C点的样品(11:00 ---12:00和16:00由于现场偶然因素,未能正常测定),样品记为A2~C2.采用YSI 6600V2-2型多参数水质监测仪现场测定各样点的水深及水温、藻蓝蛋白、叶绿素a(Chl.a)、溶解氧(DO)和pH等水质参数[18-19].每个样点均以垂直1 m为间隔,测定从距水面0.5 m处开始,到距离底泥0.5 m结束,最深的采样点在距离底泥0.5 m以上.用抽水机抽取不同深度(即在距离水面0.5、1.5、2.5、3.5 m……处采取水样)的水作为对应深度的水样.不同深度的所有水样均根据GB/T 11893----1989完成总磷浓度的实验室测定.本研究采用SPSS 16.0进行统计分析,显著性水平α=0.05.
2 结果
2.1 阳宗海夏季各理化参数的水平结构
从整体上看,阳宗海各样点的平均水温、pH、DO、藻蓝蛋白、Chl.a及总磷浓度在全湖的水平分布如图2(数据为8月7日和30日2次测定的平均值、标准差及其多重比较的结果),从A~F点,即随时间从上午到下午,水温逐步上升,水质参数pH、DO以及藻蓝蛋白和叶绿素a浓度都逐步升高.pH在7.8~8.6之间,水体呈碱性,F点pH显著高于 A点(P<0.05),DO 在 D、E和 F点显著高于 A 点(P <0.05),藻蓝蛋白和Chl.a浓度在F点显著高于A点(P<0.05).湖泊总磷浓度虽然由A~C点有增加趋势,但差异不显著(P>0.05),水平分布较均匀,在0.028~0.034 mg/L之间.
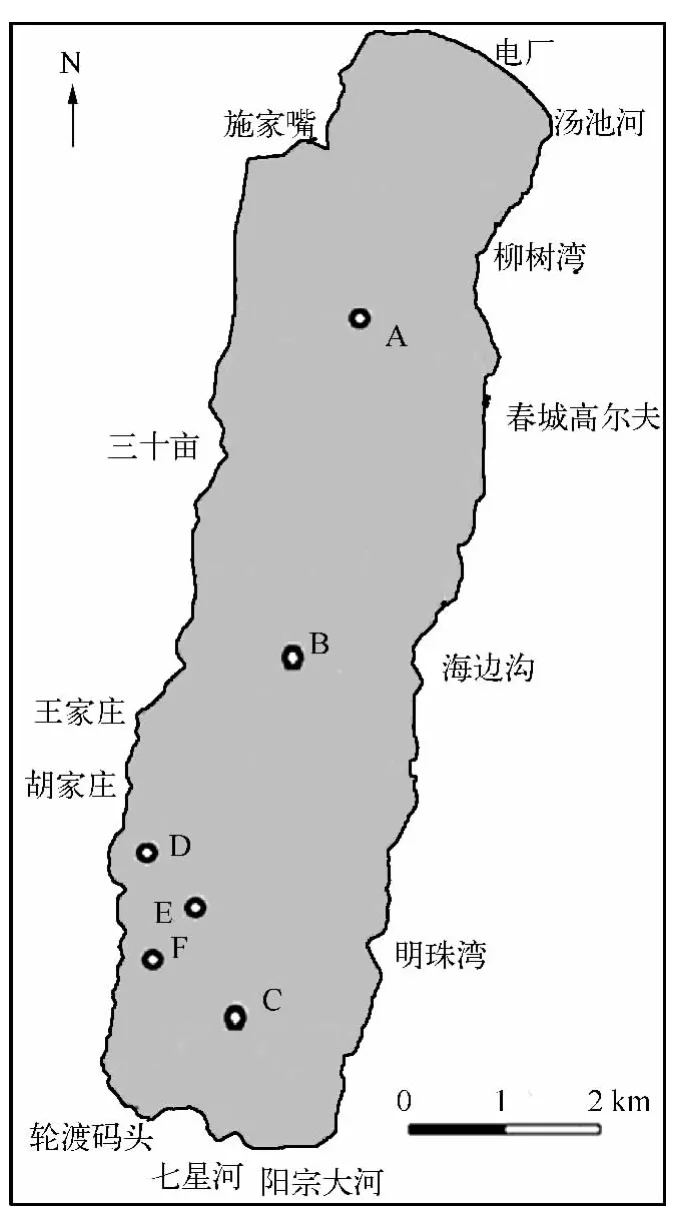
图1 阳宗海采样点分布Fig.1 Distribution of sampling sites in Lake Yangzonghai
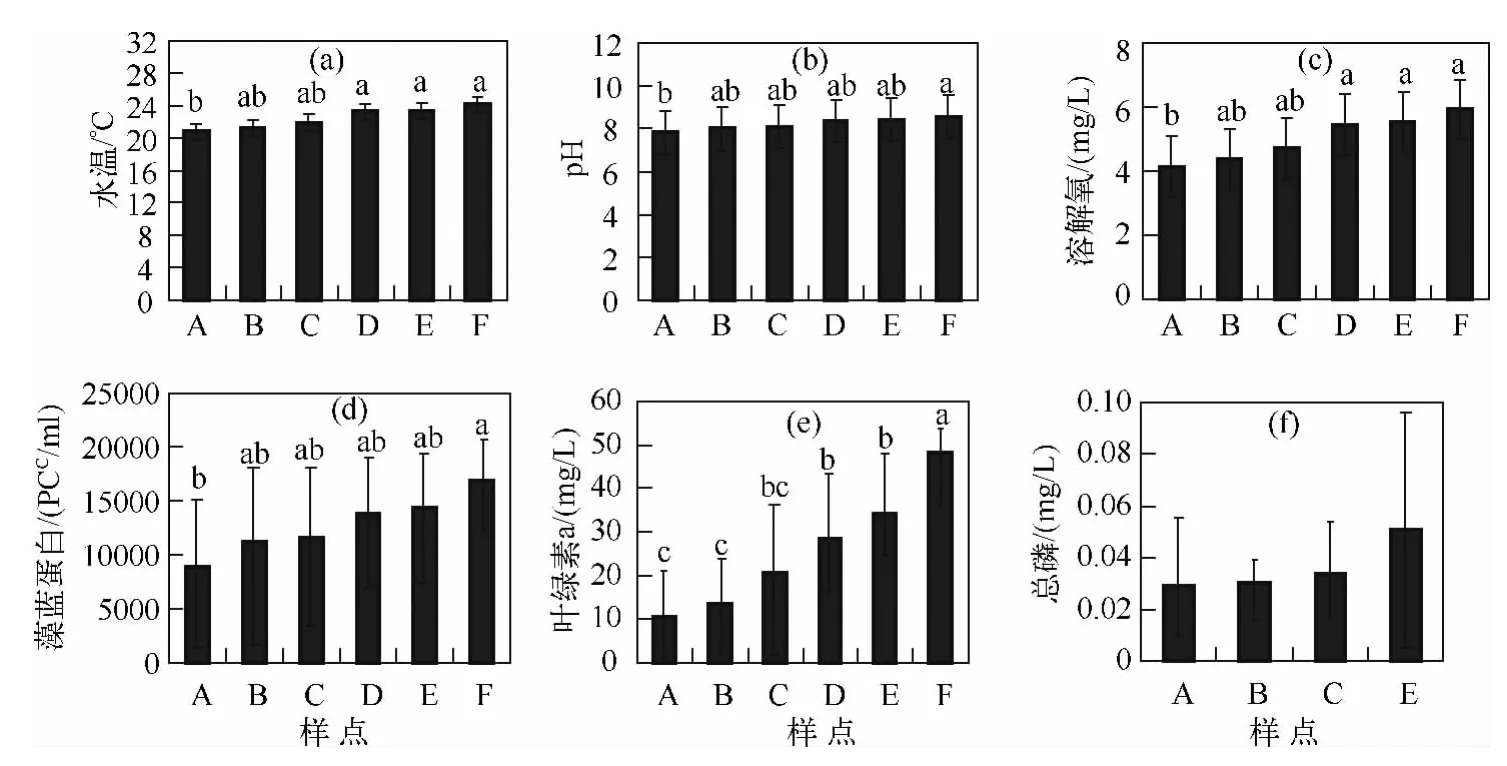
图2 阳宗海各样点平均水温、pH、溶解氧、藻蓝蛋白、叶绿素a和总磷的水平分布Fig.2 Vertical distributions of water temperature,pH,DO,phycocyanin,chlorophyll-a and TP distribution of sampling sites in Lake Yangzonghai
