广西田林浪平地区晚石炭世生物礁古生态特征
2014-09-10巩恩普杨大勇陈晓红关长庆
巩恩普, 杨大勇, 陈晓红, 关长庆
东北大学地质系,沈阳,110004
内容提要:广西田林县浪平地区发育了几处黄龙期生物格架礁。这些发育在海百合碎屑滩上的礁体规模普遍不大,造礁和附礁生物种类多样,礁生物群落成熟度低,造礁方式单一,具有斑块式分布特点。浪平地区黄龙期点礁是在缺乏全球性主要后生骨架造礁动物的背景下发育的,没有明显优势造礁生物出现;点礁受研究区内大规模发育的碎屑滩影响显著,成礁环境多变、高能水体与碎屑选择作用明显、固结和硬质基底分布不广泛、水体能量频繁变化与适应环境的海百合繁盛、藻类在研究区内不发育,这些使得浪平地区点礁具有典型的“碎屑滩点礁模式”特征;造礁生物的生态特点对点礁的具体面貌具有重要影响。晚石炭世碎屑滩后生动物骨架点礁的发现,有助于进一步加强生物礁古生态系统研究。
泥盆纪和二叠纪是晚古生代礁体的繁盛时期,生物礁类型多样、数量和规模可观(Kiessling et al.,1999,2002),右江盆地发育有多个泥盆纪或二叠纪生物礁(曾允孚,1983;吴亚生等,1989;陆刚等,2006) 。石炭纪是作为这两个截然不同的礁群落生态系统的中间时期,构造运动复杂,古地理、古气候发生重大变化(Saunders et al.,1986;Stanley,1988;Wood,1998),被认为是全球性造礁的衰落阶段(巩恩普等,1998;West,1988 ;钟建华,2005 ),其生物礁研究也相对薄弱。我国早、晚石炭世都有典型的后生动物骨架礁发育( 方少仙等,1985;巩恩普等,1998, 2003;关长庆等,2007;常洪伦等,2011 ),研究基础较好。最近在广西田林县浪平乡又发现了几处石炭纪生物礁,本文以其中晚石炭世黄龙组礁体为研究对象,结合野外工作和室内研究结果,分析浪平地区晚石炭世生物礁的类型及其生长发育过程、古生态特征等,以进一步认识晚古生代生物礁的发展和演化规律。
1 礁体概况
广西浪平地区发现的较为成型的晚石炭世礁体有3个,分别位于龙江垌村、新寨村附近(表1)。浪平地区大地构造位置归属于“滇黔桂盆地”,盆地南侧与古特提斯洋连通(陈洪德等,1994),石炭纪发育了若干被发育深水相的台沟分割形成堑垒相间格局的孤立碳酸盐台地(秦建华等,1996),导致台地岛弧和边缘台地发育,为浅海生物群落定殖、繁盛提供了适宜场所。3个礁体地层层位都属于上石炭统黄龙组,产出时代为晚石炭世巴什基尔期(图1)。石炭纪时浪平碳酸盐岩台地主要处于浅滩环境,堆积了大量以海百合屑为主的生物碎屑和内碎屑,形成各种颗粒岩、泥粒岩及粒泥岩。
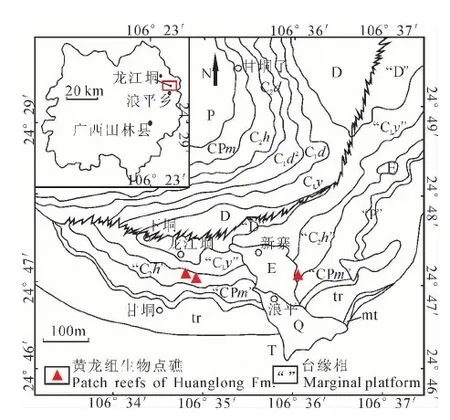
图1 广西田林浪平地区黄龙组生物礁露头地质图 (据地矿部贵州地勘局,1997❶)Fig. 1 The reef outcrops geologic map of Huanglong Group in Langping, Tianlin, Guangxi (After Guizhou Metallurgical Prospecting Bureau of the Ministry of Geology and Mineral Resources, 1997❶)
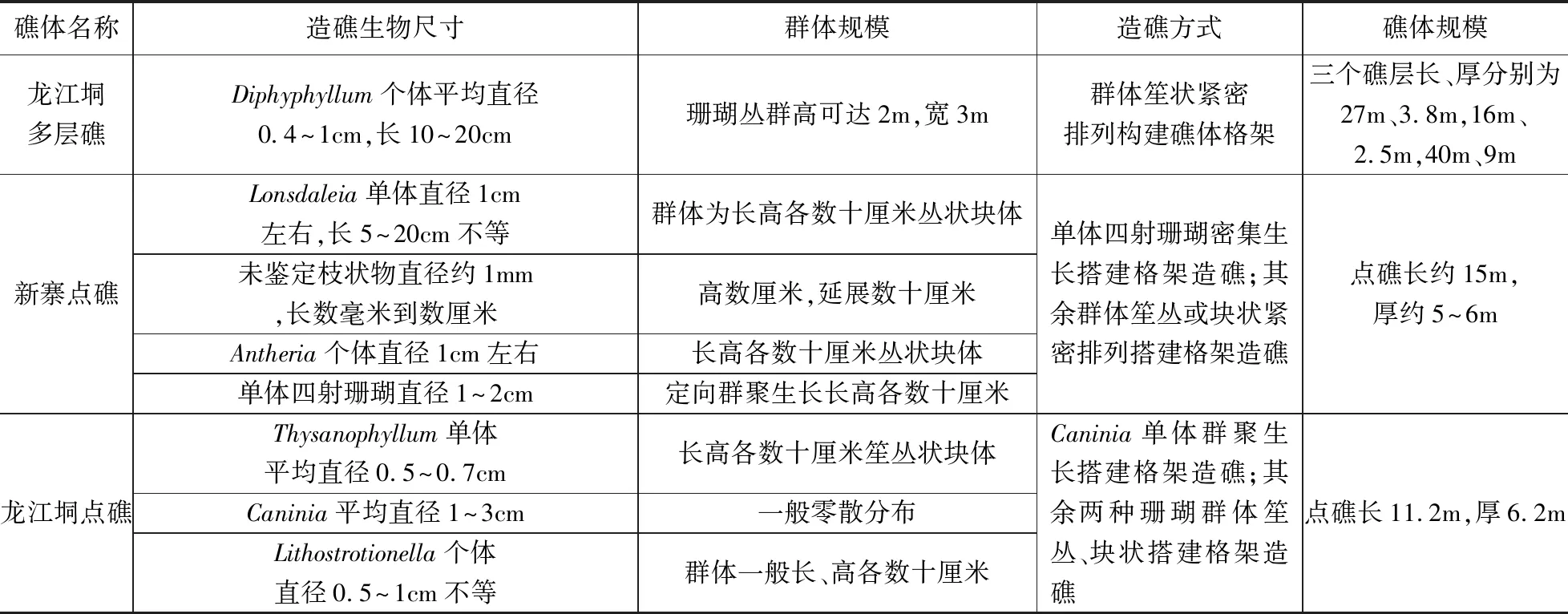
表1 广西田林浪平地区黄龙组点礁概况Table 1 The patch reefs intro of Huanglong Group in Langping, Tianlin, Guangxi

图2 广西田林浪平地区龙江垌 多层礁生长示意图Fig. 2 Idealized model of the Longjiangdong reef in Langping area, Tianlin, Guangxi
1.1 龙江垌多层礁
