氮素添加对中亚热带栎属不同树种幼苗生长及生物量分配的短期影响1)
2014-08-02王晓荣唐万鹏庞宏东郑兰英
王晓荣 潘 磊 唐万鹏 庞宏东 郑兰英
(湖北省林业科学研究院, 武汉,430075)
氮素添加对中亚热带栎属不同树种幼苗生长及生物量分配的短期影响1)
王晓荣 潘 磊 唐万鹏 庞宏东 郑兰英
(湖北省林业科学研究院, 武汉,430075)
为了探讨中亚热带栎属不同树种幼苗的生长和生物量分配对短期氮沉降的响应,选择小叶栎、栓皮栎、麻栎、短柄枹栎和白栎作为研究对象,设置了CK、N5、N15、N304个处理,分别以NH4NO3的形式施入土壤,相当于氮沉降率为0、5、15、30 g·m-2·a-1。经过4个月的氮添加处理发现:1)短期氮添加处理对5种栎属幼苗的地径、株高和叶数无显著性影响,仅显著促进栓皮栎和麻栎幼苗的分枝数的增加。除栓皮栎外,其他所有树种不同处理下茎相对生长率表现为先增加后降低的趋势,株高生长速率则无规律性变化。2)低、中浓度氮处理可以显著促进生物量的积累,而高浓度氮添加对生物量积累产生了一定的抑制作用,且栎属不同树种幼苗全株生物量累积对氮沉降的响应具有一定差异;3)氮沉降在一定程度上提高了植物的叶质量比和茎质量比,但短期氮沉降对其促进作用不十分明显,仅有栓皮栎和麻栎的茎质量比随氮浓度的增加而显著增加,同时氮添加降低了小叶栎、栓皮栎、麻栎和白栎的根质量比和根冠比,却使得短柄枹栎根冠比发生显著增加。
中亚热带;栎属;氮素添加;幼苗生长;生物量分配
近几十年来,随着人类活动的加剧,如化石燃料燃烧、含氮化肥的生产和使用及畜牧业集约化经营等,使得氮化物以大气沉降的形式不断向陆地生态系统累积[1-2],氮沉降增加已经成为全球化的生态环境问题[3-4]。有关研究表明,我国已成为全球三大氮沉降集中区之一(分别为欧洲、美国和中国),国内许多地区存在高氮沉降现象,如太湖地区的常熟生态站2001—2003年间大气氮湿沉降量约为27 kg·hm-2·a-1[5],珠江三角洲北缘的鼎湖山自然保护区1998—1999年度的降水氮沉降为38.4 kg·hm-2·a-1[6]。氮沉降的增加对陆地生态系统的影响,及其所带来的生态效应也逐渐成为人们关注和研究的热点[7-8]。
森林作为陆地中最重要的生态系统,在维护区域生态安全方面发挥着十分重要的作用。因此,在氮沉降全球化的环境背景下,研究和预测氮沉降对森林生态系统的影响及其反馈就显得尤为重要。许多学者已经开展了此方面的相关研究,包括对氮沉降下植物形态变化、生理生态响应、生物量分配格局、生产力、有机质分解、营养结构状况、抗逆性、生物多样性等诸多方面[4,7,9-12]。结果表明,氮沉降在一定范围内能够刺激植物生长,提高植物生产力[9,13],但当氮素在植物体内积累到一定程度出现氮饱和现象时,氮的输入反而会抑制植物的生长,甚至导致生态系统衰退[14-15]。