2.2 水温分层及各理化因子的垂直结构
2.2.1 水温的分层及其随时间的变化 8月7日阳宗海湖底到湖面水温在17.1~24.8℃之间.11:00时(A1),大致可以分为4层,但水温层次不明显;12:00时(B1),4层逐渐明显:0~4 m(相对恒温),4~13 m(相对恒温),13~16 m(温跃层)和大于16 m(滞温层);13:00时(C1),水温逐渐形成3层,但不太明显:0~9 m内水温相对恒定,9~13 m水温降低变化较大,13 m以下缓慢降低;14:00时(D1),水温逐渐形成很明显的3层:0~10 m为明显恒温层(24.5℃),10~12 m为温跃层,12 m以下为滞温层;15:00时(E1),水温分层与14:00相似(D1),但是恒温层增加了1 m,而温跃层减少了1 m,即0~11 m为明显恒温层(24.5℃),11~12 m为温跃层,12 m以下为滞温层;16:00时(F1),水温分层与15:00一致(E1),水温分层未显示变化.
8月30日13:00到15:00各理化因子的测定结果见图4,表明水温分为3层:0~3 m为恒温层,3~18 m为变温层,18 m以下为滞温层.13:00到14:00,3~9 m的水温也逐渐升高向恒温层靠拢,但是未能达到恒温层的温度,到15:00,分层开始变模糊.
2.2.2 水体理化参数对水温分层的响应 pH和DO作为温度的函数,在分层过程中呈现出与水温分层近似相同的分层现象和层次结构.从湖底到湖泊表面,DO在2.4~6.8 mg/L之间,水温高的地方DO也高,响应速度很快,基本上达到同步;同理,pH也呈现出同样的规律,高温区pH也较高,响应速度也很快;叶绿素a对水温分层的响应有一定迟效性,在14:00前后,叶绿素a开始出现与水温层次一致的分层,高温层叶绿素a较高,滞温层最低(图3).8月30日的测定结果显示,DO和pH随水水温分层及其变化的响应与8月7日测定的结果一致(图4),而叶绿素a从水面到湖底呈现出一个单峰曲线:3 m内随水深增加而升高,在水面下3 m的地方出现峰值,然后随水深增加而下降,水深9 m以下则变化幅度很小.