多层礁分三层发育(图2)在碳酸盐岩台地边缘,近东西向展布,礁层出露长、厚分别为27m、3.8 m(经计算后真实厚度,以下同),16 m、2.5 m,40 m、9 m。造礁生物为群体丛状四射珊瑚Diphyphyllum,个体平均直径0.4~1cm左右,长度10~20cm,以出芽方式簇状生长,个体间排列紧密(图3a),发育在以海百合茎为主的生物碎屑滩之上(图3b),搭建格架造礁;珊瑚丛规模大小不一,最大规模的高度可达2m,宽3m,多呈直立簇状。珊瑚丛群内部适于其他生物的生存空间小,多被灰泥充填,发育有大小不等的栉壳结构;珊瑚丛群障积和改变四周环境作用明显,丛群之间开阔处生活有大量的海百合、有孔虫、腕足等附礁生物。在海水动荡程度高于珊瑚耐受程度的情况下,Diphyphyllum被限制发育甚至在礁区范围内消失;当水动力条件适宜时,Diphyphyllum能迅速扩散,占据基底。三个礁层的生物群落结构和微相特征相似。

图3 广西田林浪平地区龙江垌多层礁造礁生物和基底Fig. 3 Reef-building organism and substrate of Longjiangdong reef in Langping area, Tianlin, Guangxi (a)Diphyphyllum珊瑚群丛;(b)以海百合茎为主的生物碎屑滩 (a) Diphyphyllum group; (b) crinoid stems bioclastic beach
1.2 新寨点礁
发育在新寨村东侧的点礁出露长约15m,厚约5~6 m,可明显分出三个造礁层(丘)(图4)。首先是粘结藻类与发育在泥粒岩基底上的群体笙状Lonsdaleia分别粘结灰泥、搭建格架形成碳酸盐岩建隆,形成点礁的底部。未定名藻类以丝状体形式出现,相互叠置粘结灰泥形成旋卷纹层构造,呈带状展布,规模不大,长、高约数十厘米;Lonsdaleia单体直径1cm左右,长度5~20cm不等,个体间距0.2~3cm(图5a),群聚生长形成长、高各数十厘米的丛状块体。未鉴定枝状物与群体角柱状珊瑚Antheria生长在底部之上构成点礁的第二礁层。未鉴定枝状物个体纤细,直径1mm左右,长数毫米到数厘米,个体间距变化大,以灰泥充填为主,呈小丛状搭建格架构成礁体,呈带状断续延展长约15m,斜距高约2m,周围以灰泥沉积为主(图5b),略早于Antheria出现;Antheria排列紧密,个体直径1cm左右,块体规模不大,长、高各约数十厘米(图5c)。在礁体上部,单体四射珊瑚(由于后期蚀变强烈暂未鉴定属种,图5d)群聚生长占据优势成为礁体的主要建造者,珊瑚个体直径1~2cm,定向排列密集生长,形成出露3~5m宽的礁体第三礁层(丘)。
新寨点礁不同造礁层(丘)之间被长时间的碳酸盐岩沉积分隔,即使在同一造礁阶段中不同造礁生物都是各自占据一定区域,未见不同生物之间形成共生、互利等共同栖息的种群关系。整个点礁是三个先后生长有不同造礁生物层面的简单叠加,这反映了该区黄龙期复杂多变、不利于稳定造礁的环境。
1.3 龙江垌点礁
该点礁(图6)发育在碳酸盐岩台地边缘以海百合茎为主的棘屑滩上(图7a),礁体规模不大,露头长11.2m,厚6.2m,造礁生物为群体笙状珊瑚Thysanophyllum、单体犬齿珊瑚Caninia和块状群体角柱珊瑚Lithostrotionella。Thysanophyllum个体纤细,平均直径0.5~0.7cm,排列紧密(图7b);Caninia单体弯锥状或圆锥状,平均直径1~3cm,一般零散分布,但也见小规模集中生长现象(图7c);Lithostrotionella排列紧密,几乎没有空隙,个体直径0.5~1cm不等,块体规模不大,一般长约25cm,高约15cm(图7d)。龙江垌点礁自下而上造礁生物出现一定的演替,由单独以群体丛状Thysanophyllum造礁演变为Thysanophyllum与单体珊瑚Caninia共同造礁,最后演变为角柱珊瑚Lithostrotion单独造礁,显示了造礁生物随环境变化而发生的演化。珊瑚群丛内部多以灰泥充填,有少量纤细海百合茎,珊瑚群丛周边均为以海百合茎为主的碎屑滩,其中含有大量的腕足壳,碎屑颗粒破碎程度差别很大,反映了当时水体总体动荡且水体动能变化较大的环境。龙江垌点礁不同的造礁生物之间仍然是简单的组合,没有形成具有特定结构、展现各自功能,能够维持长期稳定且广泛分布的造礁生物群落;造礁方式也仅为搭建格架造礁;属于未成熟礁。
1.4 其他底栖生物居群
