滇池水华束丝藻(Aphanizomenon flos-aquae)对低氮的生理响应*
2014-08-28吴艳龙郑凌凌殷大聪代龚圆宋立荣
吴艳龙,郑凌凌,李 林,殷大聪,代龚圆,宋立荣
(1:中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,武汉 430072) (2:中国科学院大学,北京 100049) (3:长江水利委员会长江科学院,武汉 430010)
蓝藻水华是世界范围内的热点研究问题,已报道的有几十个属的上百种蓝藻可以形成水华[1].出现最广泛、最常见的水华蓝藻为微囊藻属(Microcystis),因此微囊藻水华已经被广泛关注和深入研究[2].水华束丝藻(Aphanizomenonflos-aquae)也是一种水体中较常见的水华蓝藻种类,在世界各种类型的湖泊、水库、池塘等水域均有发生[3-6].水华束丝藻属于蓝藻门念珠藻科束丝藻属,是我国最早发现的束丝藻属种类,束丝藻水华在我国各地均有报道,并逐渐引起人们的广泛关注.
1982年12月至1983年1月以及4月,东湖浮游植物水华经鉴定为水华束丝藻水华[7].杭州西湖1981年发生水华束丝藻水华,生物量高达67.73×107cells/L,占浮游植物总量的98%,出现西湖历史上罕见的“黑水”[8].贵阳地区最大的人工湖泊红枫湖水库1998年3月-4月上旬出现罕见的大面积水华束丝藻水华最为典型,其间南湖水域表层0.5m左右形成藻浆[9].2002年起,湖北省通山县县城附近的一座小型水库——四斗朱水库连续几年在早春即开始发生严重束丝藻水华,富藻层厚度达4m,水华的唯一优势种为水华束丝藻[10].云南高原湖泊滇池2001-2009年调查研究发现,水华束丝藻与微囊藻属随季节演替,成为滇池蓝藻水华的两大优势种群[11-12].
影响水华束丝藻发生水华的因素很多,如外部营养物质的输入、周边的生态环境、浮游动物种群、具漂浮能力的气囊、固氮异形胞、能够形成孢子、广泛的水温适应性等[4,6,13-15].浮游植物异形胞固氮对固氮蓝藻水华有重要影响,水体中磷营养充足、氮成为限制因子将导致固氮蓝藻水华的发生[16-20].湖泊、水库、池塘等生态系统中,可利用的氮浓度一般处于较低浓度水平,低氮甚至缺氮对水华束丝藻的生长及对氮的利用能力较少报道.了解水华束丝藻在低浓度氮以及无氮条件下的生长特征、异形胞发生格局以及固氮特征,对蓝藻水华的发生机制以及控制可以提供理论和技术指导.本实验以滇池水华束丝藻为研究对象,旨在通过研究滇池水华束丝藻在低浓度硝态氮条件下的生长特征,无氮条件下异形胞的形成过程及固氮能力,以期初步揭示在野外较低的氮浓度条件下,滇池水体中水华束丝藻生长过程中氮的生物学和生态学作用,了解氮在富营养化湖泊滇池水华束丝藻-微囊藻演替过程中的影响,为了解类似水体中固氮和非固氮蓝藻的演替提供新的认识.
1 材料与方法
1.1 藻种分离
样品采自滇池北部海埂湾,其中水华束丝藻Aph1于2010年4月分离,Aph9于2011年3月分离.采样时,用1L有机玻璃采水器采集约30ml表层水,放于50ml样品瓶中,立即带回实验室进行纯种分离.藻种分离时,取少量新鲜湖水,置于灭菌载玻片上,在Olympus CX31型光学显微镜下用灭菌的巴斯德毛细管吸取单根藻丝,用无菌水清洗5次,再用无菌CT培养基[21]清洗3次后置于添加CT培养基的12孔真空包装细胞培养板内.培养条件为:温度20±1℃、光暗比12h∶12h、光强20μE/(m2·s).分离的培养物在1个月左右生长成簇后,取少量培养物镜检是否为水华束丝藻,水华束丝藻藻株转接到含150ml CT培养基的三角瓶扩大培养,培养条件同上.两株藻种经中国科学院淡水藻种库进一步鉴定为水华束丝藻.藻种采集地(24°56′43″N,102°38′55″E)周围的理化参数为:水温15.1±0.36℃、pH 8.92±0.42、溶解氧7.65±0.14mg/L、透明度28.5±2.0cm、叶绿素a浓度 121.2±39.6μg/L、溶解性反应磷浓度0.027±0.009mg/L、总溶解磷浓度0.037±0.010mg/L、总磷浓度0.332±0.056mg/L、总溶解氮浓度2.933±0.178mg/L、总氮浓度4.360±1.594mg/L.
