水分胁迫下新西伯利亚银白杨光合能力变化
2014-08-23邱尚志杨青霄沈海龙崔晓涛
王 晶,邱尚志,杨青霄,杨 玲,沈海龙,崔晓涛
(1.东北林业大学 林学院,哈尔滨 150040)
水的短缺始终是制约农林业生产的关键因素,是关系到造林成败的关键[1]。水在经济建设、生态环境治理和解决木材短缺方面均占有重要地位[2-3]。因此,引进和培育抗旱性强的优良杨树品种,对我国干旱、半干旱地区的生态环境改良具有重要意义。
新西伯利亚银白杨(Populusbachofenii×P.pyramidalis‘ROZ’)是适用于北方城市绿化和用材林培育的树种,具有生长迅速、观赏价值高和耐寒(耐-50℃的低温)等特点[4]。俄罗斯新西伯利亚与我国北疆接壤,与我国北方气候差异较小。试种证明俄罗斯新西伯利亚银白杨完全适合在我国东北地区栽培,并具有向我国西北、华北地区延伸的潜力(个人通讯)。目前,新西伯利亚银白杨的组织培养技术、干旱胁迫对新西伯利亚银白杨幼苗生长、叶片保护酶对干旱胁迫响应的研究已有报道[4,5-7]。但对水分胁迫下新西伯利亚银白杨生理响应的研究尚不全面。本研究探讨水分胁迫下新西伯利亚银白杨幼苗叶片的光合能力和渗透调节物质含量的变化,研究结果可为新西伯利亚银白杨耐旱苗木培育过程中合理设置水分锻炼条件提供参考依据。
1 材料和方法
1.1 试验材料
试验材料为新西伯利亚银白杨通过组织培养手段获得的当年生苗木。栽培基质由苗圃熟土和沙以2∶1的体积比例混合而成(含水量约为46%)。当苗木高度达到50 cm时采用控制浇灌水的方法进行处理。参照Hsiao[8]的水分梯度设置方法,设置对照和3种水分梯度处理,即对照(CK)、轻度胁迫(H1)、中度胁迫(H2)、重度胁迫(H3),基质含水量分别为90%~95%土壤田间持水量、60%~65%土壤田间持水量、40%~45%土壤田间持水量和 20%~25%土壤田间持水量。每处理30株苗木。胁迫试验结束后将所有处理均恢复到对照组控水量。
