小兴安岭红松日径向变化及其对气象因子的响应
2014-08-04李兴欢刘瑞鹏毛子军刘林馨
李兴欢,刘瑞鹏,毛子军,*,宋 媛,刘林馨,孙 涛
(1.东北林业大学森林植物生态学教育部重点实验室,哈尔滨150040;2.齐齐哈尔大学生命科学与农林学院,齐齐哈尔161006)
树干径向变化记录仪(Dendrometer)因为能够连续和自动测量茎干的径向变化,而逐渐被广泛应用[1-5]。这种仪器不仅可以测量径向昼夜变化和树木季节性生长动态,也可以与同步观测的环境因子相结合来揭示树木径向生长对环境的响应。气候因子虽然不能直接反映树木生物学的生长,但是与同步的树木径向增量相结合,为年内径向生长分析提供了可能[2]。近年来,越来越多的人开始研究径向生长与气候的关系。Downes等[1]监测了澳大利亚的亮果桉(Eucalyptus nitens)和蓝桉(E.globulus)的径向变化,研究了灌溉条件下气候与生长的关系,结果表明:树木生长与春季温度呈正相关,夏季呈弱相关或负相关。Deslauriers等[2]使用周期循环法研究了加拿大魁北克北部的香脂冷杉(Balsam fir),结果表明树木生长与降雨呈正相关,与膨胀阶段的水汽压亏缺呈负相关,膨胀和增量阶段的气候对径向生长的影响更大。我国对树木径向生长的研究起步较晚,且非常有限。管伟[6],熊伟[7]分别研究了六盘山北侧和南坡的华北落叶松(Larix principis-rupprechtii)的径向生长,认为温度和降雨是影响树木径向生长的主要因子。董满宇[8-9]对芦芽山林线华北落叶松和白杄(Picea meyeri)茎干生长进行了研究,发现土壤温度是茎干快速生长时段的主导环境因子。我国东北寒冷地区的相关研究尚未见报道。
阔叶红松林是较为寒冷的北温带典型的地带性植被,红松作为中国东北地区阔叶红松林的建群种和优势种,具有重要的生态学价值和经济价值。目前对红松径向生长的研究主要是通过年轮分析年轮宽度与气候的关系[10-11]。陈力等[10]通过年轮分析研究了长白山红松生长量与温度的关系,认为生长季气温是影响红松生长最重要的气候因子。王晓明等[11]研究了长白山北坡不同年龄红松的年表特征及其与气候因子间的关系,结果表明,年龄因素对红松年表的气候响应方面存在一定影响,且高龄年表对气候响应的敏感性更高;高龄红松径向生长与当年多个月份的月最高温度正相关,与当年4月、9月的月平均最低温度负相关,同时受到上年5月及当年5月总降水量的影响。然而,在昼夜和一个生长季的时间尺度上红松树干的径向生长动态如何,生长期开始时间的确定,以及影响日径向变化的关键气候因子都是尚待揭示的问题。本研究用树木径向生长记录仪连续监测了4月至11月的红松径向变化,主要目的是:(1)确定红松径向生长开始时间、生长期以及生长期内红松径向生长动态;(2)分析红松日径向变化与气候因子的关系。
1 研究区概况
1.1 研究地区概况
研究地位于小兴安岭南坡达里带岭支脉东坡的凉水国家级自然保护区(47°10'19″N,128°52'96″E),隶属于黑龙省伊春市带岭区。凉水国家级自然保护区分布着红松为主的寒温带针阔混交林,全区海拔高度在208—707 m,地带性土壤为暗棕壤。该地区属温带大陆性季风气候,夏短冬长。年平均气温-0.3℃,年平均降水量676 mm,6—8月降雨占全年降水量的60%以上(图1);无霜期100—120 d,积雪期130—150 d。

图1 凉水自然保护区2012年与1974—2008年的月平均温度与月平均降雨量Fig.1 Meanmonthlytemperatureandprecipitationat Liangshui National Nature Reserve in 2012 and 1974—2008
2 研究材料与方法
2.1 研究地概况