藻蓝蛋白对水温分层的响应类似叶绿素a,有迟滞性,在没有达到稳定的分层时,先在水面以下3~6 m的地方出现一个峰值,随水温分层稳定后,在高温区峰值两边的藻蓝蛋白密度渐渐升高,最后在高温层形成一个密度相对均匀的高藻蓝蛋白密度层(图5).TP对水温分层及其变化不敏感,在水面到9 m深的水体TP浓度没有变化,水深较深的地方,水面到水深15 m的水体TP浓度都没有变化,但在接近湖底的几米内,总磷浓度快速升高(图6).
3 讨论
3.1 阳宗海夏季各参数的水平结构分析
湖泊水温的水平分布与水深有直接的关系[20],因此,本文对相似水深的样点进行了比较.本研究中A、B、C点具有相似的水深,从A~F点,随测定时间的推移水温呈逐步升高的趋势:2012年8月7日,各点从南到北湖水均温分别为20.72、21.16和21.83℃,2012年8月30日,各点从北到南湖水均温分别为21.09、21.26和22.33℃;D、E、F 点具有相似的水深,第 1 次测定时 3 个点湖水均温分别为 23.21、23.32 和24.01℃.平均水温在D、E和F点较高,主要因为这几个样点的测定时间是午后,导致水温较高.白天受热水温升高,表层水温比更深层的水温升高快;酸碱度显示水体呈碱性,说明水体可能受到有机污染,呈一定程度的富营养化.本研究中阳宗海溶解氧平均值在4.3~6.2 mg/L之间,稍低于正常情况下地表水中溶解氧浓度(5~10 mg/L),在水藻繁生的水体中,由于光合作用使放氧量增加,也可能使水中的氧达到过饱和状态,湖泊水中一般溶解氧浓度会随水深增加而减少,深层水中甚至完全无氧;藻蓝蛋白和Chl.a在F点显著高于A点(P<0.05),说明南部藻类和浮游生物密度较北部多,南部富营养化较北部严重.从pH(图2b)、藻蓝蛋白(图2d)和Chl.a(图2e)的分布可以推断出:富营养化的有机污染源主要来自阳宗海南部,通过地表水和地下水进入湖泊,是湖泊富营养化的主要贡献部位.