土壤含水量采用美国产TDR土壤水分仪于每天16∶00测定,测定后补足损失的水分到设定的水平。试验期间白天温度25~35℃,夜间温度15~20℃,自然光照(平均光照强度800 μmol/m2·s1)。
1.2 光合能力的测定
于处理开始后第8天(胁迫前期)、第16天(胁迫中期)、第24天(胁迫后期)和干旱胁迫结束后复水处理的第8天,取茎顶端第3~4片完全展开的叶片进行测定。采用Li-Cor 6400便携式光合测定系统测定叶片光合作用相关指标。于晴天8:00~11:00测定。测定时叶温25℃、光照强度800 μmol/m2·s1、CO2浓度350 μmol/mol。可溶性糖含量测定用苯酚法[9],用不同浓度的葡萄糖溶液制作标准曲线。可溶性蛋白质含量测定用考马斯亮蓝法[9]。
1.3 数据处理
实验中各影响因素的作用用单因素方差分析方法评价并进行差异显著性检验。平均数之间统计学上显著差异用邓肯多重比较法进行检验。
2 结果与分析
2.1 水分胁迫对新西伯利亚银白杨光合能力的影响

图1 水分胁迫对新西伯利亚银白杨净光合速率(Pn)的影响
水分胁迫过程中,新西伯利亚银白杨苗木叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)的变化规律如图1~图4所示。方差分析结果表明,胁迫强度对叶片净光合速率、叶片气孔导度、蒸腾速率和胞间CO2浓度的影响均存在显著差异(P<0.05)。随着胁迫程度加深和胁迫时间延长,叶片净光合速率、气孔导度、蒸腾速率、胞间CO2浓度表现为持续下降。胁迫8 d时,H1、H2和H3处理的净光合速率分别较对照下降了8.76%、78.54%和91.70%,气孔导度分别较对照下降了5.72%、42.37%和44.71%,蒸腾速率分别较对照下降了39.46%、62.33%和63.75%,胞间CO2浓度分别较对照下降了28.3%、65%和20.1%。