在保护区选择生长状况良好的天然阔叶红松林,红松年龄约为200—250年,建立一个30 m×20 m的标准样地。海拔359 m,坡向南,坡度10°。建群种和优势树种为红松和紫椴(Tilia amurensis),林分郁密度为0.8.林下灌木主要有刺五加(Acanthopanax senticosus)、毛榛子(Corylus mandshurica)、花楷槭(Acer ukurunduense)、山梅花(Philadelphus schrenkii)、瘤枝卫矛(Euonymus pauciflorus)、黄花忍冬(Lonicera chrysantha)、金银忍冬(Lonicera maackii)、小檗(Berberisamurensis)、东北茶藨子(Ribes mandshuricum)、刺醋李(Ribes burejense)等。草本主要有凸脉苔草(Carex lanceolata)、毛缘苔草(Carex pilosa)、乌苏里苔草(Carex ussuriensis)、羊胡子苔草(Carex callitrichos)、透骨草(Phryma leptostachya)、龙常草(Diarrhena manshurica)、舞鹤草(Maianthemum bifolium)、山酢酱草(Oxalis acetosella)等。
2.2 数据收集
2012年4 月中旬,在红松萌动前,在样地内选择6株生长健康、树干较直的样木(表1),在胸径处安装带状树干径向变化记录仪,对树木生长进行连续监测。具体安装方法是,刮去垂直于地面1.5 m处树干的粗死树皮,安装探头及金属线,调节电阻使初始值在2400 Ω左右。每30 min记录存储一次数据。记录仪的工作原理是:由于树干的径向膨胀与收缩变化会对固定在树干上的探头产生压力,这种压力与传感器电阻值之间有很好的线性关系,利用这种关系可以检测树干微小的径向变化,然后通过与数据采集器连接,定时记录径向数据,从而实现树木径向变化的连续动态监测。
数据气象资料使用距样地3 km的凉水气象站(47°11'08″N,128°53'06″E)的数据,每小时记录1次数据。
2.3 红松径向生长期的确定
由于可逆的膨胀和收缩,从树干径向变化生长仪数据直接判断树木生长关键事件时,如径向生长的开始、结束,会有些困难。本实验参照Deslauriers[12]的研究方法,用连续15 d的时间尺度(即把15 d提取的径向变化数据相加)来确定生长季节的开始与结束,使用单样本的t-检验用来评估15 d的径向变化均值是否等于零(μ=0),当值的分布是显著大于零时认为开始发生径向生长。
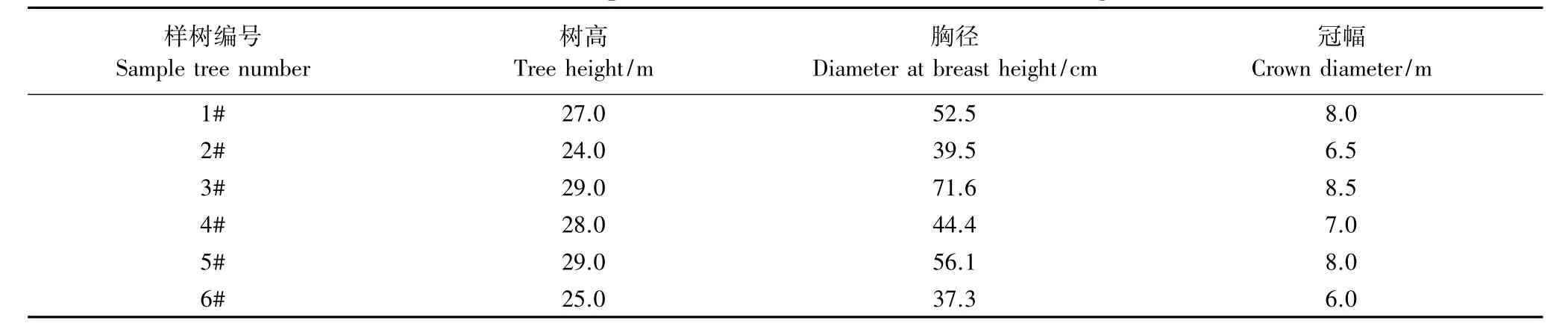
表1 2012年进行树干径向变化测定的6株红松样树特征Table1 Characters of the 6 sample trees of Pinus koraiensis for diameter change monitored in 2012
2.4 径向生长的拟合
本研究使用Gompertz生长模型对生长季红松径向生长进行拟合。很多研究表明Gompertz模型比其他“S”型函数更适用于树木的径向生长的拟合[2,9,13]。函数公式如下:

式中,Y为茎干生长累积量;t表示时间,生长开始的时间为0;A为生长累积量的上渐近线;β为与Y初始值有关的参数;k表示内秉生长率,e是自然对数的底数。
2.5 径向生长增量的提取以及与气象因子的分析
本研究使用两种径向生长增量提取方法,分析与气候因子的关系。
(1)周期循环法