除点礁外,在浪平地区发育的大规模碎屑滩中以斑块形式分布着一些规模大小不一的其他底栖生物居群(图8)。这些生物居群仅是底栖生物的集中群居生活,并未形成真正意义上的礁或丘,但可以看作是礁体发育的雏形和基础。

图4 广西田林浪平地区 新寨点礁生长示意图Fig. 4 Idealized model of the Xinzhai patch reef in Langping area, Tianlin, Guangxi
2 礁体生态特征及分析

图5 广西田林浪平地区新寨点礁造礁生物Fig. 5 Reef-building organisms of Xinzhai patch reef in Langping area, Tianlin, Guangxi (a)Lonsdaleia;(b)未鉴定枝状物;(c)Antheria;(d)单体柱状四射珊瑚群聚生长 (a) Lonsdaleia; (b) not identify branches; (c) Antheria; (d) grown together single column coral
浪平地区黄龙组生物礁造礁生物多达9种,附礁生物丰富多样,但是每个点礁中生物种群之间只是较为简单的共同栖息生活,缺少稳定、复杂的种间关系或是能对造礁起支持或促进作用的的种间影响机制;简单种群组合对环境变化的抵抗能力非常弱,在水体动能或其他条件有较大变化时,容易失去造礁主体地位甚至在造礁区域内消失;三个点礁中除了偶见未名藻类粘结灰泥外,主要是珊瑚搭建格架造礁,缺少其他造礁方式和其他生物辅助共同造礁,造礁方式简单;点礁主要由不同规模的丛群状斑块式分布的造礁生物组成,丛群规模普遍不大;礁体建造过程难以清楚区分出不同生长阶段,甚至新寨点礁只能被看作是多处造礁生物丛群的集合和叠加,礁体成熟度不高。
同为巴什基尔期的美国俄克拉荷马州生物骨架礁(Sutherland et al.,1977)厚1~2m,长2~3m,由多种藻类和后生动物形成了较复杂的群落生态系统,能区分出由定殖、拓殖、统殖到消亡的生长阶段,礁体建造过程中既有蓝细菌的粘结作用,又有苔藓虫的结壳贡献,还有多种后生动物搭建格架,造礁方式多样。浪平地区黄龙组珊瑚点礁与其相比虽然礁体规模大,但在造礁生物群落成熟度、造礁方式多样性、礁体生长阶段性等方面都明显较差。
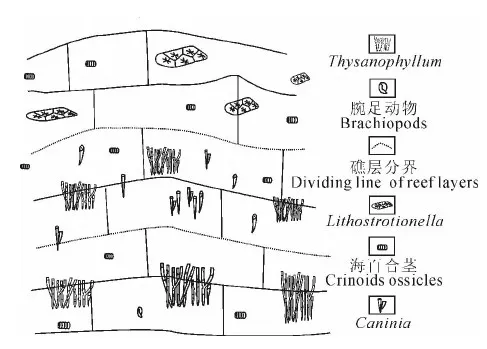
图6 广西田林浪平地区龙江垌点礁生长示意图Fig. 6 Idealized model of the Longjiangdong patch reef in Langping area, Tianlin, Guangxi

图7 广西田林浪平地区龙江垌点礁基底及造礁生物Fig. 7 Reef-building organisms and substrate of Longjiangdong patch reef in Langping ara, Tianlin, Guangxi (a)海百合碎屑滩;(b)Thysanophyllum;(c)Caninia;(d)Lithostrotionella; (a) crinoid stems bioclastic beach; (b) Thysanophyllum; (c) Caninia; (d) Lithostrotionella
石炭纪被认为是后生动物骨架礁发育的衰落期,以丛状、笙状群体或块状复体为主的珊瑚造礁在世界范围内仅发现不多的几处。广西浪平地区的黄龙组点礁具有典型的石炭纪珊瑚骨架礁色彩,与泥盆纪层孔虫—珊瑚骨架礁和二叠纪钙质海绵—钙藻成礁模式相比,自身特点鲜明。
2.1 礁群落发育的地质背景
早、晚石炭世之交的谢尔普霍夫期曾发生了对生物多样性缺失和生态系统重新构建两方面的生态影响高于奥陶纪末期大灭绝的“新的全球第五大灭绝”(图9),引发了泥盆纪F—F事件后残留在石炭纪早期的四射珊瑚动物群的生态学重组(George et al.,2012)。这次灭绝事件在珊瑚礁生态系中也反映得十分明显,大约有90%以上的种类绝灭了,而且有一个长期的、渐进式的复苏过程(童金南,1995)。巴什基尔期的广西浪平地区造礁珊瑚是从大绝灭后的残存者和复活类型中分化出来的(童金南,1995),它们在晚石炭世生态系统复苏中起了重要作用。