胁迫8 d时,重度胁迫(H3)对叶片净光合速率、叶片气孔导度、蒸腾速率和胞间CO2浓度的影响大于其他处理组。随着胁迫时间延长,中度和重度胁迫处理组的净光合速率、气孔导度、蒸腾速率呈持续下降趋势,至胁迫24 d时,H3处理组苗木叶片的净光合速率降低为-0.7 μmol/m2s1,此时呼吸速率大于光合速率。而H1处理组的净光合速率、气孔导度和蒸腾速率具升高趋势,这表明苗木逐渐适应该土壤水分条件。而胞间CO2浓度变化略有差异,即随着胁迫时间延长,H2处理组胞间CO2浓度降低,而H1、H3处理组的胞间CO2浓度则呈升高趋势。恢复供水后,各处理中以中度胁迫(H2)下的净光合速率、气孔导度、蒸腾速率和胞间CO2浓度的回升幅度最大。

图2 水分胁迫对新西伯利亚银白杨气孔导度(Gs)的影响
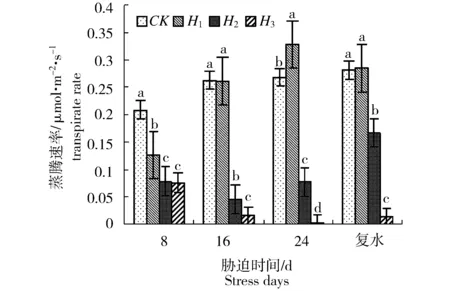
图3 水分胁迫对新西伯利亚银白杨蒸腾速率(Tr)的影响
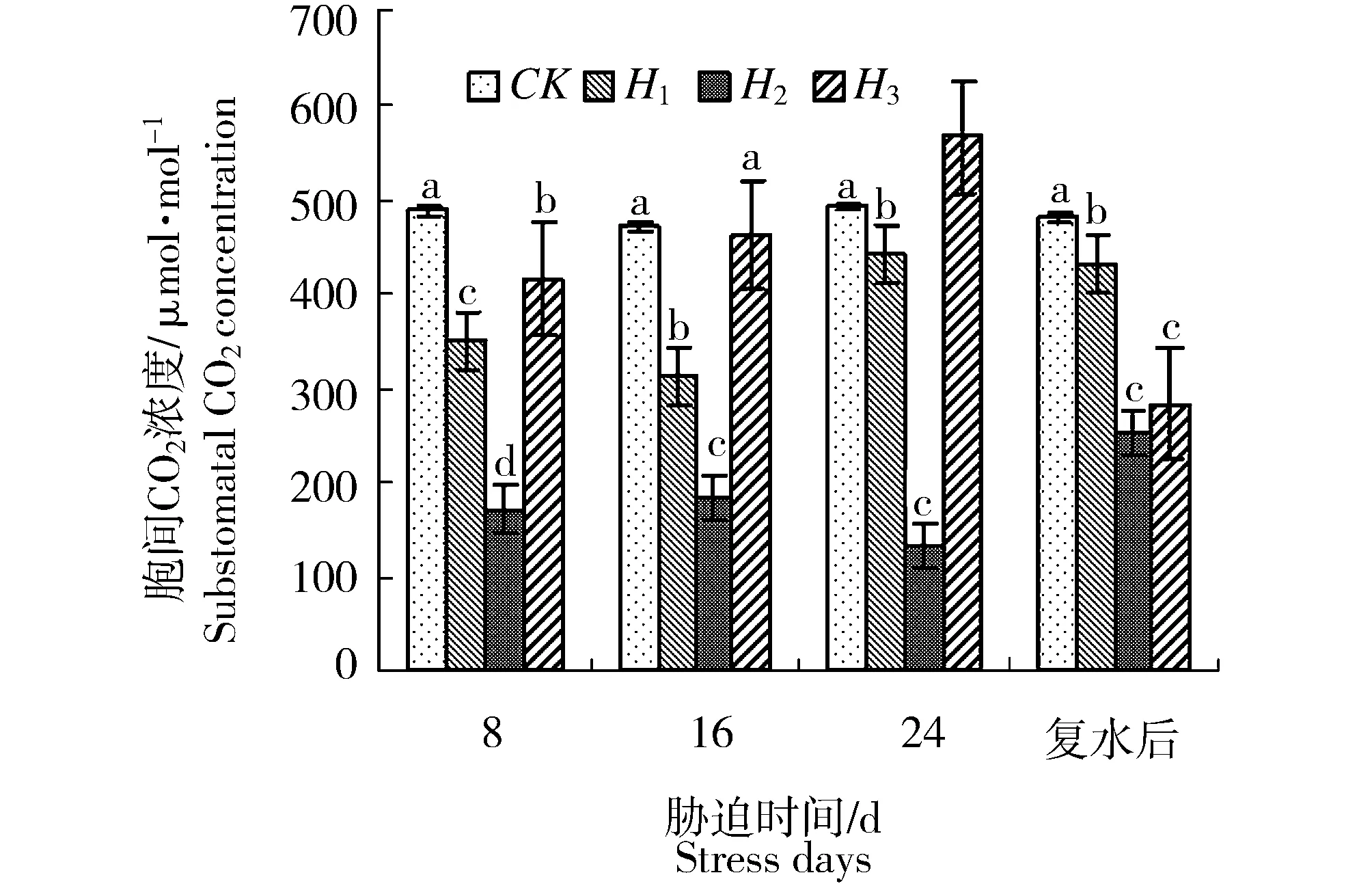
图4 水分胁迫对新西伯利亚银白杨胞间CO2浓度(Ci)的影响
2.2 水分胁迫对渗透调节物质的影响
2.2.1 水分胁迫对叶片可溶性糖含量的影响
水分胁迫对新西伯利亚银白杨叶片可溶性糖含量影响如图5所示。方差分析结果表明,胁迫8 d和16 d时各处理组叶片的可溶性糖含量均显著高于对照(P<0.05),这表明水分胁迫启动了渗透调节机制,提高了叶片中可溶性糖含量,这可以起到脱水保护的作用。但是随着胁迫时间延长,各处理组的可溶性糖含量出现不同程度的减少。恢复供水后叶片可溶性糖含量可恢复至对照水平,但轻度胁迫(H1)的苗木仍具有较高的可溶性糖含量。
2.2.2 水分胁迫对可溶性蛋白质含量的影响