树干直径变化包括两部分,一部分是由于新细胞的形成、扩大、细胞壁增厚等引起的不可逆生长;另一部分是由于树干含水量的变化引起的可逆的直径变化。树干径向生长增量的提取是使用Downes等[1]的方法进行的,将生理周期分为时间约24 h的3个不同阶段(图2)。生理周期不同阶段的区分,允许提取径向生长增量以及更好的与气象数据结合。3个不同阶段分别为:1)收缩阶段,树干直径最大值与最小值之间的这段时间;2)膨胀阶段,树干直径由最小值恢复到前一循环最大值之间的时间;3)增长阶段,树干直径超过膨胀阶段最大值到这一循环最大值的时间。收缩量、膨胀量和增长量分别指的是这3个阶段期间的径向变化量。径向生长增量一直是通过比较相邻周期最大值估计的,当没有超过前一周期最大值时,计算的树干半径减少(ΔR-,mm);当超过前一周期最大值时,计算的树干半径增加(ΔR+,mm)。
(2)最大值法
本研究又依据Bouriaud等[5]的最大值法,从红松茎干生长数据中提取日最大值,计算连续两日的最大值之差求得树木茎干的日变化,分析红松日径向变化与日气象因子的关系。
气象因子选取平均气温(Tmean,℃)、最高气温(Tmax,℃)、最低气温(Tmin,℃)、降雨量(P,mm)、相对湿度(RH,%)、土壤10 cm深处的温度(Tsoil,℃)以及水汽压亏缺(VPD,kPa)进行统计分析。水汽压亏缺是基于空气相对湿度和气温计算出来的。
本文涉及的分析使用SPSS 18.0软件完成,图使用Origin 8.0完成。使用Pearson相关与偏相关分析径向变化量与气象因子的关系,当日生长量为负值时,负日生长量不参与与气象因子关系的分析;利用逐步回归分析建立多元线性模型。
3 结果
3.1 径向生长日变化
树干日径向变化周期循环一般历时24 h,但是降雨以及冻融会造成较长的收缩或膨胀,使周期时间超过24 h[14]。那些持续(24±3)h的周期被称为正常周期(REG),持续时间超过28 h的循环称为长周期(LG),持续时间少于20 h为短周期[1,12]。本研究统计了生长期内各个阶段的开始时间。结果表明,在每日的8:00—10:00时,由于树木的蒸腾作用加强和液流速率增加,使茎干组织内产生了水势梯度,茎干形成层外侧的组织(韧皮部、周皮)因水势差脱水,从而使树干半径减少;在16:00—19:00时,树木茎干直径降至一天中的最低值,之后由于蒸腾作用的减弱以及根系的吸水补充,茎干形成层外侧的组织吸水膨胀,使树干半径增加[15];在次日1:00—4:00时开始进入增长阶段,持续到8:00时左右,然后进入下一个循环(图2)。
3.2 径向生长期的确定
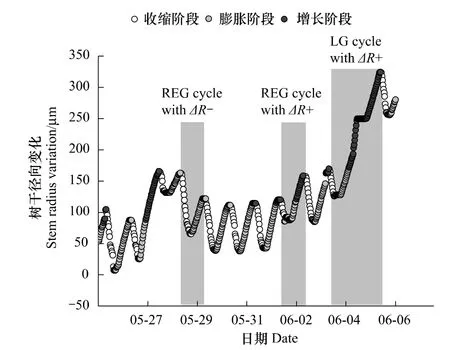
图2 红松树干日径向变化,当径向生长超过上一个周期最大值时,树干半径增加(ΔR+);当没有超过上一个周期最大值时,树干半径减少(ΔR-);正常周期(REG)一般持续24 h左右,而长周期(LG)持续时间则会超过24 h。Fig.2 Daily variation of stem radial of Pinus koraiensis;The stem radius increment(ΔR+)was calculated when the stem radius exceeds the previous cycle maximum,A△R-was calculated when the previous cycle maximum was not reached.A regular cycle(REG)lasts about 24 h,while a long cycle(LG)lasts more than one day
于2012年早春树液开始流动前将树干径向自动记录仪开启,开始记录。图3为整个生长季红松树干径向累计变化动态,可见6—7月份是树干直径生长的主要时期。从4月中旬到10月中旬,使用15 d的时间尺度对红松径向变化进行了分析。结果如图4所示:在4月中下旬有一个与0有显著差异的正值,这属于早春茎干水分恢复阶段。之后从5月中下旬开始连续出现2个与0有显著差异的正值,在7月中下旬出现最后一个与0有显著差异的正值,综合判断红松的径向生长开始于5月中旬,在7月末生长趋于结束。
