低负荷静态收缩诱发屈肘肌疲劳的肌电与脑电相干性分析
2014-08-07王乐军陆爱云郑樊慧龚铭新
王乐军,陆爱云,郑樊慧,龚铭新,张 磊,董 菲
低负荷静态收缩诱发屈肘肌疲劳的肌电与脑电相干性分析
王乐军1,2,陆爱云2,郑樊慧3,龚铭新1,张 磊1,董 菲1
目的:观察低负荷静态收缩诱发屈肘肌疲劳对肱二头肌sEMG与EEG频率一致性的影响,探索低负荷静态收缩诱发屈肘肌疲劳引起运动皮层与外周肌肉活动的耦合变化情况。方法: 以15名青年男性志愿者为研究对象,记录受试者以20%MVC负荷静态收缩诱发屈肘肌疲劳过程中肱二头肌表面肌电信号(sEMG)与脑电信号(EEG)。截取实验开始最前和最后4.096 s数据,分别记为实验开始阶段和实验结束阶段。对开始阶段和结束阶段所记录的sEMG与脑电Fz、C5、Cz、C6、Pz导联记录EEG进行相干性分析,获取相干函数在alpha(8~12 Hz)、beta(15~30 Hz)、gamma(30~60 Hz)频段内的值。结果: 实验结束阶段内记录sEMG与C5导联EEG相干函数在beta频段内的值、sEMG与Pz导联EEG相干函数在alpha频段内的值具有显著性差异,运动结束阶段记录sEMG与EEG相干函数在相应频段内的值显著高于运动开始阶段。结论:恒定低负荷静态收缩诱发肌肉疲劳使得外周肌肉与对侧运动皮层生物电活动在beta频段内的同步性增加,中枢神经系统对运动皮层与外周肌肉活动进行调节,使得对侧运动皮层与外周肌肉活动之间的耦合性增加,对侧运动皮层与外周肌肉活动有着更为紧密的联系。这可能与疲劳后多种因素引起的皮层调节活动改变有关。
运动性肌肉疲劳;表面肌电信号;脑电信号;相干性分析
1 前言
运动性肌肉疲劳(exercise-induced muscle fatigue),是指运动引起肌肉产生最大收缩力量或最大输出功率暂时性下降的生理现象[10,16,29],其是由于肌肉外周收缩能力(外周因素)下降和中枢对参与收缩肌肉的激活能力(中枢因素)下降共同作用的结果[14,16,29]。在运动性肌肉疲劳发生发展的过程中,由于参与收缩肌肉收缩能力下降和运动神经元、运动皮层神经元的激活能力下降等改变,并通过神经肌肉调控的前馈系统与反馈系统[14,16],引起中枢与外周活动相互联系、相互统一的一系列变化。因大脑是人体随意运动的高级中枢,肌电信号是源于大脑运动皮层控制之下的脊髓运动神经元生物电活动[3,13],而脑电信号是应用脑电图仪记录到的大脑皮层自发或诱发的脑电活动[1,5],因此,疲劳中参与收缩肌肉肌电信号及脑电信号变化之间存在内在的影响和联系。探索运动性肌肉疲劳中肌电信号与脑电信号的关联关系可以为深入揭示外周肌肉与运动皮层之间的联系及更深入地认识疲劳的本质提供极有价值的信息[6,34,37]。
在探索运动性肌肉疲劳的肌电信号与脑电信号关系上,研究者主要采用相干性分析观察不同疲劳条件下肌电信号与脑电信号的频率一致性关系,进而探讨疲劳对肌肉与皮层活动关联的影响[27,31,33,34,37]。相干性分析是描述两分析信号在频域内相似程度的方法,为探索外周肌肉与运动皮层活动的耦合性和相互联系提供了有效的工具。通过综述国内、外有关疲劳相关肌电与脑电相干性分析的研究成果可以发现,目前的研究在疲劳对肌电与脑电相干性分析影响方面仍存在较大争议。其中,Yang Q等[37]、Tuncel D等[33]、Siemionow V等[27]的研究表明,疲劳引起脑电与肌电相干函数主要在beta频段内的值减小;而Ushiyama J等[34]和Tecchio F等[31]的研究则发现,疲劳后大部分受试者脑电-肌电相干函数在beta频段内的值出现显著性的增加。通过对这些研究进行比较分析可以发现,一方面,运动负荷、肌肉收缩方式、测试肌肉等因素都可能会对肌电信号与脑电信号之间的频率一致性关系产生影响[34,35],进而引起上述研究差异性的研究结果;另一方面,目前对疲劳相关肌电、脑电相干性分析的研究仍处于争议与探索的阶段,对运动性肌肉疲劳引起肌电与脑电频率一致性改变问题的认识尚需进一步的研究与探讨。