图8 广西田林浪平地区其他底栖生物居群Fig. 8 Other benthos groups in Langping area , Tianlin, Guangxi (a)腕足动物群;(b)单体珊瑚群;(c)苔藓虫;(d)粘结藻类 (a) brachiopoda group; (b) solitary corals group; (c) bryozoon; (d) adhesive algae
早石炭世晚期至晚石炭世晚期,主要位于南半球的冈瓦纳大陆持续北移与位于赤道附近的欧美大陆接近,横跨赤道的特提斯洋成为西端收缩闭合、东端宽阔的楔形,环赤道洋流受阻,引起许多门类底栖生物数量和分异度的明显下降乃至大量灭绝(王治平等,1988)。石炭纪晚期礁生态系统缺乏具有普遍代表意义的以大型后生动物为主的全球性的造礁生物群落(Ross et al.,1985;巩恩普等,2010)。
晚石炭世是广西石炭纪海侵范围最大的时期,整个广西基本处于远岸广海环境中,碳酸盐台地生物滩相分布广泛,四射珊瑚明显衰落,其属种数与个体总数大为减少(广西壮族自治区地质矿产局,1999),其他底栖生物繁盛。这样的地质背景奠定了广西浪平地区晚石炭世造礁珊瑚群落难以大规模繁盛发育的基础。
从野外岩性分析、生物化石识别、统计和微相特征研究等结果看,广西浪平地区的海洋物理环境总体上代表了适合底栖生物群落发育的海水清澈、温度和盐度适宜、阳光充足、含氧量丰富、水体动能多变的环境。研究区所在的滇黔桂盆地一直存在断裂的频繁活动,引起等效海平面的升降,对当时面积只有约七百平方公里浪平台地(方少仙等,1985)的水深及水文条件影响显著,碎屑滩非常发育。点礁主要处于孤立碳酸盐台地内动荡潮下带,形成由造礁珊瑚点礁代表的较深水环境与棘屑腕足颗粒岩组成的浅滩环境间互相出现的情况。以笙丛状珊瑚为主的造礁生物,仅能定殖在海水较深的相对低能区域。浪平地区适合成礁的区域还是非常受限制的。
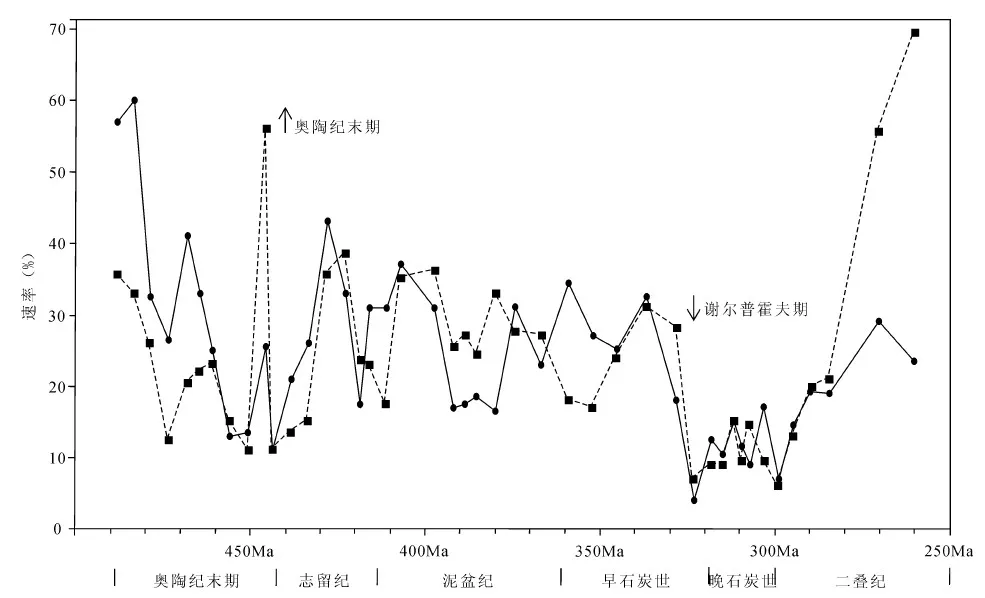
图9 奥陶纪末期和谢尔普霍夫期生物多样性危机对比图(据George et al.,2012)Fig. 9 The biodiversity crises contrast of the end-Ordovician and Serpukhovian (After George et al., 2012) 横轴代表地质年代,纵轴代表灭绝速率(虚线和方形点)和新生繁衍率(实线和圆点)。奥陶纪末期和谢尔普霍夫期生物多样性的不同 影响由箭头显示:向上箭头代表奥陶纪点处灭绝率突然升高,向下的箭头代表谢尔普霍夫阶点处的物种形成率骤降 Horizontal axis is geologic time, and vertical axis is magnitude of extinction rate (dashed line and square data points) and origination rate (solid line and circular data points). Different effects of the end-Ordovician and Serpukhovian biodiversity crises are emphasized by arrows: upward-pointing arrow for Ordovician points out abrupt elevation of extinction rate, and downward-pointing arrow for Serpukhovian pointing out precipitous drop in origination rate
2.2 碎屑滩的影响
晚石炭世早期是广西棘皮类动物发育的极盛期,棘皮类死亡后骨骼近原地堆积可形成数十米至百米以上厚度的大规模棘屑滩堆积层(Thomas et al.,2006),形成典型的滩相沉积。通过野外实地勘察测量可知,广西浪平地区海百合碎屑滩出露面积占据明显优势,能达50%~80%,代表该区主要为地形相对平坦、水体能量较高的浅水动荡环境。在碎屑滩中,发育有很多出露面积不等、微相类型差异显著的区域(图10)和多种生物骨架岩、粘结岩,是该区水体动荡水平变化明显、水体深浅不同、有机物含量差别显著等物理环境变化的直接证据。正是这些多样性的环境,为底栖生物多样性和点礁的形成提供了物质基础。