3.3 红松径向生长的拟合
使用Gompertz模型对红松径向生长进行了拟合(图5)。从图中可以看出Gompertz生长模型很好的拟合了红松的径向生长,3株红松样木的方程拟合度达到了极显著水平(P<0.01),可以解释92%以上的红松径向变化。红松径向生长在5月中旬开始,从5月末(第150天左右)开始快速生长,在6月10日(第160天)左右径向生长速率达到最大值,然后7—8月是缓慢生长期。从图5还可以看出,3株红松的径向变化存在着明显的个体差异。1号和3号样木生长主要集中在一年的第145天(5月末)—185天(6月末),径向生长速率曲线较陡,生长量分别为882.74 μm和577.89 μm(图5A,C)。而2号样木径向生长从第140天(5月下旬)一直持续到第200天(7月中旬),径向生长速率曲线平缓,生长量为1007.71 μm(图5B)。

图3 红松树干径向累积变化Fig.3 Cumulative radial variation of Pinus koraiensis in 2012箭头表示估计的径向生长开始和结束的时间
3.4 径向生长与气候因子的分析
表2给出了径向生长期间,周期循环3个阶段与气候因子的Pearson相关与偏相关系数。从Pearson相关系数可以来看,收缩阶段的树干收缩量与这一时期的最高气温、水汽压亏缺呈正相关,与最低气温、相对湿度、土壤温度以及降雨量呈负相关。膨胀阶段的树干恢复与水汽压亏缺、降雨量呈正相关,与平均气温、最低气温、相对湿度、土壤温度呈负相关。增长阶段的树干径向生长与最高气温、相对湿度、土壤温度、降雨总量呈正相关,与水汽压亏缺呈负相关。

图4 2012年4月15日—10月12日,以15 d为1组的红松树干径向变化分布Fig.4 Stem radius variation distribution of Pinus koraiensis grouped at 15 d from 15 April to 12 October 2012
表3则给出了日径向变化与气候因子的关系。从Pearson相关系数来看,日径向变化与每天的相对湿度、最低气温和降雨量呈显著正相关,而与水汽压亏缺和日最高气温呈负相关。
利用逐步回归分析分别建立多元线性模型,得到如下3个模型:

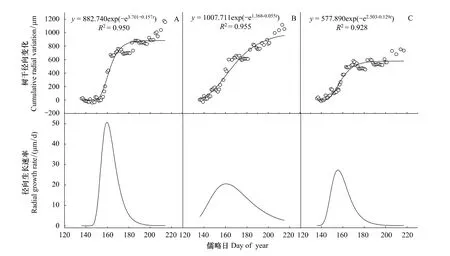
图5 生长季三株记录完整的红松径向累积变化与径向生长速率Fig.5 Curve fittings for cumulative radial variation and radial growth rate of Three well-documented Pinus koraiensis during the growing season
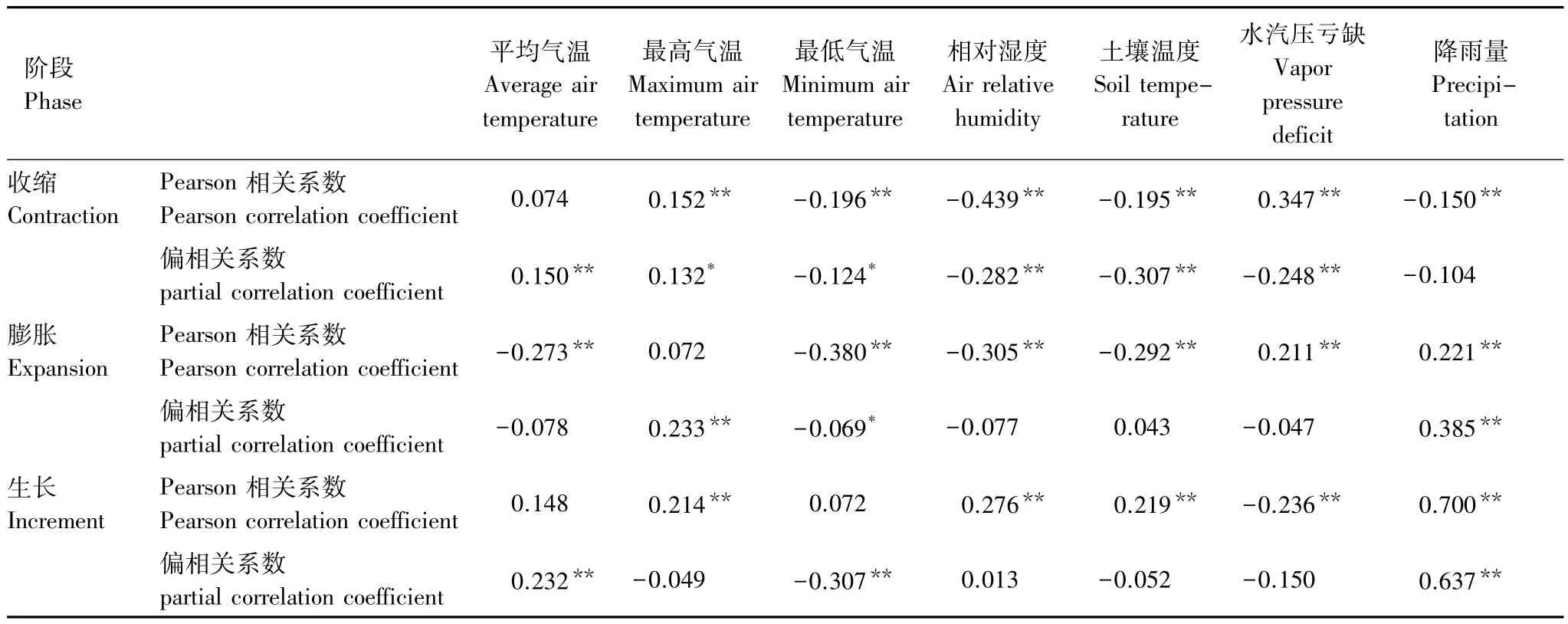
表2 生长季3个阶段径向变化量与相应阶段气候因子的Pearson相关性与偏相关系数Table2 Pearson correlation and partial correlation coefficient between three phase radial variation and the corresponding phase of meteorological factors during the growing season

表3 生长季红松日径向变化与日气象因子的Pearson相关性与偏相关系数Table3 Pearson correlation and partial correlation coefficient between daily stem growth of Pinus koraiensis and daily meteorological factors