水分胁迫对新西伯利亚银白杨叶片可溶性蛋白质含量的影响如图6所示。方差分析结果表明,胁迫8 d和16 d时各处理组的叶片可溶性蛋白含量均低于对照,但这种差异不显著(P>0.05)。胁迫24 d时,各处理组的可溶性蛋白含量迅速增加,均高于对照。其中H2处理组的可溶性蛋白含量最高,其次是H3处理组,H1处理组的可溶性蛋白质含量与对照相近。推测是随着水分胁迫时间延长,对植物的胁迫伤害加重,胁迫诱导蛋白被合成,总蛋白含量增加,细胞保水力提高。恢复供水后,各处理组可溶性蛋白含量均有不同程度的减小,并趋于对照水平。
2.3 渗透调节物质与叶片光合能力的相关性分析
以胁迫24 d时各指标的平均值作相关性分析(表1),结果表明:
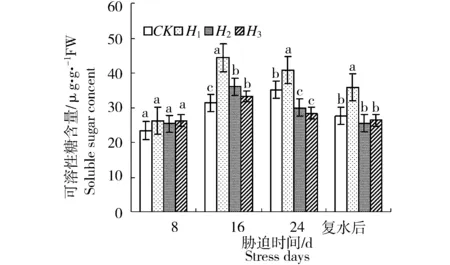
图5 水分胁迫对新西伯利亚银白杨可溶性糖含量的影响
(1)净光合速率(Pn)与气孔导度(Gs)和蒸腾速率(Tr)呈极显著正相关(P﹤0.01),与土壤水分含量呈显著正相关(P﹤0.05)
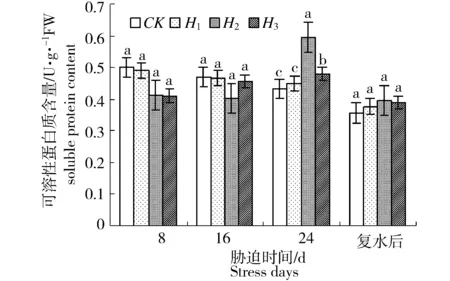
图6 水分胁迫对新西伯利亚银白杨可溶性蛋白含量的影响
(2)气孔导度(Gs)与叶片蒸腾速率(Tr)呈极显著正相关(P﹤0.01),与可溶性糖含量和土壤含水量呈显著正相关(P﹤0.05)。
(3)蒸腾速率(Tr)与可溶性糖含量呈显著正相关(P﹤0.05)。
(4)胞间CO2浓度(Ci)、可溶性蛋白质含量与其他参数以及土壤水分含量间的相关性均不显著(P﹥0.05)。
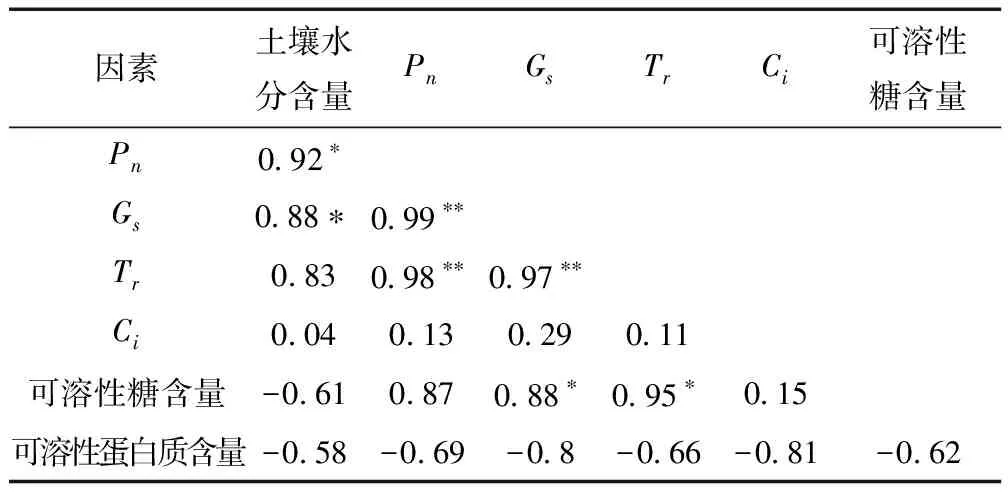
表1 新西伯利亚银白杨的叶片光合能力与渗透调节物质含量的相关分析
注:*表示显著水平(P<0.05),**极显著水平(P<0.01)。* indicste significsncy at 0.05 level while ** at 0.01 level.