3.2 阳宗海夏季各参数的垂直结构分析
3.2.1 水温的分层及其随时间的变化规律 阳宗海水温分层存在日变化,而不是简单地分为3层,上午随气温开始上升,水温分层由不明显变得明显,由最初的4层渐渐变成3层(图3),随着气温的升高,湖上层即恒温层不断扩大,同时,温跃层逐渐减小,因为温跃层的上部接近恒温层的水层,水温逐渐升高变成恒温层.由于样点水深差异,A ~C 点水深较深(18.5~22.5 m),D ~F点水深较浅(12.5~14.5 m),这可能造成稳定后恒温层深度的差异.第1次测定时E、F点恒温层深度最大达到水面下11.5 m(图3);第1次和第2次测定时C点恒温层深度最大均达到水面下10 m深度(图3、图4).夏季阳宗海的湖泊区水温层相对稳定后,在9~13 m处形成温跃层,湖上层与深水层的温差最大为7.8℃,温跃层深度较新安江水库湖泊浅(在10~20 m处形成温跃层[10]),表层和底层水温差也较小(新安江水库为20℃[10]).阳宗海8月初白天水温分层明显,但到8月末,白天水温分层已经不明显,水温层模糊,并在出现2 h后就基本消失.太阳辐射强度和湖面风速等气象条件是影响日成层产生和强弱的主要因素[3,13],因此,恒温层和温跃层的大小受太阳辐射强度和湖面风速的控制,水温分层在每天15:00能够达到相对稳定状态.

图3 2012年8月7日样点A~F的水温、DO、pH和Chl.a的垂直分布Fig.3 Vertical distributions of water temperature,DO,pH and Chl.a of the sites A ~ F on August 7,2012
3.2.2 各参数对水温分层的响应规律 阳宗海的水温分层能够引起DO、pH、浮游植物和蓝藻的分层.在本研究中,DO和pH呈现出与水温分层近似相同的分层现象和层次结构,响应速度很快,达到基本上同步.在淡水中,DO的溶解度主要取决于温度[21],而浮游植物的光合作用会改变水体碳酸盐平衡,使pH升高,另一方面,浮游植物的光合作用深受季节的影响,即夏季温度的影响[22],且温度和DO本身就是温度的函数,本研究中pH与DO的变化均与温度的变化一致,支持了前人的研究结论[23].叶绿素和藻蓝蛋白对水温分层的响应有一定迟效性,滞后的时间约2 h(图3和图4).叶绿素被广泛认为是间接反映浮游植物生物量的参数,但在一定程度上与藻类的组成有关[24],浮游植物细胞、藻蓝蛋白的数量在垂向分布上湖上层高于深水层,温跃层以下细胞数急剧减少,水温分层决定着浮游植物的垂向分布[5].本研究显示,在没有达到稳定的分层时,先在水面下恒温层与变温层交界的地方出现一个峰值,随水温分层稳定后,在高温区峰值两边的叶绿素或藻蓝蛋白密度渐渐升高,最后在高温层形成一个密度相对均匀的高密度层(图3、图4和图5).从8月初到月末,叶绿素a浓度呈下降趋势(图3和图4).
TP对水温分层及其变化不敏感,可能是夏季的水温分层会导致混合层与深水底质之间的联系受湖泊分层的影响而减弱,从而致使包括氮、磷和硅在内的营养盐进入真光层能力减弱[21].但TP浓度在接近湖底的几米内显著升高(图6),可能是因为夏季水温分层及消失的过程中,水动力会引起一些底泥里的磷释放到水体里,造成湖泊底部的水体TP浓度高[25].8月末,水温分层不明显,并在16:00时分层渐渐消失(图4),而此时,9 m以上的水层总磷浓度并没有增加,说明当前时间段磷内源的影响不大,仍然保持在湖底部附近.但不能排除在风强的天气,沉积物释放的磷可能由于风浪引起的湖水运动而把湖下层中的磷带入湖上层中,导致湖上层磷浓度升高[21],另外,室内模拟的风浪扰动实验证明,大风浪扰动后水体TP浓度大幅增加,且恢复到扰动前水平至少需要10 d[26].