1.2 藻细胞培养与计数
分离的水华束丝藻培养到一定生物量后转接至CT培养基,以对数生长期的水华束丝藻作为实验材料.水华束丝藻藻丝接种前用无氮培养基清洗3次,不进行氮饥饿处理直接作为材料接种于不同浓度的硝态氮培养基中,实验培养条件为:BG-11培养基[22]、温度20±1℃、光暗比为12h∶12h、光强30μE/(m2·s).每天手动摇动藻种两次并更换藻种在培养箱的位置.接种后,每隔两天取样一次,取样时间为9:00-11:00,在岛津1800分光光度计上于680nm处测定吸光度.藻丝以及异形胞数目在Olympus CX31型光学显微镜下直接用浮游植物计数框计数.整个实验过程中,分离和培养的水华束丝藻一直处于漂浮状态.
1.3 理化参数的测定
水体pH、溶解氧及水温的数据均在取样时使用YSI 550A(YSI, USA)现场测定,透明度通过塞氏盘测定,营养盐的测定采用文献[23]的方法.无氮培养基中水华束丝藻固定的氮以测定的总氮表示,其方法同上,所有玻璃器皿使用前于盐酸(1+9)中浸泡48h,然后用无氨水洗净.
1.4 统计学方法
数据绘图在Origin 8.0软件中完成,用SPSS 18.0进行One-way ANOVA 分析,P<0.05为显著相关.
2 结果
2.1 不同硝态氮浓度下水华束丝藻的生长特征
两株水华束丝藻Aph1和Aph9分别接种于添加不同浓度(0、0.5、2、5、10、20mg/L,对照组BG-11含硝态氮约274mg/L)硝态氮的BG-110(不含硝态氮)培养基中. 结果表明(图1):随着培养时间的增加,水华束丝藻在各浓度硝态氮培养基中的生物量均逐渐增加. 两株水华束丝藻对不同浓度硝态氮的响应比较一致,即随着硝态氮浓度的升高,水华束丝藻的生长速率逐渐增加. 水华束丝藻在硝态氮浓度越高的培养基中,其达到稳定期的时间也越短. 水华束丝藻Aph1在生长约40d后达到稳定,680nm处的吸光度为1.354,但是其在硝态氮浓度为20、10、5和2mg/L培养基中的生物量在45 d已经接近BG-11培养基中的生物量水平(P>0.05),0.5mg/L硝态氮中水华束丝藻生物量虽然与对照组差异比较显著(P<0.05),但其生物量也较高,并保持上升的趋势.无氮培养基中,水华束丝藻生长受到一定的抑制,生长相对比较缓慢,但是实验后期生物量也比较高,吸光度接近0.9.相比Aph1,Aph9生长潜力更大,即使培养45d其生物量仍然处于增长的趋势,在不同浓度的硝态氮培养基中,生物量均很高,仅无氮培养基中处理组的生物量与对照组存在显著差异(P<0.05).

图1 不同硝态氮浓度对滇池水华束丝藻生长的影响Fig.1 The effects of different nitrate-nitrogen concentrations on the growth of A.flos-aquae isolated from Lake Dianchi

图2 无氮诱导藻丝异形胞分化特征Fig.2 Heterocyst characteristics of A. flos-aquae induced in N-free status
2.2 无氮诱导异形胞的分化

图3 Aph9不同时期异形胞以及异形胞荧光特征Fig.3 Lightmicroscope ofheterocyst formation at different stages and autofluorescence of A. flos-aquae strain Aph9
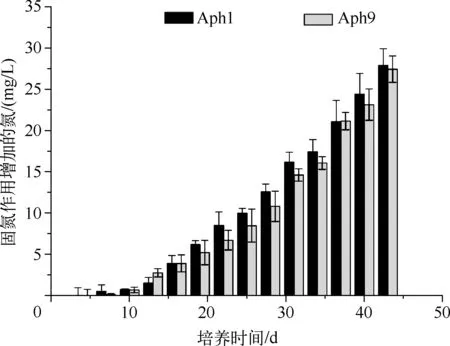
图4 两株水华束丝藻固氮特征Fig.4 Nitrogen fixation characteristics of the two strains of A. flos-aquae
固氮蓝藻能够在缺氮环境下形成异形胞,固定大气中的氮气以适应氮胁迫.对分离的两株水华束丝藻进行无氮培养,诱导其异形胞的分化,发现水华束丝藻能够迅速分化形成异形胞,Aph9经过诱导后,第3d含有异形胞的藻丝即达到50%以上,第7d高达72%,之后维持较高的比例持续波动.Aph1诱导形成异形胞的总体趋势与Aph9非常相似,其形成异形胞的比例也很高,达50%以上.对照组BG-11培养基中初始时期含少量异形胞,可能是初期阶段收集藻丝过程中经历短暂的缺氮过程诱导所致,培养10d以后对照组完全没有异形胞形成(图2).