由于以海百合茎为主的粗粒碎屑随海水涌动游移很容易将纤细的枝状生物和以横向延展生长的生物撞击破碎、迅速埋没致死,浪平地区黄龙组地层内后生骨架动物多以群体笙丛状或是复体块状形式出现,枝状骨架生物仅在较深静水区发现有少量苔藓虫,覆盖方式生长生物未见。海百合碎屑滩和高能量水体环境的共同选择作用使得造礁生物形貌特征近似,由于造礁生物的形貌特征与造礁方式关系密切(吴亚生,1992),这造成浪平地区点礁的造礁方式单一。
固结基底和硬质基底是海底稀缺资源也是底栖造礁生物定殖的唯一基础(陈源仁,1985;Boucot,1991)。从野外点礁露头分析看,浪平地区黄龙组发育有一些泥粒岩同沉积胶结基底、管状生物结壳硬质基底、藻类及微生物结壳硬质基底、钙质生物碎屑硬质基底(图11)。这些固结或硬质基底与游移碎屑滩相比所占比例非常小,分布不广泛,不利于造礁生物群聚生长以抵抗强水流的冲击。由于珊瑚幼虫只能在固结或硬质基底上定殖,这些小面积斑块状硬质基底分布的方式决定了造礁珊瑚也以同样的方式分布。这种斑块状分布生长方式是浪平地区造礁生物的典型特点。

图10 广西田林浪平地区碎屑滩中其他微相类型Fig. 10 Other microfacies types in the clastic beach in Langping area, Tianlin, Guangxi (a)厚层灰泥团块,长时间静水环境下的沉积产物,缺乏生物的扰动痕迹;(b)生物碎屑质灰泥岩,成分以灰泥为主,代表了潮下带低能环境,生物数量稀少;(c)灰泥质生物碎屑颗粒岩,含有大量各类生物碎屑颗粒,代表潮下带中等能量环境,生物丰富多样;(d)层状灰泥粘结岩,以粘结藻席为主导的生物建造,低—中低能量水体环境下的产物;(e)内碎屑颗粒灰岩,内碎屑为圆状,反映了在高能环境中复杂的改造作用;(f)底栖动物碎屑杂积岩,碎屑以腕足、腹足为主,碎屑较完整、分选、磨圆不明显,代表了低能体系下快速沉积环境;(g)近原地海百合碎屑堆积,海百合茎较完整,粗、细肢体和碎屑、灰泥堆积在一起,疑似为原地碎屑坍塌后快速沉积的产物 (a) thick-layer plaster lumps, the deposition product of long time hydrostatic environment, lack of biological disturbance signs; (b) bioclastic wackestone, composed mainly with lime mud, which represents a low-energy subtidal environment, organisms scarce; (c) bioclastic packstone, containing various substantial bioclastic grains, on behalf of medium energy subtidal environment, bio-diverse; (d) layered lime mud boundstone, a biological construction dominated by adhesive algae mats, low — low and medium energy water environment product; (e) endogenous fragment grainstone, endogenous fragment is rounded, reflecting the complex transformation in the high-energy environment; (f) mixed bioclastic rocks, which are main brachiopods and gastropods, conserved completely and not obvious sorting and rounding, represents a low-energy system for rapid deposition environment; (g) near situ crinoid debris accumulation, crinoids stems conserved completely, thick, thin limbs and debris, lime mud piled together, may be the product of rapid deposition after debris collapsing