公式2表示的是周期循环法3个阶段的气象因子对红松径向生长影响的多元线性模型;公式3表示的是膨胀和增长阶段的气象因子对红松径向生长影响的多元线性模型。公式4表示的是日气象因子对红松日径向变化的影响(日最大值法)。式中,Y1,Y2,Y3表示树干直径生长量(μm),P表示降雨量(mm),VPD表示水汽压亏缺(KPa),Tmean表示平均温度(℃),Tmin表示最低温度(℃),Tmax表示最高温度(℃),RH表示空气相对湿度,Tsoil表示土壤温度(℃)。公式2和公式3中的下标1,2,3分别表示周期循环法的3个阶段。
4 讨论
4.1 径向生长期的确定
确定生长的开始与结束对于研究径向生长与气候因子的关系是重要的[16-17]。尤其是树木生长缓慢的寒冷地区,树干水分储存的波动会对径向生长的提取和一些关键事件(如径向生长的开始,增长,结束)的识别造成影响。树木茎干经过冬天的严重缩水,会在春季进行快速的补水,树干迅速增粗,造成生长开始的假象。当试图计算寒冷环境下的径向生长时,从径向变化记录仪数据中鉴定这一时期至关重要,因为补水阶段可能很容易与径向生长的开始相混淆[2,18]。
本研究结果表明,红松径向生长累积增量在5月初至5月中旬,有一段相对稳定的平台期(图3),这段时间的记录有助于生长期开始的鉴定。Deslauriers等[12]提出大于10 d的时间尺度能够协助识别较低海拔的生长开始和茎干生长的主要时期。在较高海拔,大于15 d的时间尺度只能有助于生长主要时期的确定。根据样地所处海拔(359 m)及所处的高纬度特点,本研究使用15 d的时间尺度确定红松的径向生长期,在经过半月的平台期后,红松在5月中旬开始径向生长。在6月中旬至7月中旬的径向变化与0没有显著差异,造成这一结果的原因可能是6月上旬频繁的降雨量,一方面使土壤含水量的增加以及叶片和枝干表皮对树干径流的吸收,促进了细胞的伸长生长[19-20];而另一方面则是红松树皮吸水膨胀造成的可逆的增长,在降雨停止之后,树皮因脱水而收缩,造成树干直径的减少,从而使6月中旬至7月中旬的径向没有显著变化。在7月末出现最后一次显著的径向生长表明径向生长逐渐停止。
红松径向生长速率分析(图5)表明:红松最大生长速率出现在6月10日(第160天)左右。王业蘧[21]使用游标卡尺对小兴安岭凉水林场红松幼苗进行了研究,结果是6月和7月红松地径生长迅速,对全年地径总生长贡献最大;但王业蘧的研究结果表明7月份是红松径向生长的迅速期,与本研究6月中上旬是红松径向生长迅速期不同。本研究组同年用微树芯法研究了红松形成层的活动(未发表)表明:6月中旬至7月初是红松形成层分化、木质部细胞形成的主要时期,这与Rossi等[22]得出的高纬度和高海拔地区的树木径向生长高峰基本都出现在夏至日附近相一致。通过观察红松径向生长累积增量和降雨量的关系可以看出红松径向变化对降雨非常敏感,每一次大的降雨都会使径向生长发生显著变化,6月上旬的密集降雨使红松径向生长大幅增加(图3),从而得到的最大生长速率期提前。因此,虽然树木径向生长测量仪对树干径向持续变化的监测是其他方法无法比拟的,但是在应用上要十分谨慎,对于受降雨影响较大的树木,应当结合其他监测手段作对比。
4.2 红松径向变化与气候的关系
本研究通过两个方面对红松径向生长与气候因子的关系进行了分析。首先使用日最大值法分析红松的日径向变化与气象因子的关系,Pearson相关分析以及偏相关分析表明,红松日径向变化与日空气相对湿度,土壤温度以及降雨量的关系是极显著的。在相对湿度很高的情况下,树冠蒸腾作用减弱和根部吸水导致树干径向增加[7]。管伟和熊伟的研究也同样表明水分因子对落叶松日径向生长起着最为重要的作用[6-7]。生长季节的降雨对茎干径向生长的影响是重大的[1-2,7,9],一方面降雨能够增加土壤含水量,促进树干细胞的伸长生长[19-20],而另一方面则是由于树皮吸水膨胀而导致的对径向生长错误的估计[2]。土壤温度与净光合作用显著相关,影响着光合产物的运输与分配[23];同时,土壤低温还会抑制水分和养分的吸收[24],从而影响径向生长。