低负荷静态收缩是人们日常工作和生活中常见的肌肉收缩方式。前期研究表明,低负荷静态收缩诱发的运动性肌肉疲劳在引起中枢与外周变化方面与肌肉在较高负荷收缩所诱发运动性肌肉疲劳存在一定的差异性[17,18,28,30,38],但对低负荷静态收缩诱发运动性肌肉疲劳的研究在前期并未引起众多研究者的关注与重视,特别是在恒定的低负荷收缩致运动性肌肉疲劳引起肌电信号与脑电信号相干性分析变化方面尚鲜见研究报道。
本文以20%MVC(Maximum Voluntary Contraction,MVC)恒定负荷静态收缩诱发屈肘肌疲劳,记录负荷运动实验中屈肘肌肱二头肌sEMG,并同步记录运动皮层脑电信号EEG。通过对比分析疲劳负荷实验开始、结束阶段记录sEMG与EEG的相干性分析结果,探讨恒定低负荷静态收缩诱发屈肘肌疲劳对肱二头肌sEMG与EEG频率一致性的影响,进而探索低负荷静态收缩诱发屈肘肌疲劳引起运动皮层与外周肌肉活动的耦合变化情况。
2 研究对象、方法与步骤
2.1 研究对象
本实验在上海体育学院、同济大学等高校本科生和研究生中随机选取15名健康青年男性志愿者作为受试者(表1)。受试者皆无专业体育运动训练经历,且都是右利手;所有受试者在实验前需接受问卷调查,确保受试者身体健康状况良好;在实验前告知受试者相关的测试方法、程序及注意事项,并签订研究内容知情同意书。

表 1 本研究受试者基本情况一览表Table 1 The General Information of Subjects
2.2 实验程序
2.2.1 实验总体流程
在正式实验开始前先让受试者熟悉实验流程及熟练掌握实验操作方法,并测定受试者屈肘关节最大自主收缩力(MVC,Maximal Voluntary Contraction);待受试者充分休息后,进行正式实验。
正式实验进行之前,受试者坐在座椅上,上身保持直立,分别调节座椅高度和身体姿势,使得髋、膝、踝关节保持90°,双脚平放于地面。右前臂与右上臂保持垂直,右上臂与水平面保持垂直,另一侧手臂自然下垂于体侧。嘱咐受试者身体保持放松,内心保持平静,记录受试者在安静状态自由眨眼20次过程中的眼电和脑电活动,已备后期去除眼电伪迹时用。之后进行屈肘疲劳负荷实验。受试者在保持实验开始前准备姿势的条件下,使受试者右臂腕部拉住可在肘关节处产生20%MVC的重物,至受试者不能坚持负荷运动任务时结束实验[11,12]。记录静态屈肘疲劳负荷过程中受试者肱二头肌表面肌电信号(sEMG)及脑电信号(EEG)。
为减小实验过程中因视觉、眼动、听觉等引起的EEG伪差,实验在光线较暗的实验室进行,并保持实验室的安静。在正式实验前嘱咐受试者放松,使受试者心理处于非常平静的状态。在疲劳负荷实验过程中受试者一直注视着放置于其正前方约3 m处的目标点。
2.2.2 MVC测试
受试者坐在Contrex肌力测试系统(瑞士CONTREX公司,型号PM1/MK2a,b)的座椅上,躯干保持垂直姿势并利用绑带将上身与椅背缚在一起以限制躯干的运动。左侧上肢自然放于体侧,右侧前臂与上臂保持90°,右侧前臂保持与地面平行,两眼平视前方。调节Contrex肌力测试系统上杠杆臂的位置,并固定在右侧腕部位置,在右臂腕部用带子将右前臂绑在杠杆臂上。受试者保持此姿势,对受试者进行3次肘关节屈肌静态自主最大收缩力(MVC)测试,每次测试时间20 s,各次测试的时间间隔为5 min,取最大值作为疲劳运动前最大屈肘关节的MVC。
2.2.3 表面肌电信号采集
记录实验过程中受试者右侧肱二头肌表面肌电信号(sEMG)。放置电极前利用磨砂膏打磨皮肤表面以去除角质和表面污垢,之后用75%酒精棉球清理皮肤表面,以减小阻抗。利用医用胶带将测试电极置于右侧肱二头肌肌腹表面,两电极间距2 cm。通过注射针管往每个肌电测试电极内注入导电膏,并通过导联电阻监测图了解测试导联与皮肤的接触情况。实验过程中保证2个测试肌电电极的电阻低于10 000 Ω,sEMG采样频率设置为2 000 Hz,带通滤波1~1 000 Hz。
2.2.4 脑电信号采集