本研究通过分析发现阳宗海水体温度存在分层现象,DO、pH、藻蓝蛋白都存在与水温层对应的分层,蓝藻对水温分层变化的响应有迟滞性.藻类生长需要的最基本条件是温度[27],温度不但决定细胞内酶促反应的速率,还是藻类进行光合作用的必要条件,且与植物的合成代谢及呼吸强度密切相关[28].而湖上层藻蓝蛋白密度较大(图5)是因为4 m/s以下的小风浪有利于蓝藻生长或漂浮于表层水体[29],且2次监测阳宗海风速均为5.4 m/s以下,对蓝藻在湖上层的漂浮有一定影响.因此,浮游植物蓝藻与湖泊水温分层有密切关系,它们对水温响应的迟效性也与诸多化学过程有关.

图4 2012年8月30日各样点水温、DO、pH和Chl.a的垂直分布Fig.4 Vertical distributions of the water temperature,DO,pH and Chl.a of the sites on August 30,2012
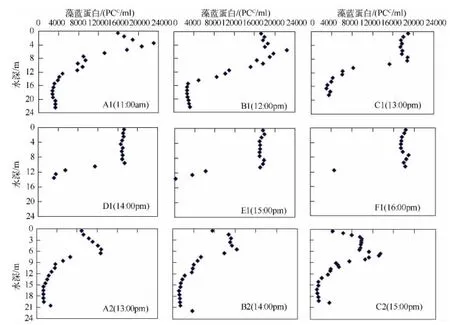
图5 藻蓝蛋白在不同水深的垂直分布Fig.5 Vertical distribution of phycocyanin in different water depths

图6 2012年8月30日总磷在不同水深的垂直分布Fig.6 Vertical distribution of TP in different water depths on August 30,2012
4 结论
阳宗海水温在夏季出现分层现象,从上午开始逐渐形成4层,午后形成3层,并在14:00时达到分层的相对稳定,稳定时温跃层出现在水深9~13 m处,湖水表层与湖底的温差最大为7.8℃,滞温层很难受到白天高温的影响;DO、pH、叶绿素a和藻类密度随水温的分层出现分层,其中,DO和pH响应迅速,而叶绿素和藻类密度的响应有迟效性,迟滞约2 h才能达到对应的分层;藻类和浮游植物的成层过程是先在高温层出现一个峰值:藻类在水深3~7 m,水温23.3~23.8℃之间的区域出现一个峰值,而浮游植物在水深1~4 m,水温24.1~24.9℃之间的区域出现一个峰值.然后在周边的区域藻蓝蛋白和浮游植物密度逐渐增加而最终形成一个与水温对应的层次;在晴朗风小的天气,水温分层基本上不影响总磷的分布,全湖泊的总磷平均浓度为0.033±0.03 mg/L,总磷浓度只有在接近湖底的地方迅速升高,在之上的垂直剖面上的分布较均匀,说明当前状态下水温分层及消失过程并不会造成上层水的磷浓度升高,磷内源对上层水没有影响.而在风强大浪的天气条件下的结果仍然具有不确定性,有待于进一步研究.
[1]王雅慧,李 兰,卞俊杰.水库水温模拟研究综述.三峡环境与生态,2012,34(3):29-36.
[2]Han B,Armengol J,Carlos GJ et al.The thermal structure of Sau Reservoir(NE:Spain):a simulation approach.Ecological Modeling,2000,125(2):109-122.
[3]张玉超,钱 新,钱 瑜等.太湖水温分层现象的监测与分析.环境科学与管理,2008,33(6):116-121.
[4]隋 欣,杨志峰.龙羊峡水库蓄水对水温的净影响.水土保持学报,2004,18(4):154-157.