由于高能浅水环境水体动荡、水体能量变化频繁,适宜造礁生物生长的时间间隔短。浪平地区的礁体发育过程中频繁被抑制或是中止、破坏,难以出现由稳定造礁生物群落形成的大型骨架礁;三个点礁多是由规模普遍不大的珊瑚丛群组成,丛群分布分散、占据面积小,在多变的水体动能影响下,很容易在造礁过程中被迫停止生长甚至在点礁内消失,令浪平地区点礁具有整个礁体由不同单元或阶段组合而成的特点。
浪平地区海百合类分布广泛、个体数量极为丰富,它们固着于底质之上(Grimmer et al.,1984;闫旭等,2010),滤食水流中有机物生长(Thomas et al.,1992;Kerri et al.,2002),对环境的适应范围非常宽泛,与底栖后生动物争夺共同的生存空间以及食物资源。海百合数量巨大,能在一系列环境下优势生长(杨式溥,1993),抑制着其他后生动物生长,这是广西晚石炭世早期底栖后生动物不能大规模繁盛的重要原因。
由于水流频繁扰动和未固结的碎屑滩难以为附着微生物和藻类所接受,在浪平地区没有大规模“藻类草皮”(K. H. 曼等,1989)出现的明显证据(图12)。自养微生物和藻类能为底栖动物提供大量的有机物颗粒食物、复杂多样的栖居环境,诱导底栖动物幼虫附着和变态(雷新明等,2012),能为反硝化菌提供理想的缺氧环境(雷新明等,2012)。微生物和藻类的稀少造成礁群落环境简单、微生态环境氮循环受到阻碍,不能有效大量吸引和承载更多后生动物,物种多样性相对低。钙质微生物和藻类稀少使得粘结和捆绑、缠绕等重要辅助造礁方式在浪平地区极为少见,造礁方式单一,礁体骨架受强水流影响显著、稳固性能差,更没有形成像华南二叠纪钙质藻类—钙质海绵(陆廷清等,1998;吴亚生,1994)和现代南海珊瑚藻—石珊瑚(王国忠,2001)类似的成礁模式。这是浪平地区点礁生物群落对环境频繁变化抵抗能力差的主要原因。
浪平地区三个点礁发育地质背景近似,环境因素的影响作用类同。研究区最主要的地层特征——大规模海百合碎屑滩发育,为浪平地区晚石炭世生物礁的生态特征定下了最基本的格调。浪平地区点礁具有特有的“碎屑滩点礁模式”特征:环境多元化、底栖生物丰富多样;造礁生物斑块式分布,种类多样但形貌近似、造礁方式单一;造礁生物群落成熟度低,受环境影响明显;礁体规模变化大、阶段区分不显著,分布局限。
2.3 造礁生物种群的生态特点
浪平地区的造礁珊瑚基本都属于生态对策中的K对策者(孙儒泳等,2002),种群数量稳定、寿命长,生物个体较大,周边繁殖和侧方繁殖方式使亲代具有一定的后代保护机制(王志根,1989)。这些特点令浪平地区造礁珊瑚具有一定的竞争能力,能够在较长期适宜环境下成为优势种,搭建格架造礁。但K对策生物缺乏较强的繁殖和迁移、扩散能力,在频繁变化的环境中难以利用暂时有利的生境迅速繁盛而形成巨大的种群,尤其对于来自各方面的干扰不能很快做出反应以适应环境的需要。这些生态特点给造礁珊瑚在浪平地区复杂多变的环境下繁育和造礁带来相当大的困难,需要遇到较长时间的“机遇期”才能形成礁丘。
浪平地区的造礁生物进入点礁中新的适合其生长的环境后,没有出现某一种类能迅速扩散成为普遍优势种或适应多种不同生境而广泛分布的情况;点礁新鲜断层面上的侵入者更多的是以抽彩式竞争方式(周淑荣等,2004)随机出现。K对策生物的这些生态特点是龙江垌点礁和新寨点礁中出现多种不同的造礁生物但没有哪一种处于明显优势的原因。