另外,本文通过树干循环周期法对径向生长进行了比较细致的研究,分析了3个阶段与相应气候因子的关系,结果显示不同阶段影响红松径向变化的环境因子并不相同。在收缩阶段,土壤温度影响着根系的吸水能力,能减小树干内部的水分胁迫,降低树干收缩。相对湿度提高,空气中水汽增大,使边界层水汽压与叶片气孔腔的水汽梯度减少,水汽化过程变慢,液流速度降低[25]。Herzog指出[15],树干径向日变化是与树干液流活动紧密相连的,在不考虑生长的情况下,树干径向变化是由树体水分含量变化引起的。土壤温度和相对湿度从不同的方面影响着树干体内的水分,从而与径向收缩呈显著相关。在膨胀和增长阶段,径向变化与最低气温极显著负相关,因为夜间温度影响树干形成层细胞伸长和增大,从而影响着树干径向生长[26-28],Xiaoxia Li等[29]研究了中国青藏高原东南部色季拉山的急尖长苞冷杉(Abies georgei var)之后,发现在高纬度和高海拔地区,急尖长苞冷杉幼树和老树都会对最低气温做出快速响应,认为最低温是限制树木生长最显著的气象因子。
多元线性模型(公式3)分析发现,第二和第三阶段的气象因子可以解释57.1%的径向变化。Dünisch[30]使用细胞分析和径向生长测量仪监测得出:当供水充足时81%的挪威云杉苗木径向细胞扩大发生在18:00至次日6:00之间。同样Deslauriers[2]在研究了加拿大冷杉(Abies balsamea(L.)Mill.)之后也得出了同样的结论,并且认为8:00至17:00收缩阶段的气象对径向生长的影响可能并不大。在模型中去除第一阶段的气象因子后(公式3),R2并没有明显的减少,也支持了这一结果。但是不论是公式2还是公式3,模型效果都明显优于公式4,公式4只解释了34.1%的日径向变化,说明周期循环法比日最大值法更加适合分析红松日径向变化和气象因子的关系。相比之下,Downes等[1]发现,增长阶段期间的平均气候并没有比日气象解释更多的生长变化;但Deslauriers[2]用自助法(Bootstrap)1 000次重采样计算响应函数,相关系数解释了90%以上的径向生长变化。在本研究中,红松径向生长并不是仅受气象因子的影响(气象因子最多解释58%的径向变化),可能还与树木本身的生物学特性和生理学特性有关,如树干形成层的活动与分化[1]、基因表达[31]和激素信号[32]等。因此要更好的理解气候对径向生长的影响,还需要对树木生物学和生理学特性进行更深入的研究。
5 结论
不论是周期循环法还是日最大值法都显示红松径向生长与气候因子有显著的关系,但是周期循环法能更加细化的分析日气象因子对径向生长的影响,能够解释更多的红松径向变化,其中膨胀和增长阶段的气象因子对红松径向生长的影响更大。在本研究中,降雨和最低气温是影响红松径向生长的关键气候因子。通过径向生长测量仪监测,红松径向生长在5月中下旬开始,在7月末趋于结束。生长模式为“S”形,最大径向生长速率出现在6月中上旬。径向生长测量仪的测定结果与直径生长的实际周期还存在一定差异,因此其应用还应结合其他监测手段。
[1] Downes G,Beadle C,Worledge D.Daily stem growth patterns in irrigated Eucalyptus globulus and E.nitens in relation to climate.Trees,1999,14(2):102-111.