[5]林 佳,苏玉萍,钟厚璋等.一座富营养化水库——福建山仔水库夏季热分层期间浮游植物垂向分布.湖泊科学,2010,22(2):244-250.
[6]阮晓红,石晓丹,赵振华等.苏州平原河网区浅水湖泊叶绿素a与环境因子的相关关系.湖泊科学,2008,20(5):556-562.
[7]王松波,薛庆举,高 光等.新疆博斯腾湖浮游甲壳动物群落结构.湖泊科学,2011,23(6):926-933.
[8]张玉超,钱 新,石川忠晴等.浅水型湖泊水温日成层现象的初步探讨——以太湖为例.四川环境,2008,27(3):45-48.
[9]戴凌全,李 华,陈小燕.水库水温结构及其对库区水质影响研究.红水河,2010,29(5):30-35.
[10]吴志旭,刘明亮,兰 佳等.新安江水库(千岛湖)湖泊区夏季热分层期间垂向理化及浮游植物特征.湖泊科学,2012,24(3):460-465.
[11]赵巧华,孙绩华.夏秋两季洱海、太湖表层混合层的深度变化特征及其机理分析.物理学报,2013,62(3):1-9.
[12]王得玉,冯学智,周立国等.太湖蓝藻爆发与水温的关系的MODIS遥感.湖泊科学,2008,20(2):173-178.
[13]赵林林,朱梦圆,冯龙庆等.太湖水体理化指标在夏季短时间尺度上的分层及其控制因素.湖泊科学,2011,23(4):649-656.
[14]祝 艳.阳宗海流域环境背景状况.环境科学导刊,2008,27(5):75-78.
[15]杨常亮,陈建中,贺 彬等.阳宗海总磷环境容量与总磷超标的原因分析.环境科学导刊,2008,27(1):44-46.
[16]Yang CL,Li SY,Yuan LN et al.The eutrophication process in a plateau deepwater lake ecosystem:Response to the changes of anthropogenic disturbances in the watershed.2012 International Conference on Biomedical Engineering and Biotechnology(iCBEB),2012,3:1815-1821.
[17]Yang CL,Sun PS,Li SY et al.Analysis of non-point pollution sources and countermeasures of Lake Yangzonghai watershed.Desalination and Water Treatment,2013(Accepted).
[18]Friedrich J,Oberhänsli H.Hydrochemical properties of the Aral Sea water in summer 2002.Journal of Marine Systems,2004,47(1-4):77-88.
[19]Uncles RJ,Stephens JA,Harris C.Towards predicting the influence of freshwater abstractions on the hydrodynamics and sediment transport of a small,strongly tidal estuary:The Devonshire Avon.Ocean and Coastal Management,2013,79:83-96.
[20]张运林,陈伟民,杨顶田等.天目湖热力学状况的监测与分析.水科学进展,2004,15(1):61-67.
[21]Kalff J.Limnology:Inland water ecosystems.Canada:Pearson Education,2002:155,162,229,258.
[22]秦伯强,许 海,董百丽.富营养化湖泊治理的理论与实践.北京:高等教育出版社,2011:117.
[23]危起伟,柯 福,恩庄平等.富水水库温度分层的调查研究.海洋湖沼通报,1991,3:12-18.
[24]Calijur MC,Santos AC.Temporal changes in the phytoplankton community structure in a tropical and eutrophic reservoir(Barra Bonita,SP-Brazil).Journal of Plankton Research,2001,24:617-634.
[25]吴宗义.湖泊内源磷释放及形态转化的影响理化因素.化学工程与装备,2009,4:134-135.
[26]高永霞,孙小静,张战平等.风浪扰动引起湖泊磷形态变化的模拟试验研究.水科学进展,2007,18(5):668-673.
[27]孙慧群,朱 琳,高文宝.淡水湖泊中微囊藻水华的成因分析.生物学通报,2005,40(8):23-24.
[28]陈宇炜,秦伯强.太湖梅梁湾藻类及相关环境因子逐步回归设计和蓝藻水华的初步预测.湖泊科学,2001,13(1):63-71.
[29]孙小静,秦伯强,朱广伟等.风浪对太湖水体中胶体态营养盐和浮游植物的影响.环境科学,2007,28(3):506-511.