由图3可见,水华束丝藻细胞能感受氮胁迫状态,缺氮能快速诱导形成异形胞.Aph9接种初期即观察到有部分藻丝形成异形胞,随着培养时间延长,异形胞的比例开始增加,培养后期也有大量的藻丝含有异形胞(图3A~D),但是,对照组BG-11培养的水华束丝藻几乎完全不含异形胞(图3E).Aph9藻丝上形成的异形胞具有明显不同的自发荧光特征,异形胞红色荧光比营养细胞弱,并且在与营养细胞结合的部分无荧光(图3F).
2.3 水华束丝藻的固氮作用
Aph1、Aph9接种于无氮培养基,测得不同培养时间培养基中的总氮,后期测得的总氮去除初始条件接种时期的氮即为水华束丝藻因固氮作用增加的氮.实验结果显示(图4):两株水华束丝藻培养基中氮含量增加趋势比较一致.在开始的1周内培养基中氮含量未见增加,从第7d开始稳步增加.随着培养时间的增加,培养基中因水华束丝藻固氮作用增强,总氮逐步增加.到实验后期,培养基中增加的总氮含量超过25mg/L.
3 讨论
氮是浮游植物生长所需的大量必需元素之一,它存在于所有组成蛋白质的氨基酸中,同时也是合成藻蓝蛋白、叶绿素a以及DNA等核酸的基本元素.藻类在氮饥饿胁迫条件下,藻蓝蛋白做为氮库降解,叶绿素a含量下降,光合作用降低,细胞代谢速率减缓[24],藻类最后可能死亡或者以类似休眠体或孢子的形式长期生存[25-26].蓝藻中具有明显固氮能力的种类,已作过研究或测定过固氮能力的约有160余种和变种,其中绝大多数属于念珠藻目中的种类,如念珠藻、鱼腥藻、束丝藻、单岐藻、眉藻等.固氮蓝藻在有氮的情况下不形成异形胞,只有当培养环境中缺乏可利用的氮源时,才开始分化异形胞.当细胞中缺乏可用的NH3时,导致α-酮戊二酸的大量积累,细胞内自由Ca2+浓度的升高,它们是氮饥饿程度的重要信号,诱导异形胞的分化.异形胞利用N2合成NH3,NH3通过谷氨酰胺合成酶-谷氨酸合成酶循环形成谷氨酰胺和谷氨酸,被转运到营养细胞中[27-29],供细胞生长和分化的需要.
滇池分离的水华束丝藻不经氮饥饿处理直接接种于无氮BG-11培养基,能够诱导藻丝体异形胞的分化,含有异形胞的藻丝比例在第3d即可达到50%以上,表明水华束丝藻能快速应对缺氮胁迫.水华束丝藻在无氮培养基中生长的初期会受到一定程度的抑制,生长速率相比BG-11对照组低,但在后期生长差异减小,当添加的氮浓度达到0.5mg/L以上时,其生物量与BG-11对照组无显著差异.以上结果表明:在富营养化水体中可供利用的氮浓度高于0.5mg/L时,水华束丝藻等固氮蓝藻的生长可能不受氮营养的限制.氮在春季通常被认为是浮游植物生长的限制因子[30],滇池束丝藻水华在春季发生,除了水华束丝藻对低温有较强的适应性外,固氮作用可能在滇池水华束丝藻水华的发生过程中起到很重要的作用.