究其原因发现,这是因为在氮沉降的临界负荷之内,适宜程度的氮沉降能改变植物的生物学特性、化学组成和生化过程并加速植物的生长,而如果超过了氮的临界负荷,营养缺乏和其他负面效果才使得植物生长缓慢[7,16]。然而,国内外关于亚热带森林植物对氮沉降的响应机制还存在很多不确定性[4,10,17],氮素对植物生长的影响因森林类型、植物种类和氮的输入量、生长条件和实验设施等诸多因素的差异而不同,特别是不同树种的生长及生理反应与其生态适应能力有关[17]。以往关于氮沉降的研究多以单一树种或者不同树种为对象[4,18-20],将同属不同树种相结合研究还比较少见,同属物种常常具有相似的生境要求,但又有自身独特的生态适应特征,是进行模拟实验的天然材料。
壳斗科栎属(Quercus)植物是我国亚热带最重要的森林树种,在天然林和人工林中往往成为优势种和建群种。本文选择中亚热带广泛分布的小叶栎(Q.chenii)、麻栎(Q.acutissima)、栓皮栎(Q.variabilis)、短柄枹栎(Q.glanduliferavar.brevipetiolata)、白栎(Q.fabri)作为研究对象,通过研究短期氮素添加对栎属不同树种幼苗期生长变化特征以及生物量积累和分配格局的影响,了解植物对氮沉降的初始反应状况,以探讨中亚热带森林幼苗对短期氮沉降的响应机制,从而为该区域气候变化下森林的培育和管理提供科学依据。
1 研究地概况
研究地点位于湖北省武汉市郊区的九峰国家森林公园,地理中心位置为北纬30°30′33″~30°31′04″,东经114°29′50″~114°30′26″。气候属亚热带季风性湿润气候,具有热富雨丰,雨热同季,四季分明的特点。全年平均气温16.7 ℃,极端最高气温为41 ℃,极端最低气温-17.6 ℃,年日照时间1 600 h,无霜期235 d,年降水量1 200~1 400 mm。由于工农业和城市化的发展,本区域大气污染相当严重,武汉市郊区2005年降雨氮沉降总量为26.07 kg·hm-2·a-1,2006年则上升为33.09 kg·hm-2·a-1,与全国湿沉降最高值相近,远高于同期欧洲和美国的氮沉降量[11],已经成为我国典型的高氮沉降区域。
2 材料与方法
于2011年9中旬,采集九峰山生长的小叶栎、麻栎、栓皮栎、短柄枹栎、白栎当年所产种子,种子选取自行脱落、种皮饱满有光泽、无虫蛀的成熟种子,经过沙藏处理后播于湖北省林业科学研究院温室内进行育苗生长。2012年4月份选择株高与地径大小及生长状况基本一致的苗木,每个物种各选择48株,共计240株,移栽于直径17.2 cm,高15 m的生长盆内,用V(九峰山内土壤)∶V(草炭土)=2∶1混合的土壤进行栽植,且放置在自然环境下生长。待实验材料生长稳定后,设置4个氮沉降处理水平:CK(对照)、N5(低氮)、N15(中氮)、N30(高氮),每个处理12盆。