3 讨论与结论
3.1 水分胁迫对光合能力的影响
水分胁迫可以抑制新西伯利亚银白杨幼苗生长、影响幼苗形态发育和活性氧代谢[4]。本研究表明,水分胁迫抑制了新西伯利亚银白杨幼苗的光合能力。轻度胁迫(土壤相对含水量60%~65%)对新西伯利亚银白杨幼苗叶片光合能力的影响较小,当恢复供水后,各光合生理指标均有所降低,这表明新西伯利亚银白杨幼苗可以适应轻度水分胁迫(土壤相对含水量60%~65%)。中度胁迫(土壤相对含水量40%~45%)下的叶片净光合速率、气孔导度、蒸腾速率在恢复供水后显著增加,表明中度胁迫(土壤相对含水量40%~45%)对叶片光合机构的损伤可以得到恢复。叶片光合速率的降低主要是气孔部分关闭的结果,恢复供水后可以得到修复[10-12],这与本研究结果一致。严重水分胁迫可使细胞失水,抑制叶绿体中参与碳固定的酶的活性,当叶片相对含水量低于30%时叶绿体会发生不可逆破坏[13]。本研究中,重度胁迫(土壤相对含水量20%~25%)使新西伯利亚银白杨净光合速率大幅下降,恢复供水后仍未得到恢复,说明重度胁迫破坏了光合器官的结构和功能,复水后光合机构很难恢复。
植物叶片净光合速率Pn是衡量植物光合作用能力的重要指标,Ci、Gs和Tr是其主要影响因素[14]。本研究结果表明,叶片净光合速率Pn与气孔导度Gs和蒸腾速率(Tr)呈极显著正相关。气孔导度Gs与叶片蒸腾速率Tr呈极显著正相关。净光合速率Pn与气孔导度Gs均与土壤含水量呈显著正相关关系,即随着土壤含水量下降(胁迫强度增加),叶片净光合速率Pn与气孔导度Gs下降。
逆境胁迫下,植物光合速率降低的原因包括气孔限制和非气孔限制两方面[15-17]。研究结果表明,轻度(土壤相对含水量60%~65%)和中度(土壤相对含水量40%~45%)水分胁迫可以抑制苗木的光合作用,光合下降的原因是气孔导度下降导致的气孔限制。而重度胁迫(土壤相对含水量为20%~25%)下胞间CO2含量变化趋势与净光合速率、气孔导度变化趋势相反,表明重度胁迫下净光合速率下降的主导因子不是气孔限制。研究结果与水分胁迫对云杉光合特性的影响相似[18]。
3.2 水分胁迫对渗透调节物质含量的影响
渗透调节是植物适应水分胁迫的一种重要机制[19,20]。逆境条件影响细胞内的可溶性蛋白质和可溶性糖含量[20-22]。可溶性蛋白质具有较强的亲水胶体性质,可影响细胞的保水力。可溶性糖对细胞膜和原生质胶体有稳定作用[9,21,23-25]。研究结果表明,新西伯利亚银白杨在受到水分胁迫后迅速启动渗透调节机制,利用可溶性糖、可溶性蛋白等渗透调节物质来使细胞渗透势的下降,维持一定的膨压,保证细胞内各种生理生化代谢的正常进行。这个研究结果与陈成升等(2009)对冬小麦的研究结果[26-27]相一致。胁迫过程中,叶片细胞中可溶性蛋白质含量出现了先减小后增加现象,推测是水分胁迫激活了新西伯利亚银白杨体内抗逆基因的表达,影响了苗木叶片中蛋白质的合成,从而使水分胁迫中后期叶片内可溶性蛋白质含量增加。
综上研究结果,认为水分胁迫抑制了新西伯利亚银白杨苗木叶片的光合能力,提高了叶片细胞中可溶性蛋白质含量,并影响了叶片细胞中可溶性糖含量。在土壤相对含水量≥25%条件下叶片维持较好光合作用,气孔因素是导致其净光合速率的下降主要因素。在土壤相对含水量<25%条件下,叶片光合系统受到损伤,非气孔限制是导致净光合速率下降的主要因素。
【参 考 文 献】
[1] 尹春英,李春阳.杨树抗旱性研究进展[J].应用与环境生物学报,2003,9(6):662~668.
[2] 万雪琴,夏新莉,尹伟伦,等.不同杨树无性系扦插苗水分利用效率的差异及其生理机制[J].林业科学,2006,42(5):133-137.
[3] 付士磊,周永斌,何兴元,等.干旱胁迫对杨树光合生理指标的影响[J].应用生态学报,2006,17(11):2016-2019.