[2] Deslauriers A,Morin H,Urbinati C,Carrer M.Daily weather response of balsam fir(Abies balsamea(L.)Mill.)stem radius increment from dendrometer analysis in the boreal forests of Québec(Canada).Trees,2003,17(6):477-484.
[3] Mäkinen H,Nöjd P,Saranpää P.Seasonal changes in stem radius and productionofnewtracheidsinNorwayspruce.Tree Physiology,2003,23(14):959-968.
[4] Tardif J,Flannigan M,Bergeron Y.An analysis of the daily radial activityof7borealtreespecies,northwesternQuebec.Environmental Monitoring and Assessment,2001,67(1/2):141-160.
[5] Bouriaud O,Leban J M,Bert D,Deleuze C.Intra-annual variations in climate influence growth and wood density of Norway spruce.Tree Physiology,2005,25(6):651-660.
[6] Guan W,Xiong W,Wang Y H,Yu P T,He C Q,Du A P,Liu H L.Stem diameter growth of Larix principis-rupprechtii and its response to meteorological factors in the north of Liupan Mountain.Scientia Silvae Sinicae,2007,43(9):1-6.
[7] Xiong W,Wang Y H,Yu P T,Liu H L,Shi Z J,Guan W.The growth in stem diameter of Larix principis-rupprechtii and its response to meteorological factors in the south of Liupan Mountain.Acta Ecologica Sinica,2007,27(2):432-441.
[8] Dong M Y,Jiang Y,Wang M C,Zhang W T,Yang H C.Seasonal variations in the stems of Larix principis-rupprechtii at the treeline of the Luya Mountains.Acta Ecologica Sinica,2012,32(23):7430-7439.
[9] Dong M Y,Jiang Y,Yang H C,Wang M C,Zhang W T,Guo Y Y.Dynamics of stem radial growth of Picea meyeri during the growing season at the treeline of Luya Mountain.Chinese Journal of Plant Ecology,2012,36(9):956-964.
[10] Chen L,Yin Y H,Pan T,Wu S H,Wang H.Growth of Pinus koraiensis and its response to temperature change at Changbai Mountain,Northeast China.Resources Science,2012,34(11):2137-2145.
[11] Wang X M,Zhao X H,Gao LS,Jiang Q B.Age-dependent growth responses of Pinus koraiensis to climate in the north slope of Changbai Mountain,North-Eastern China.Acta Ecologica Sinica,2011,31(21):6378-6387.
[12] Deslauriers A,Rossi S,Anfodillo T.Dendrometer and intraannual tree growth:What kind of information can be inferred?Dendrochronologia,2007,25(2):113-124.
[13] Rossi S,Deslauriers A,Morin H.Application of the Gompertz equationforthestudyofxylemcelldevelopment.Dendrochronologia,2003,21(1):33-39.
[14] Zweifel R,Häsler R.Frost-induced reversible shrinkage of bark of mature subalpine conifers.Agricultural and Forest Meteorology,2000,102(4):213-222.
[15] Herzog K M,Häsler R,Thum R.Diurnal changes in the radius of a subalpine Norway spruce stem:their relation to the sap flow and their use to estimate transpiration.Trees,1995,10(2):94-101.
[16] Worbes M.Annual growth rings,rainfall-dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela.Journal of Ecology,1999,87(3):391-403.