按照国际10/20系统安放电极。实验时先让受试者带上电极帽,并将具有松紧性的电极帽套罩在电极帽上,以增加脑电电极与头皮的接触。通过注射针管往每个脑电电极内注入导电膏,在此过程中通过NeuroScan Acquire脑电记录系统自带的脑电导联电阻监测图可以确定脑电导联与头皮的电阻值及接触情况,并根据接触情况对某些特定电极进行处理(在电极表面施加压力、增加注入导电膏量等方法)。实验过程中保证每一脑电导联与头皮接触的电阻低于5 000 Ω。选取头顶作为参考电极。脑电信号的采集参数设置为:采样频率2 000 Hz,带通滤波0.01~100 Hz。设置Acquire脑电信号采集系统Amplifiers的Notch参数为50 Hz以屏蔽50 Hz交流电信号对测试脑电信号的影响。
在记录64导联脑电信号的同时,采用NeuroScan脑电信号记录系统自带的双极导联记录右眼垂直眼电信号(VEOG)。在放置电极前采用磨砂膏对皮肤表面进行打磨去除角质及污垢,之后采用医用胶带将2个电极分别固定在左眉上和左眼睑下。眼电信号的记录采样频率为2 000 Hz,带通滤波0.01~100 Hz。
2.3 数据的处理与分析
2.3.1 脑电信号、表面肌电信号预处理
通过对脑电信号、表面肌电信号进行预处理,降低脑电信号与肌电信号中的干扰成分,为后期的数据分析提供数据源,其中,脑电信号的预处理包括脑电预览(去除波形漂移明显的数据部分)、转换参考电极(由头顶参考转换为以双侧乳突参考)、拼接自由眨眼20次记录脑电数据、去除眼电干扰、滤波(FIR滤波器,零相位偏移3~60 Hz带通滤波,滤波强度24 dB/oct)、去伪迹。
肌电信号的预处理主要是滤波处理。采用FIR滤波器对采集的肌电信号进行带通滤波处理,滤波频率5~500 Hz,滤波模式为零相位偏移滤波,滤波强度24 dB/oct。
脑电与肌电信号的预处理工作在NeuroScan Edit 4.3数据分析软件环境下完成。
2.3.2 计算屈肘疲劳实验中肱二头肌sEMG的RMS、MF指标
将疲劳负荷实验中采集的肱二头肌sEMG按等时间间隔(2.048 s)进行分段,计算每段sEMG的均方根振幅RMS、基于傅里叶功率谱转换计算中值频率MF。以每位受试者第1段sEMG所计算指标为初始值,对所计算的RMS和MF值相对于初始值进行标准化[4]。由于每位受试者负荷运动实验的时间不同,为了便于横向计算及比较分析,采用线性插值的方法将前期按等时间间隔计算的各sEMG指标数据标准化为20个数据点[7]。
sEMG的RMS、MF指标计算工作在MATLAB 2009Ra环境下编程实现。
2.3.3 屈肘疲劳实验开始、结束阶段肱二头肌sEMG与EEG功率谱分析
截取实验开始最前和最后4.096 s数据,分别记为实验开始阶段和实验结束阶段数据。分别对实验开始阶段和实验结束阶段记录的肱二头肌sEMG与脑电C5导联记录EEG信号进行傅立叶变换获取分析信号的功率谱曲线,并据此计算sEMG与EEG在alpha(8~12 Hz)、beta(15~30 Hz)、gamma(30~60 Hz)频段内的能量均值。
2.3.4 屈肘疲劳实验开始、结束阶段肱二头肌sEMG与EEG相干性分析
参考相关研究文献[27],分别对上述选取的实验开始阶段和实验结束阶段记录的肱二头肌sEMG与脑电Fz、C5、Cz、C6、Pz导联记录EEG信号进行相干性分析,获取相干函数在alpha(8~12 Hz)、beta(15~30 Hz)、gamma(30~60 Hz)频段内的值等指标。在进行相关性分析的具体计算上,参照Terry等[32]的研究,分段长度(segment length)设置为2 048个样本点,重叠(overlap)设置为50%,短时傅里叶变换窗口类型(segment taper)设置为Hanning window。
sEMG与EEG的相干性分析计算工作在MATLAB 2009Ra环境下编程实现。
2.4 数据的统计分析
选用KS检验(Kolmogorov-Smirnov test)对各分析数据进行正态性检验。根据分析数据的分布特征选用适宜的统计推断方法。静态屈肘运动过程中不同时间阶段内相关指标的差异性采用多个相关样本非参数检验的Friedman方法进行检验;屈肘疲劳实验开始、结束阶段sEMG、EEG的功率谱能量的差异性采用两个相关样本非参数检验的Wilcoxon方法进行检验;疲劳负荷实验开始、结束阶段sEMG与EEG相干性分析结果的差异性检验采用配对t检验进行,差异的显著性水平为P<0.05。数据的统计分析工作在SPSS 13.0软件环境下完成。