7月初开始,每月初和月中向幼苗土壤浇灌NH4NO3溶液,溶液浓度为0、0.013 75、0.027 5、0.041 25 mol·L-1,每盆每次50 mL,保证所浇灌溶液不会造成烧苗现象发生,共施氮6次(以代表年氮沉降量),累计施氮量折算为氮沉降率分别是0、5、15、30 g·m-2·a-1,不包括大气沉降的氮含量,这些施氮量是基于本地区的氮沉降量和国际上同类研究的情况而确定[4,17]。
2.1 生长指标测量
从控制实验开始采集生长数据,每隔30 d测量1次生长指标,共测定4次。生长指标包括株高、基径、叶片数、分枝数等。
相对生长速率。生长期不同处理植株的株高、基径的相对生长速率按以下公式计算,R=(lnH2-lnH1)/Δt,其中H1和H2分别表示前后两次测量时的株高、基径,Δt表示测量间隔时间。
2.2 生物量测定
于2012年植株生长末期,每个处理每个物种随机各抽取5株,应用全收割法,将幼苗整棵挖出清洗干净后,按叶、茎、根进行鲜质量称量,之后放置在烘箱中80 ℃烘干至恒质量,分别称取其干质量,计算叶生物量、茎生物量、根生物量、叶质量比(叶干质量/全株干质量)、茎质量比(茎干质量/全株干质量)、根质量比(根干质量/全株干质量)、根冠比(根干质量/地上部分干质量)。
2.3 数据统计分析
数据统计均采用SPSS 16.0和Excel 2007软件进行处理和分析,对生长数据和生物量采用单因素方差分析(one-way ANOVA),且进行LSD多重比较,以检验数据之间的差异显著性(P=0.05);采用SigmaPlot 10.0作图。
3 结果与分析
3.1 短期氮处理对栎属幼苗形态特征影响
如表1可知,短期不同氮水平添加处理对5种栎属幼苗的地径、株高和叶片数无显著性影响(P>0.5)。仅有麻栎和栓皮栎幼苗的分枝数CK组与氮处理组均表现出显著差异(P>0.5),麻栎N5和N15分别较对照增加了23%和54%,栓皮栎N15和N30分别交对照增加了3.76%和9.77%,可见短期氮沉降仅在一定程度上促进了栓皮栎和麻栎幼苗的分枝数的增加,而其他树种没有显著变化,这可能主要与树种生物特性相关。另外还可发现,5种树种幼苗分枝整体变化趋势与基径和株高相反,基径和株高生长较好的幼苗分枝较少,反之幼苗分枝较多。
3.2 短期氮处理对栎属幼苗相对生长速率的影响
从图1中可以看出,除栓皮栎在中高氮水平一直保持快速生长外,其他所有树种不同处理下茎相对生长率表现为先增加后降低的趋势,均在8—9月份生长最快,10—11月份生长速度又开始下降,但高氮水平均促使其季末生长速度高于对照,说明栎属植物幼苗在生长季后期,基径生长速度虽然变慢,但是氮添加有利于提高它的生长,或者说在一定程度上延缓了植株的衰落。不同氮处理水平下,5种栎属幼苗的株高生长均没有表现出规律性的变化趋势,但在10—11月份生长末期,株高生长速率在不同氮沉降水平基本保持一致,说明此时外界气候及物种生物学特性起着关键性作用,株高生长基本停滞。