Schindler等基于实验湖沼学研究提出了富营养化治理控磷是关键的观点[31-34],但是单一的控磷措施在很多湖泊中很可能失败,这些水体中磷在底泥和水体之间循环的速度很快,氮元素对蓝藻水华起着重要作用,如:阿波普卡湖、奥基乔比湖、太湖、东湖以及霞浦湖[30].营养盐添加实验表明,太湖冬、春季出现磷限制,夏、秋季出现氮限制,微囊藻水华维持需要的氮来源于新增加的氮以及先前的氮源[35].水华束丝藻即便在无氮培养基中生长,通过固氮作用增加的氮可达30mg/L,在水华束丝藻水华消退的过程中,释放的大量的氮可能成为其它非固氮蓝藻水华的诱发因子.
滇池存在水华束丝藻-微囊藻的演替,结合野外条件下氮浓度的数据分析和水华束丝藻异形胞的形成特征,将可望揭示氮在滇池水华束丝藻-微囊藻演替过程中的作用,这将有助于阐明两种蓝藻演替中营养盐的驱动机制.国内诸多水体经常出现固氮蓝藻与非固氮蓝藻演替或者共同存在的现象,如洱海存在微囊藻、束丝藻和鱼腥藻;巢湖存在微囊藻与鱼腥藻之间的演替;太湖以微囊藻为主,但是同时也有束丝藻、鱼腥藻等固氮蓝藻存在.水体中的氮处于频繁动态变化中,进一步认识固氮蓝藻对氮的利用方式和机制将有助于揭示其水华的发生机制,并对固氮蓝藻与非固氮蓝藻的竞争及演替提供新认识.
[1] 余博识,吴忠兴,朱梦灵等.水果湖湾蓝藻水华的形成及其对东湖影响的评价.水生生物学报,2008,32(2):286-289.
[2] 吴忠兴,虞功亮,施军琼等.我国淡水水华蓝藻——束丝藻属新记录种.水生生物学报,2009,33(6):1140-1144.
[3] Carmichael WW, Drapeau C, Anderson DM. Harvesting ofAphanizomenonflos-aquaeRalfs ex Born.& Flah.var.flos-aquae(Cyanobacteria) from Klamath Lake forhuman dietary use.JournalofAppliedPhycology, 2000,12: 585-595.
[4] Tsujimura S, Ishikawa K, Tsukada H. Effect of temperature on growth of the cyanobacteriumAphanizomenonflos-aquaein Lake Biwa and Lake Yogo.PhycologicalResearch, 2001,49(4): 275-280.
[5] Rücker J, Stükenb A, Nixdorfa Betal. Concentrations of particulate and dissolved cylindrospermopsin in 21Aphanizomenon-dominated temperate lakes.Toxicon, 2007,50:800-809.
[6] Üveges V, Tapolczai K, Krienitz Letal. Photosynthetic characteristics and physiological plasticity of anAphanizomenonflos-aquae(Cyanobacteria,Nostocaceae) winter bloom in a deep oligo-mesotrophic lake (Lake Stechlin, Germany).Hydrobiologia, 2012,698(1): 263-272.
[7] 林婉莲,刘鑫洲.武汉东湖浮游植物各种成份分析与沉淀物中浮游植物活体碳、氮、磷的测定.水生生物学报,1985,9(4):359-364.
[8] 吴 洁,虞左明.西湖浮游植物的演替及富营养化治理措施的生态效应.中国环境科学,2001,21(6):540-544.
[9] 陈作州,陈 椽,晏 妮等.红枫湖水库浮游植物演变(1980-2006年)和富营养化趋势研究.贵州师范大学学报,2007,25(3):5-10.
[10] 李敦海,刘景元,邢 伟等.四斗朱水库蓝藻水华爆发成因分析及治理对策研究.环境科学与管理,2010,35(2):43-46.
[11] Liu YM, Chen W, Li DHetal. First report of aphantoxins in China-water blooms of toxigenicAphanizomenonflos-aquaein Lake Dianchi.EcotoxicologyandEnvironmentalSafety, 2006,65:84-92.
[12] 张 梅,李 原,王若南.滇池浮游植物的生物多样性调查研究.云南大学学报,2005,27(2):172-176.
[13] Yamamoto Y, Nakahara H. The formation and degradation of cyanobacteriumAphanizomenonflos-aquaeblooms: the importance of pH, water temperature, and day length.Limnology,2005,6:1-6.