3 研究结果
3.1 屈肘疲劳负荷实验中肱二头肌sEMG指标变化特征
静态屈肘疲劳负荷实验过程中肱二头肌sEMG指标随运动时间的变化曲线如图1所示。

图 1 本研究静态屈肘疲劳负荷实验过程中肱二头肌sEMG指标变化曲线图
通过对各受试者疲劳负荷实验运动持续时间进行统计发现,各受试者运动持续时间为364.93±93.92 s,其中,运动持续时间的最大值为561.00 s,最小值为249.00 s。
随着静态屈肘运动持续时间的延长,肱二头肌sEMG指标RMS表现出逐渐增大的趋势(P<0.05),而MF表现出逐渐减小的趋势(P<0.05,图1)。
3.2 屈肘疲劳实验开始、结束阶段肱二头肌sEMG与EEG功率谱特征
屈肘疲劳实验开始、结束阶段记录肱二头肌原始肌电图、脑电图及相关功率谱如图2所示。疲劳负荷实验开始、结束阶段肱二头肌sEMG与EEG能量在不同频段内的值如图3所示。

图 2 本研究疲劳负荷实验开始、结束阶段记录原始肌电图、脑电图及相关功率谱示例图

图 3 本研究疲劳负荷实验开始、结束阶段肱二头肌sEMG与EEG功率谱在不同频段内的均值示意图
疲劳结束阶段记录的肱二头肌原始肌电图和脑电图在时域的振幅值较运动开始阶段有所增加。此外,从肌电信号与脑电信号频谱图可以看出,在大部分频率范围内,实验结束阶段记录的sEMG与EEG功率谱曲线值要高于实验开始阶段(图2)。
运动开始、结束阶段内记录的EEG功率谱在alpha、beta、gamma频段内的均值分别在3 000 μV2、2 000 μV2和1 000 μV2左右,而开始、结束阶段内记录的sEMG功率谱在alpha、beta、gamma频段内的均值则分别在20 000 μV2、40 000 μV2和40 000 μV2左右。运动开始、结束阶段所记录的EEG与sEMG功率谱在alpha、beta、gamma频段内的值皆有显著性差异,运动结束阶段内记录的EEG与sEMG功率谱值要显著高于运动开始阶段(图3)。
3.3 屈肘疲劳实验开始、结束阶段肱二头肌sEMG与EEG相干性分析
从图4开始、结束阶段肱二头肌sEMG与脑电C5导联记录EEG相干函数均值变化曲线可以看出,在运动结束阶段记录信号相干函数的值在大部分频段范围内的值都要比运动开始阶段记录信号相干函数值要高。从图5疲劳负荷实验开始、结束阶段肱二头肌sEMG与脑电不同导联记录EEG相干函数在各频段内的值可以看出,疲劳负荷实验结束阶段sEMG与脑电C5导联记录EEG相干函数在beta频段内的值、sEMG与脑电Pz导联记录EEG相干函数在alpha频段内的值具有显著性差异,实验结束阶段内相干函数值要显著高于实验开始阶段。

图 4 本研究开始、结束阶段肱二头肌sEMG与脑电C5导联记录EEG相干函数变化曲线图
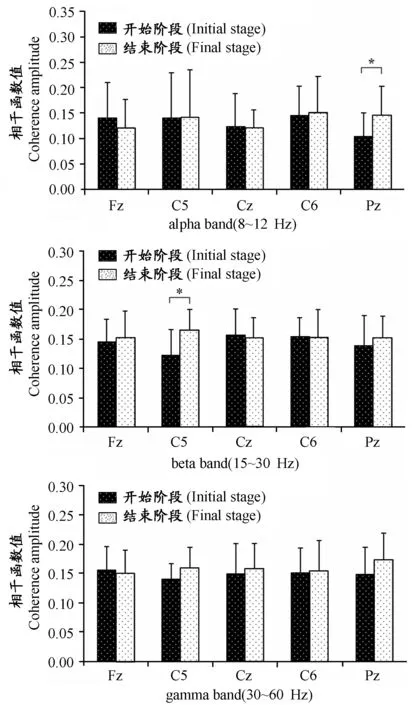
图 5 本研究疲劳负荷实验开始、结束阶段肱二头肌sEMG与脑电不同导联记录EEG相干函数在各频段内的值示意图
4 讨论
本研究的主要研究目的是观察恒定低负荷静态收缩诱发屈肘肌疲劳对肌肉sEMG与EEG频率一致性的影响,探索恒定低负荷静态收缩诱发疲劳引起运动皮层与外周肌肉活动的耦合变化情况。基于此目的,本研究以20%MVC恒定负荷静态收缩方式诱发屈肘肌疲劳,记录此过程中主要收缩肌肉肱二头肌的sEMG,并同步记录大脑皮层电活动EEG。为探讨疲劳对分析指标的影响作用,本研究分别选取每位受试者实验数据的最前4.096 s(开始阶段)和最后4.096 s(结束阶段)数据作为分析数据,以此分别作为受试者非疲劳和疲劳两种生理状态下的测试数据。通过对比分析开始阶段和结束阶段sEMG、EEG的功率谱特征和相干性分析结果,探讨恒定低负荷静态收缩诱发肌肉疲劳对外周肌肉与运动皮层活动耦合性的影响作用。