图11 广西田林浪平地区固结和硬质基底Fig. 11 Consolidated and hard substrate of Langping area, Tianlin, Guangxi (a)灰泥质生物颗粒岩,生物碎屑颗粒包括海百合茎片(C)、有孔虫(F)、钙藻碎屑(AF)、苔藓虫(B)、以及似球粒(P)和软体动物壳碎片(MF),生物碎屑首先被等厚刀刃状亮晶胶结物(白色箭头所指)环绕,碎屑之间的孔隙由亮晶胶结物(黑色箭头所指)充填;(b)生物碎屑颗粒岩,碎屑颗粒破碎明显,大小杂积在一起,大个体生物碎屑之间栉壳状胶结物发育(白色箭头所指);(c)管状生物结壳硬质基底在研究区偶有发现,管状物为圆柱形的,有可能为蠕虫管;(d)藻类粘结生物碎屑及团块形成的固结基底;(e)核形石硬质基底;(f)珊瑚丛生长在钙质生物碎屑硬质基底上 (a) biopackstone, the bioclastics includ the crinoid term fragments(C), foraminiferas(F), calcareous algae fragments(AF), bryozoans(B), peloids(P) and mollusk shell fragments(MF), bioclastic earlier grow around equal thickness blade-like sparry cements (white arrows) , the pores of debris are filled by the sparry cement (black arrow); (b) bioclastic grainstone, clastic grains are broken obviously, all of them cumulate together complexly, pectinate texture cement between the large individual bioclastics develops well (white arrows); (c) the hard substrates crusted by cylindrical tubes which are possible the worm tube occasionally are found in the study area, the tubes are cylindrical; (d) the consolidated substrate formed by algae cohering bioclastics and clumps; (e) oncolite hard substrate; (f) coral grows in calcareous bioclastic hard substrate
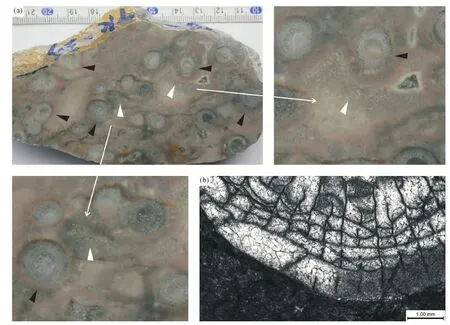
图12 广西田林浪平地区微生物和藻类发育情况Fig. 12 Microorganisms and algae growth conditions in Langping area, Tianlin, Guangxi (a)丛状珊瑚(内部水体动荡水平相对较低)个体四周出现厚度差别明显的微生物和藻席粘附灰泥形成的包裹物(黑色箭头所指),包裹物出现在碎屑沉积(白色箭头所指)之前,说明其在珊瑚是活体时就已存在;(b)单偏光显微镜下显示单体珊瑚边界清楚、平滑,没有微生物或藻类粘附、结壳、钻孔等的痕迹。(a)只出现在密集生长的丛状珊瑚内部,数量少,微生物和藻类对珊瑚影响不显著(吴义布等,2013);(b)在浪平地区非常常见 (a) different thickness wrappage formed by microbe and algal mats cohering lime mud appear around fasciculate corals (black arrows); (b) single polarizing microscope shows that the boundary of single corals is distinct, smooth, no traces of microbe or algae adhering, crusting and drilling. (a) only appears in densely growing fasciculate corals, a small number of microbe and algae had no significant effect on corals (Wu Yibu, et al.,2013); (b) is very common in Langping