[4] 崔晓涛,杨 玲,沈海龙.干旱胁迫对新西伯利亚银白杨抗氧化系统和幼苗生长的影响[J].植物研究,2009,29(6):701-707.
[5] 沈海龙,孔冬梅,王爱芝,等.树木组织培养微枝试管外生根育苗技术[M].北京:中国林业出版社.2009,143-163.
[6] 梁立东,杨 玲,沈海龙,等.新西伯利亚银白杨微枝试管外生根与微环境因子的关系[J].安徽农业科学,2009,37(30):14983~14986,14992.
[7] 梁立东,杨 玲,沈海龙.新西伯利亚银白杨组培不定芽生根质量的影响因子[J].东北林业大学学报,2010,38(1):20-23.
[8] Hsiao T C.Plant responses to water stress[J].Ann Rev Plant Physiology,1973,24:519-570.
[9] 刘祖祺,张石城.植物抗性生理学[M].北京:中国农业出版社,1994.
[10] Calos G M,Lorenzo L.Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress[J].Plant Physiology,2001,126:1196-1200.
[11] 关义新,戴俊英,林 艳.水分胁迫下植物叶片光合的气孔和非气孔限制[J].植物生理学通讯,1995,31(4):293-297.
[12] 邹春静,韩世杰,徐文铎.沙地云杉生态型对干旱胁迫的生理生态响应[J].应用生态学报,2003,14(9):1446-1450.
[13] 蒋明义,杨文英,徐 江,等.渗透胁迫下水稻幼苗中叶绿素降解的活性氧损伤作用[J].植物学报,1994.36(2):142-149.
[14] Berry J A,Downton W J S.Environmental regulation of Photosynthesis[M].NewYork:Academic Press,1982.
[15] Bethke P C,Molcolm C D.Stomatal and nonstomatal compoents to inhibition of photosynthesis in leaves of Capsicum anmam during progressive exposure to NaCl salinity[J].Plant Physiology,1992,99(1):219-226.
[16] 丁友芳,张晓霞,史玲玲,等.葛根净光合速率日变化及其与环境因子的关系[J].北京林业大学学报,2010,32(5):132-137.
[17] 刘会超,孙振元,彭镇华.NaCl胁迫对五叶地锦生长及某些生理特性的影响[J].林业科学,2004,40(6):63-67.
[18] 敖 红,张 羽.水分胁迫对云杉光合特性的影响[J].植物研究,2007,27(4):445-448.
[19] Hellmann H,Funek D,Rentseh D.Hypersensitivity of an Arabidopsis sugar signaling mutant toward exogenous praline application[J].Plant Physiol.,2000,123:779-790.
[20] 王 娟,李德全.逆境条件下渗透调节物质及活性氧清除系统的积累[J].植物学通报.2001,18(4):459-465.
[21] Ludlow M M,Muchhow R C.Acritical evaluation of the traitsfor improving crop yield in water-limited environments[J].Advances in Agronomy,1990,43:107-153.
[22] 赵可夫.植物抗盐生理[M].北京:中国科学技术出版社.1993
[23] 赵黎芳,张金政,张启翔.水分胁迫下扶芳藤幼苗保护酶活性和渗透调节物质的变化[J].植物研究,2003,23(4):437-442.
[24] 肖春旺,周广胜,马风云.施水量变化对毛乌素沙地优势植物形态与生长的影响[J].植物生态学报,2002,26(l):69-76.
[25] 郭卫东,沈 向,李嘉瑞.植物抗旱分子机理[J].西北农业大学学报.1999,27(4):102-108.
[26] 陈成升,谢志霞,刘小京,等.渗盐分、干旱胁迫下冬小麦叶片部分渗透调节物质的动态变化[J].植物研究,2009,29(6):708-713.
[27] 李美兰,李德文,祖元刚.植物体内一氧化氮的来源及其在非生物胁迫中的作用[J].森林工程,2013,29(3):10-12.