运动性肌肉疲劳,是指运动引起肌肉产生最大收缩力量或者最大输出功率暂时性下降的生理现象,是肌肉在收缩过程中由于代谢产物堆积、氧气和营养物质供应不足等引起局部肌肉代谢变化、形态结构改变及神经系统功能状况改变等一系列变化的一个具有连续性、动态性的复杂过程[10]。因此,“肌肉产生最大收缩力量或者最大输出功率暂时性下降”是大部分负荷运动任务诱发运动性肌肉疲劳的典型特征,并被广泛应用到运动性肌肉疲劳的定量评价之中[10]。
由于本研究的主要研究目的是探讨疲劳对肌电与脑电相干性分析结果的影响作用,根据实验设计的对照原则,研究只需要对两种不同程度疲劳状态下的实验结果进行比较即可鉴别出疲劳因素的实验效应。因此,对每位受试者,本研究并不需要对疲劳的程度作出定量的评价。参考相关研究文献[19,33,34,37],本研究未测定疲劳实验前后收缩肌肉MVC值的变化或受试者主观疲劳感觉的变化情况,而是通过对受试者实验开始阶段(未疲劳状态)和实验结束阶段(疲劳状态)两种条件下实验结果的差异性比较来了解疲劳对相关研究结果的影响作用。在实验中,受试者运动持续时间为364.93±93.92 s,其中,运动持续时间的最大值为561.00 s,最小值为249.00 s,这与前期近似负荷静态收缩诱发屈肘肌疲劳实验的运动持续时间结果较为相近[18,19]。提示,受试者在实验结束时测试肌肉已达到了疲劳状态。此外,从实验过程中测试肌肉肱二头肌sEMG的变化情况看,时域指标RMS随运动持续时间表现出良好的增加趋势,频域指标MF随运动持续时间表现出逐渐减小的趋势。前期对次最大负荷静态收缩诱发运动性肌肉疲劳引起sEMG指标变化的相关研究表明,RMS随运动持续时间逐渐增加、频域指标MF随运动持续时间逐渐减小,是肌肉在次最大负荷静态收缩诱发局部肌肉疲劳的典型特征[12]。本研究中,RMS、MF随运动持续时间的变化情况一方面反映了疲劳负荷实验过程中神经肌肉系统功能状况的变化;另一方面,也提示本实验疲劳模型设计的科学性和所获实验数据的有效性。
在运动性肌肉疲劳引起参与收缩肌肉sEMG与EEG功率谱能量变化的研究方面,Abdul-latif等[8]的研究表明,在拇收肌进行静态最大自主收缩诱发疲劳后,脑对侧区导联记录EEG在alpha、beta和gamma频段内的均方根振幅RMS都出现增加,其中,RMS在beta频段内的值最大,在gamma频段内的值最小。Yang Q等[37]的研究表明,30%MVC静态屈肘诱发屈肘肌疲劳运动后半段内测试肌肉sEMG及EEG功率谱能量皆出现显著性的增加,其中,EEG能量在beta频段内的增加尤为显著。本研究中,疲劳引起测试肌肉sEMG与EEG能量在各频段皆出现显著性的增加,这与Yang Q等[37]的研究结果也是一致的。提示,由于运动性肌肉疲劳中运动皮层、脊髓前角运动神经元激活能力下降及肌肉外周收缩能力下降,为维持既定的负荷运动任务和补偿由中枢与外周的一系列变化,中枢神经系统募集更多的皮层脊髓细胞和运动单位参与肌肉收缩[20,39]。
两信号的相干性分析结果反映了两分析源信号在不同频率或频段内的接近程度。已有相关研究在相干性分析方法应用于探索运动皮层与外周肌肉间的耦合作用关系方面做出了成功的尝试。Yang Q等[37]记录了受试者以30%MVC静态屈肘诱发肌肉疲劳直至力竭过程中受试者脑电信号及测试侧肱二头肌、肱桡肌、肱三头肌肌电信号,通过对比运动前、后半段记录左、右、中、前、后脑区脑电信号与肌电信号相干函数在beta频段内的值,发现实验后半段的值较实验前半段显著性降低。Tuncel D等[33]采用小波相干性分析方法对疲劳负荷实验不同疲劳阶段内记录的C3、C4导联脑电和肌电进行相干性分析,发现疲劳引起脑电与肌电相干函数在alpha和beta频段内的值减小。Siemionow V等[27]记录了受试者200次最大力量握物体诱发肌肉疲劳过程中的肌电和脑电信号并进行相干性分析,发现疲劳引起肌电和脑电信号相干函数在beta频段和gamma频段内的值减小。而Ushiyama J等[34]对比分析了胫骨前肌在50%MVC静态负荷收缩至力竭前、后分别以30%MVC静态负荷收缩过程中的肌电和Cz导联脑电,结果发现,疲劳后7位受试者中有6位受试者脑电-肌电相干函数在beta频段内的值出现显著性的增加。而从本研究的研究结果看,20%MVC恒定负荷静态收缩诱发屈肘肌疲劳主要引起肱二头肌sEMG与脑电C5导联记录EEG相干函数在beta频段内的值、sEMG与脑电Pz导联记录EEG相干函数在alpha频段内的值出现显著性的增加,实验开始、结束阶段内记录肱二头肌sEMG与Fz、Cz、C6导联记录脑电信号之间的相干函数在各频段内的值无显著性差异。