[14] Suikkanen S, Kaartokallio H, Hällfors Setal. Life cycle strategies of bloom-forming filamentous cyanobacteria in the Baltic Sea.DeepSeaResearchPartⅡ:TopicalStudiesinOceanography, 2010,57(3/4):199-209.
[15] Yamamoto Y, Nakahara H. Life cycle of cyanobacteriumAphanizomenonflos-aquae.Taiwania, 2009,54(2): 113-117.
[16] Vahtera E, Conley DJ, Gustafsson BGetal. Internal ecosystem feedbacks enhance nitrogen-fixing cyanobacteria blooms and complicatemanagement in the Baltic Sea.AMBIO, 2007,36(2): 186-194.
[17] Conley DJ, Paerl HW, Howarth RWetal. Controlling eutrophication: nitrogen and phosphorus.Science, 2009,323:1014-1015.
[18] Oliver RL, Ganf GG. Freshwater blooms-The ecology of cyanobacteria. Netherlands: Kluwer Academic Publishers, 2000:149-194.
[19] Wood SA, Prentice MJ, Smith Ketal. Low dissolved inorganic nitrogen and increasedheterocyte frequency: precursors toAnabaenaplanktonica blooms in a temperate, eutrophic reservoir.JournalofPlanktonResearch, 2010,32: 1315-1325.
[20] Carey CC, Ibelings BW, Hoffmann EPetal. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate.WaterResearch, 2012,36(5):1394-1407.
[21] Watanabe MM, Ichimura T. Fresh-and salt-water forms ofSpirulinaplatensisin axenic cultures.BullJpnSocPhycol, 1977,25: 371-377.
[22] Stanier RY, Kunisawa R, Mandel Metal. Purification and properties of unicellular blue-green algae (order Chroococcales).BacteriologicalReviews, 1971,35(2): 171.
[23] 国家环境保护局《水和废水监测分析方法》编委会.水和废水监测分析方法:第4版.北京:中国环境科学出版社,2002.
[24] Sauer J, Schreiber U, Schmid Retal. Nitrogen starvation-induced chlorosis inSynechococcusPCC 7942. Low-level photosynthesis as amechanism of long-term survival.PlantPhysiol, 2001,126(1): 233-243.
[25] Brussaard CPD, Noordeloos AAM, Riegman R. Autolysis kinetics of themarine diatomDitylumbrightwellii(Bacillariophyceae) under nitrogen and phosphorus limitation and starvation.JournalofPhycology, 1997,33(6): 980-987.
[26] Peters E, Thomas DN. Prolonged nitrate exhaustion and diatommortality: a comparison of polar and temperateThalassiosiraspecies.JournalofPlanktonResearch, 1996,18(6): 953-968.
[27] Li RH, Watanabe M, Watanabe MM. Akinete formation in planktonicAnabaenaspp.(Cyanobacteria) by treatment with low temperature.JournalofPhycology, 1997,33(4): 576-584.
[28] Muro-Pastor MI, Reyes JC, Florencio FJ. Ammonium assimilation in cyanobacteria.PhotosynthesisResearch, 2005,83: 135-150.
[29] Adams DG. Heterocyst formation in cyanobacteria.CurrentOpinioninMicrobiology, 2000,3: 618-624.
[30] Conley DJ, Paerl HW, Howarth RWetal. Controlling eutrophication: nitrogen and phosphorus.Science, 2009,323(5917): 1014-1015.
[31] Wang HJ, Wang HZ. Mitigation of lake eutrophication: Loosen nitrogen control and focus on phosphorus abatement.ProgressinNaturalScience, 2009,19: 1445-1451.
[32] Carpenter SR. Phosphorus control is critical tomitigating eutrophication.PNAS, 2008,105(32): 11039-11040.
[33] Schindler DW, Hecky RE, Findlay DLetal. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment.PNAS, 2008,105(32): 11254-11258.
[34] Havens KE, Fukushima T, Xie Petal. Nutrient dynamics and the eutrophication of shallow lakes Kasumigaura (Japan), Donghu (PR China), and Okeechobee (USA).EnvironmentalPollution, 2001,111(2): 263-272.
[35] Paerl HW, Xu H, McCarthy MJetal. Controllingharmful cyanobacterial blooms in ahyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P)management strategy.WaterResearch, 2011,45(5): 1973-1983.