[17] Biondi F,Hartsough P C,Estrada I G.Daily weather and tree growth at the tropical treeline of North America.Arctic,Antarctic,and Alpine Research,2005,37(1):16-24.
[18] Zweifel R,Häsler R.Dynamics of water storage in mature subalpine Picea abies:temporal and spatial patterns of change in stem radius.Tree Physiology,2001,21(9):561-569.
[19] Zweifel R,Zimmermann L,Zeugin F,Newbery D M.Intra-annual radial growth and water relations of trees:implications towards a growth mechanism.Journal of Experimental Botany,2006,57(6):1445-1459.
[20] Zahner R.Internal moisture stress and wood formation in conifers.Forest Products Journal,1963,13(6):240-247.
[21] Wang Y J.Broad-leaved Korean Pine Forest.Harbin:Northeast Forestry University Press,1995:335-337.
[22] Rossi S,Deslauriers A,Anfodillo T,Morin H,Saracino A,Motta R,Borghetti M.Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length.New Phytologist,2006,170(2):301-310.
[23] Körner C.A re-assessment of high elevation treeline positions and their explanation.Oecologia,1998,115(4):445-459.
[24] Körner C.Alpine Plant Life:Functional Plant Ecology of High Mountain Ecosystems.New York:Springer-Verlag,1999.
[25] Ru T Q,Li J Y,Kong L S,Zhu Y L.Review on the research of water consumption characteristic of Robinia psedudoacia.Research of Soil and Water Conservation,2005,12(2):135-140.
[26] Denne M P.Temperature and tracheid development in Pinus sylvestris seedlings.Journal of Experimental Botany,1971,22(2):362-370.
[27] Antonova G F,Stasova V V.Effects of environmental factors on wood formation in Scots pine stems.Trees,1993,7(4):214-219.
[28] Richardson S D,Dinwoodie J M.Studies on the physiology of xylem development.I.The effect of night temperature on tracheid size and wood density in conifers.II.Some effects of light intensity,daylength and provenance on wood density and tracheid length in Picea sitchensis.III.Effects of tempe.Journal of the Institute of Wood Science,1960,(6):3-13.
[29] Li X X,Liang E Y,Gricˇar J,Prislan P,Rossi S,Cˇufar K.Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau.Tree Physiology,2013,33(1):48-56.
[30] Dünisch O,Bauch J.Influence of soil substrate and drought on wood formation of spruce(Picea abies[L.] Karst.)under controlled conditions.Holzforschung-International Journal of the Biology,Chemistry,Physics and Technology of Wood,1994,48(6):447-457.
[31] Schrader J,Nilsson J,Mellerowicz E,Berglund A,Nilsson P,Hertzberg M,Sandberg G.A high-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial stem cell identity.The Plant Cell Online,2004,16(9):2278-2292.
[32] Schrader J,Baba K,May S T,Palme K,Bennett M,Bhalerao R P,Sandberg G.Polar auxin transport in the wood-forming tissues of hybrid aspen is under simultaneous control of developmental and environmental signals.Proceedings of the National Academy of Sciences of the United States of America,2003,100(17):10096-10101.
参考文献:
[6] 管伟,熊伟,王彦辉,于澎涛,何常清,杜阿朋,刘海龙.六盘山北侧华北落叶松树干直径生长变化及其对环境因子的响应.林业科学,2007,43(9):1-6.
[7] 熊伟,王彦辉,于澎涛,刘海龙,时忠杰,管伟.六盘山南坡华北落叶松(Larix principis-rupprechtii)树干直径生长及其对气象因子的响应.生态学报,2007,27(2):432-441.
[8] 董满宇,江源,王明昌,张文涛,杨浩春.芦芽山林线华北落叶松径向变化季节特征.生态学报,2012,32(23):7430-7439.
[9] 董满宇,江源,杨浩春,王明昌,张文涛,郭媛媛.芦芽山林线白杄生长季径向生长动态.植物生态学报,2012,36(9):956-964.
[10] 陈力,尹云鹤,潘韬,吴绍洪,王辉.长白山红松生长量及其对温度变化的响应.资源科学,2012,34(11):2139-2145.
[11] 王晓明,赵秀海,高露双,姜庆彪.长白山北坡不同年龄红松年表及其对气候的响应.生态学报,2011,31(21):6378-6387.
[21] 王业蘧.阔叶红松林.哈尔滨:东北林业大学出版社,1995:335-337.
[25] 茹桃勤,李吉跃,孔令省,朱延林.刺槐耗水研究进展.水土保持研究,2005,12(2):135-140.