在肌电与脑电相干性分析结果的影响因素方面,相关研究表明,运动负荷、肌肉收缩方式、测试肌肉等原因都会对肌肉肌电信号与脑电信号直接的频率一致性关系产生影响[34,35]。相比较于其他疲劳负荷实验条件,本研究中受试者进行的负荷运动方式为恒定负荷静态收缩,受试者无需根据特定仪器反馈数据动态调整输出力量的大小。此外,由于本实验所采用的静态收缩负荷为20%MVC,比前期研究所采用的负荷要低。根据Karen S等[18]和Smith J L等[28]的研究,20%MVC负荷静态屈肘诱发的屈肘肌疲劳与肌肉在较高负荷静态收缩或动态收缩所诱发运动性肌肉疲劳的疲劳产生机理存在一定的差异性。因此,本研究与前期研究结果的差异性或许跟恒定低负荷收缩诱发肌肉疲劳机理有别于相关研究所采用的负荷和收缩方式有关,但对于其中的具体机制及中枢调控策略等问题尚需进一步的研究。
前期研究表明,通过观察肌电信号与脑电信号在不同频段内相干函数值的大小可以反映运动皮层与外周肌肉耦合作用的相关信息[25,27,33,34,37],其中,15~30 Hz的beta频段被认为是研究中枢神经系统运动控制的重要频段,相干函数值在该频段内的活动是与大脑运动皮层驱动相关的[11,26],在较大负荷的静态收缩和动态收缩条件下,肌电-脑电在gamma频带内的相干性情况是运动皮层驱动的[19,25]。而alpha频段内的相干函数值似乎受到中枢支配及外周环境改变等多种因素的影响[24]。因此,本研究结果表明,以20%MVC恒定负荷静态收缩诱发屈肘肌疲劳引起肱二头肌sEMG与脑电C5导联记录脑电在beta频段、sEMG与脑电Pz导联记录EEG相干函数在alpha频段内频率一致性的增加。由于运动皮层对躯体运动的对侧性支配原理[2],在受试者右侧肘关节屈曲运动过程中,左脑区作为支配该运动的最主要皮层区域,在其表面记录到的EEG可以反映出皮层对运动控制的重要信息;而C5导联作为左脑区区域的导联,其记录EEG与肱二头肌sEMG相干函数在beta频段内的值增加,提示,疲劳引起外周收缩肌肉与对侧脑区运动皮层活动的振荡耦合性增加。
对于运动皮层与外周肌肉生物电活动频率一致性产生的生理学机制,最初的研究认为其与皮质脊髓轴突及其发出抵达脊髓运动神经元的单突触连接调节活动密切相关[11,34]。此外,有研究认为,肌电与脑电相干性分析结果可能跟同步放电的运动皮层神经元数目有关[23]。最近的研究则表明,肌电与脑电相干性分析结果是一个受到多种因素影响的复杂问题,其中,外周肌肉的感觉反馈作用[9]及伦肖细胞抑制作用的变化[36]等都会对运动皮层与外周肌肉生物电活动频率一致性产生影响。本研究中发现的运动性肌肉疲劳引起肌电与脑电相干函数在beta频段内的增加可能跟疲劳后各种原因引起上运动神经元对脊髓内α运动神经元支配性的改变[11,15]、疲劳后参与活动的运动皮层细胞活动的同步性增加[23]、Ia类传入纤维反馈调节作用变化[21]、疲劳引起的伦肖细胞抑制作用降低[22]等因素共同作用进而引起皮层调节活动的改变有关。因此,通过以上讨论可以看出,运动性肌肉疲劳诱发肌电与脑电信号相干函数在beta频段内值的变化是疲劳后中枢与外周特定性改变引起的必然外在反映。这种变化一方面揭示了疲劳引起中枢与外周耦合性的总体变化;另一方面,也为深入认识和探索疲劳中运动皮层、脊髓、外周肌肉、神经传导通路及相关调控系统之间功能活动的联系提供了参考依据。
5 结论
恒定低负荷静态收缩诱发肌肉疲劳使得外周肌肉与对侧运动皮层生物电活动在beta频段内的同步性增加,中枢神经系统对运动皮层与外周肌肉活动进行调节,使得对侧运动皮层与外周肌肉活动之间的耦合性增加,运动皮层与外周肌肉活动有着更为紧密的联系,这可能跟疲劳后多种因素引起的皮层调节活动改变有关。
[1]鲍九枝,郭磊,侯宽.运动疲劳后恢复期脑电变化的探讨[J].吉林医药学院学报,2007,28(1):17-19.