图13 珊瑚侧方繁殖Fig. 13 Lateral increase of corals
从Lonsdaleia、Thysanophyllum、Lithostrotionella和Diphyphyllum等群体珊瑚的骨骼形态分析,可将其分为丛状复体和多角柱状复体两类。丛状复体的个体排列稀疏,相互间留有空隙,外具表壁,基本营侧方繁殖。繁殖时,子体的基部与母体连接,当芽体跌落到基质上时与母体分开,平行母体或与母体呈一小角度向外侧生长(图13),整个复体呈笙状或树枝状。多角柱状复体珊瑚排列紧密,个体间具公共的双层外壁,主要以体壁内部边缘带分异进行周边繁殖。繁殖时,子体在母体边缘带中逐渐膨胀,与母体间具公共骨骼,直至一个新的柱壁形成,新体腔完全与母体分离。
通过分析可知,营侧方繁殖的丛状珊瑚比营周边繁殖的多角柱状珊瑚能更有效的占据相对多的基底面积,其所形成的珊瑚礁丛规模一般也较大。在浪平地区,丛状珊瑚种类多于角柱状珊瑚,生物数量也远远大于后者。但无论是哪种生殖方式,都是以母体为中心的、逐步拓展的斑块状分布方式,这使得浪平地区造礁珊瑚丛群规模扩展非常缓慢,丛群规模普遍不是很大。并且由于点礁中珊瑚个体纤细,很容易受到强水流的扰动而被迫中止造礁,稳定造礁持续时间短。在地层上表现出这些珊瑚形成的礁层规模和厚度都不大。生物造礁规模是造礁生物对环境适应能力的重要表现,造礁生物越是能长时间地、更充分地适应环境,对应的礁体规模也越大。生物礁的规模与造礁生物的生长速度、个体(复体)形态、造礁方式、对环境的适应能力以及与附礁生物的生态关系等直接相关。
珊瑚个体粗大、生长迅速,生物骨量就大,能迅速搭建起一定规模的格架,继而营造出适合珊瑚幼体和其他附礁生物定殖、生长,有利于礁体进一步发展的局部环境。这种环境的形成对于大个体珊瑚来说是快速的、相对容易的;小个体珊瑚则需要长时间的生物骨量的积累,在水体环境多变的情况下,这个期间内很内容易被不利的外界条件打断。同时,珊瑚个体粗大、群体生长,能具有很强的抗浪性,一旦它们建立起一定规模的格架,就不容易受外界条件左右,很快进入良性循环发展阶段。
从另一个角度讲,礁体建造规模对造礁生物群落持续发育的影响也是显而易见的。规模巨大的礁体能够更加有效的阻挡水流,为群体内部个体和亲代营造更好的发育环境,保证种群长时间繁盛。同时,大型礁体不同部位环境差异显著,具有多样的生境,能为不同种类生物提供栖息环境,容易形成具有复杂生态关系的造礁生物群落,提高群落的抗扰动能力和恢复能力,增强群落的稳定性(王国宏,2002)。由于点礁规模尤其造礁珊瑚丛群规模普遍偏小,浪平地区礁体发育受环境干扰严重;三个点礁中珊瑚丛群大小不同,受环境影响的程度也有所区别。
3 结论
广西浪平地区黄龙组生物礁是晚石炭世早期发育的后生动物骨架点礁,它们的发现和研究是对石炭纪造礁生物研究的重要补充。碎屑滩中点礁的发育更是具有自身典型的特点,通过对点礁和造礁生物群落古生态特征的详细研究,进一步深入认识了生物礁及礁群落与环境之间的相互关系,得到以下几点认识。
(1)浪平地区黄龙组点礁具有典型特征:造礁生物多样,附礁生物丰富,点礁群落成熟度低、造礁方式单一;造礁生物斑块式分布,受环境影响明显;礁体生长大多明显分层,被碳酸盐沉积间隔,层(丘)规模变化大,分布不广泛。
(2)点礁是在当时缺乏全球性的主要后生骨架造礁生物的背景下发育的,由于生态演化、地质条件等因素的限制,没有出现明显优势造礁生物。
(3)碎屑滩对浪平地区礁群落及点礁的形成具有最直接的、最主要的影响作用,为研究区内晚石炭世生物礁的生态特征定下了最基本的格调。
(4)造礁生物本身的生态特点是影响点礁发育的重要因素。
注 释 / Note
❶ 地矿部贵州地勘局.1997. 1∶50000中华人民共和国区域地质图说明书(水塘幅,G48E015017). 内部资料.