[2]潘珊珊.运动解剖学(第1版)[M].北京:人民体育出版社,2007.
[3]王健,方红光,杨红春.运动性肌肉疲劳的表面肌电非线性信号特征[J].体育科学,2005,25(5):39-43.
[4]王乐军,黄勇,龚铭新,等.运动性肌肉疲劳过程中主动肌与拮抗肌sEMG相干性分析[J].体育科学,2011,31(10):79-84.
[5]尧得中.脑功能探测的电学理论与方法[M].北京:科学出版社,2003.
[6]张立,周彬.不同形式肌肉运动疲劳前后肌电参数及脑电α指数的变化特征[J].武汉体育学院学报,2010,44(4):65-69.
[7]A W D.Biomechanics and Motor Control of Human Movement[M].New York:John Wiley Sons,2009.
[8]ABDUL-LATIF A,COSIC I,KUMAR D,etal.Power changes of EEG signals associated with muscle fatigue:The root mean square analysis of EEG bands:Intelligent Sensors,Sensors Networks and Information Processing Conference[Z].Melbourne,2004.
[9]BAKER S N,CHIU M,FETZ E E.Afferent encoding of central oscillations in the monkey arm[J].J Neurophysiol,2006,95(6):3904-3910.
[10]CIFREK M,MEDVED V,TONKOVIC S,etal.Surface EMG based muscle fatigue evaluation in biomechanics[J].Clin Biomech (Bristol,Avon),2009,24(4):327-340.
[11]CONWAY B,HALLIDAY D,FARMER S.Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man[J].J Physiol,1995,489(3):917-924.
[12]DE LUCA C J.The use of surface electromyography in biomechanics[J].J Appl Biomech,1997,13(2):135-163.
[13]DUCHENE J,HOGREL J Y.A model of EMG generation[J].IEEE Trans Biomed Eng,2000,47(2):192-201.
[14]ENOKA R M,BAUDRY S,RUDROFF T,etal.Unraveling the neurophysiology of muscle fatigue[J].J Electromyography Kinesiol,2011,21(2):208-219.
[15]FARMER S F,BREMNER F D,HALLIDAY D M,etal.The frequency content of common synaptic inputs to motoneurones studied during voluntary isometric contraction in man[J].J Physiol,1993,470:127-155.
[16]GANDEVIA S C.Spinal and supraspinal factors in human muscle fatigue[J].Physiol Rev,2001,81(4):1725-1789.
[17]HUNTER S K,BUTLER J E,TODD G,etal.Supraspinal fatigue does not explain the sex difference in muscle fatigue of maximal contractions[J].J Appl Physiol,2006,101(4):1036-1044.
[18]KAREN S,SIMON C G,GABRIELLE T,etal.The effect of sustained low-intensity contractions on supraspinal fatigue in human elbow flexor muscles[J].J Physiol,2006,573(2):511-523.
[19]KATTLA S,LOWERY M M.Fatigue related changes in electromyographic coherence between synergistic hand muscles[J].Exp Brain Res,2010,202(1):89-99.
[20]LIU J Z,SHAN Z Y,ZHANG L D,etal.Human brain activation during sustained and intermittentsubmaximalfatiguemusclecontractions:anFMRI study[J].J Neurophysiol,2003,90:300-312.
[21]LOSCHER W N,CRESSWELL A G,THORSTENSSON A.Excitatory drive to the alpha-motoneuron pool during a fatiguing submaximal contraction in man[J].J Physiol,1996,491( Pt 1):271-280.
[22]LOSCHER W N,CRESSWELL A G,THORSTENSSON A.Electromyographic responses of the human triceps surae and force tremor during sustained submaximal isometric plantar flexion[J].Acta Physiol Scand,1994,152(1):73-82.
[23]LUNDBYE-JENSEN J,NIELSEN J B.Central nervous adaptations following 1 wk of wrist and hand immobilization[J].J Appl Physiol,2008,105(1):139-151.
[24]MCAULEY J H,MARSDEN C D.Physiological and pathological tremors and rhythmic central motor control[J].Brain,2000,123(8):1545-1567.
[25]OMLOR W,PATINO L,HEPP-REYMOND M C,etal.Gamma-range corticomuscular coherence during dynamic force output[J].Neuroimage,2007,34(3):1191-1198.
[26]SALENIUS S,PORTIN K,KAJOLA M,etal.Cortical control of human motoneuron firing during isometric contraction[J].J Neurophysiol,1997,77:3401-3405.
[27]SIEMIONOW V,SAHGAL V,YUE G H.Single-Trial EEG-EMG coherence analysis reveals muscle fatigue-related progressive alterations in corticomuscular coupling[J].IEEE Trans Neural Syst Rehabil Eng,2010,18(2):97-106.
[28]SMITH J L,MARTIN P G,GANDEVIA S C,etal.Sustained contraction at very low forces produces prominent supraspinal fatigue in human elbow flexor muscles[J].J Appl Physiol,2007,103:560-568.
[29]TAYLOR J L,BUTLER J E,GANDEVIA S C.Changes in muscle afferents,motoneurons and motor drive during muscle fatigue[J].Eur J Applied Physiol,2000,83(2-3):106-115.
[30]TAYLOR J L,GANDEVIA S C.A comparison of central aspects of fatigue in submaximal and maximal voluntary contractions[J].J Appl Physiol,2008,104(2):542-550.
[31]TECCHIO F,PORCARO C,ZAPPASODI F,etal.Cortical short-term fatigue effects assessed via rhythmic brain-muscle coherence[J].Exp Brain Res,2006,174(1):144-151.
[32]TERRY K,GRIFFIN L.How computational technique and spike train properties affect coherence detection[J].J Neurosci Methods,2008,168:212-223.
[33]TUNCEL D,DIZIBUYUK A,KIYMIK M K.Time frequency based coherence analysis between EEG and EMG activities in fatigue duration[J].J Med Syst,2010,34(2):131-138.
[34]USHIYAMA J,KATSU M,MASAKADO Y,etal.Muscle fatigue-induced enhancement of corticomuscular coherence following sustained submaximal isometric contraction of the tibialis anterior muscle[J].J Appl Physiol,2011,110(5):1233-1240.
[35]USHIYAMA J,TAKAHASHI Y,USHIBA J.Muscle dependency of corticomuscular coherence in upper and lower limb muscles and training-related alterations in ballet dancers and weightlifters[J].J Appl Physiol.2010,109(4):1086-1095.
[36]WILLIAMS E R,BAKER S N.Renshaw cell recurrent inhibition improves physiological tremor by reducing corticomuscular coupling at 10 Hz[J].J Neurosci,2009,29(20):6616-6624.
[37]YANG Q,FANG Y,SUN C K,etal.Weakening of functional corticomuscular coupling during muscle fatigue[J].Brain Res,2009,1250:101-112.
[38]YASSIERLI,NUSSBAUM M A.Utility of traditional and alternative EMG-based measures of fatigue during low-moderate level isometric efforts[J].J Electromyography Kinesiol,2008,18(1):44-53.
[39]YUE G H,BILODEAU M,HARDY P A,etal.Task-dependent effect of limb immobilization on the fatigability of the elbow flexor muscles in humans[J].Exp Physiol,1997,82:567-592.
CoherenceAnalysisbetweenEEGandEMGinLow-levelIsometricElbowFlexorMuscleFatigueContraction
WANG Le-jun1,2,LU Ai-yun2,ZHENG Fan-hui3,GONG Ming-xin1,ZHANG Lei1,DONG Fei1
Objective:The aim of this study was to examine coherence between EEG and EMG in elbow flexor muscle fatigue duration induced by low-level isometric contraction and thus to explore the coupling of muscle and cortex during low-level isometric elbow flexor contraction induced muscle fatigue.Methods:Fifteen healthy male volunteers sustained isometric elbow flexion at 20% maximal level until exhaustion while their brain (EEG) and biceps brachii muscle (sEMG) activities were recorded.The initial and final 4.096 second data were chosen and coherence was observed between sEMG of biceps brachii muscle and EEG recorded from Fz,C5,Cz,C6,Pz electrodes in alpha (8~12 Hz),beta (15~30 Hz) and gamma (30~60 Hz) bands.Results:The power of both EEG and EMG increased significantly in alpha,beta,gamma frequency bands while coherence in beta band between sEMG and EEG of C5 and coherence in alpha band between sEMG and EEG of Pz increased significantly in final stage compared with initial stage.Conclusions:The coupling between contralateral sensorimotor cortex and muscle activities was enhanced as a result of constant low-level isometric fatiguing contraction.This may be related to changes of cortical regulation activities relevant to exercise-induced muscle fatigue.
exercise-inducedmusclefatigue;sEMG;EEG;coherenceanalysis
1000-677X(2014)02-0040-08
2013-08-24;
:2014-01-03
中央高校基本科研业务费专项资金资助(1430219025);上海体育学院研究生教育创新基金资助(YJSCX201107)。
王乐军(1982-),男,山东临沂人,实验师,博士,研究方向为运动生物力学,Tel:(021)65981711,E-mail:wlj0523@163.com;陆爱云(1943-),女,江苏苏州人,教授,博士研究生导师,研究方向为运动能力的提高和运动损伤机理的探讨。
1.同济大学 体育部 运动与健康研究中心,上海 200092;2.上海体育学院 运动科学学院,上海 200438; 3.上海体育科学研究所,上海 200030 1.Sport and Health Research Center,Physical Education Department,Tongji University,Shanghai 200092;2.School of Kinesiology,Shanghai University of Sport,Shanghai 200438;3.Shanghai Research Institute of Sports Science,Shanghai 200030,China.
G804.6
:A