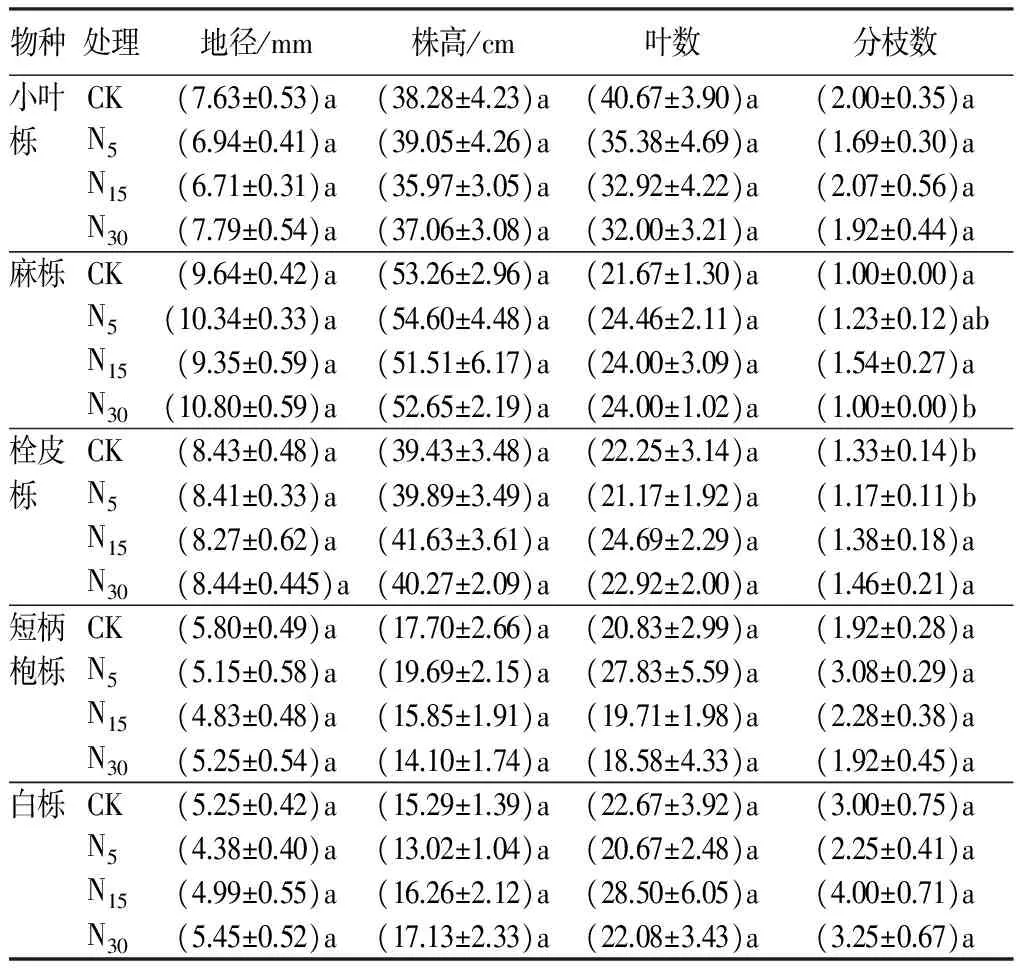
表1 氮素添加对栎属不同树种幼苗生长的短期影响
注:表中数据平均值±标准差;同列不同字母表示不同氮添加量的影响差异显著(P<0.05)。
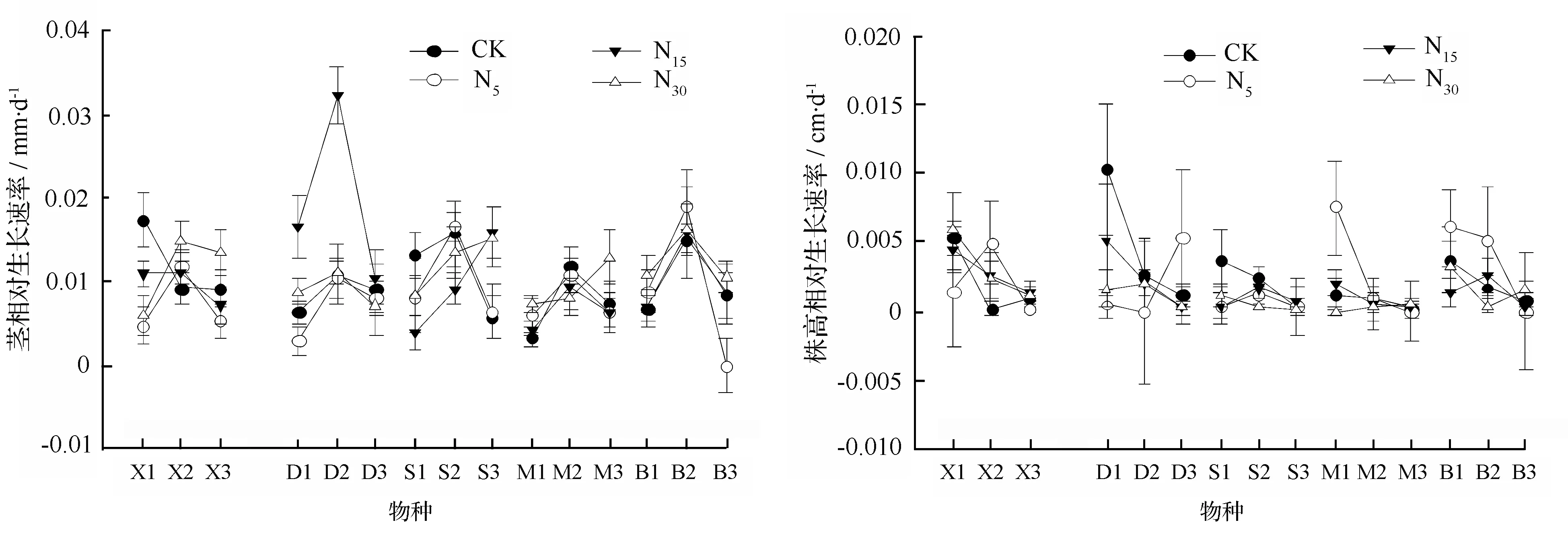
图中X、D、S、M、B分别代表小叶栎、短柄枹栎、栓皮栎、麻栎、白栎;1,2,3分别代表8—9月、9—10月和10—11月相对生长速率。
3.3 短期氮处理对栎属幼苗生物量的影响
从表2可知,短期氮沉降对栎属不同树种幼苗全株生物量影响程度具有一定差异,分别是小叶栎变化趋势为N5>CK>N15>N30;栓皮栎和短柄枹栎为N5>N15>CK>N30,而麻栎为N15>N5>CK>N30,此3种树种低、中浓度氮处理组与对照组和高浓度氮处理组均表现为显著性差异(P<0.05),分别较对照增加了10.82%、9.07%、-20.45%,52.26%、49.07%、-18.22%和42.97%、18.85%、-0.21%;白栎为N15>N30>N5>CK,氮处理组与对照组变现出显著性差异(P<0.05),分别较对照增加了99.81%、88.39%、10.86%。从中可以发现,除白栎高浓度氮沉降对植株生物量积累表现为促进作用外,其他4种均表现为高浓度氮沉降对生物量积累产生了一定的抑制作用,而低中浓度氮处理可以显著促进生物量的积累。
注:表中数据为平均值±标准差;同列不同字母表示不同氮添加量的影响差异显著(P<0.05)。
3.4 短期氮处理对栎属幼苗生物量分配格局的影响
从表3可知,不同水平氮处理均对各树种叶质量比、茎质量比和根质量比具有不同程度的影响作用,其中氮浓度一定程度上促进了叶质量比的增加,栓皮栎中氮和高氮水平与对照存在显著差异外(P<0.05),其他物种均无显著性差异,说明短期氮沉降对不同树种叶生物量的分配比例影响不明显。对于茎质量比,土壤氮素的增加对栓皮栎和白栎均存在明显的促进作用,表现为随着氮浓度的增加而显著增加,其他3种的茎质量比在不同处理间未表现出显著性差异,但仍可以看出土壤氮素的增加对其茎生物量积累存在一定的促进作用。对于不同树种幼苗根质量比而言,除短柄枹栎根质量比在高氮水平促进其比例显著增加(P<0.05),低氮和中氮水平则变化较小外,其他4树种幼苗根质量比和根冠比则均发生降低,其中小叶栎和麻栎幼苗根质量比对氮素添加无显著性差异(P>0.05),栓皮栎和白栎则中高浓度氮水平下幼苗根质量比显著减小(P<0.05),分别较对照降低了5.97%、14.92%和14.28%、14.28%,说明短期氮沉降可导致植物根质量比和根冠比具有减小的作用。
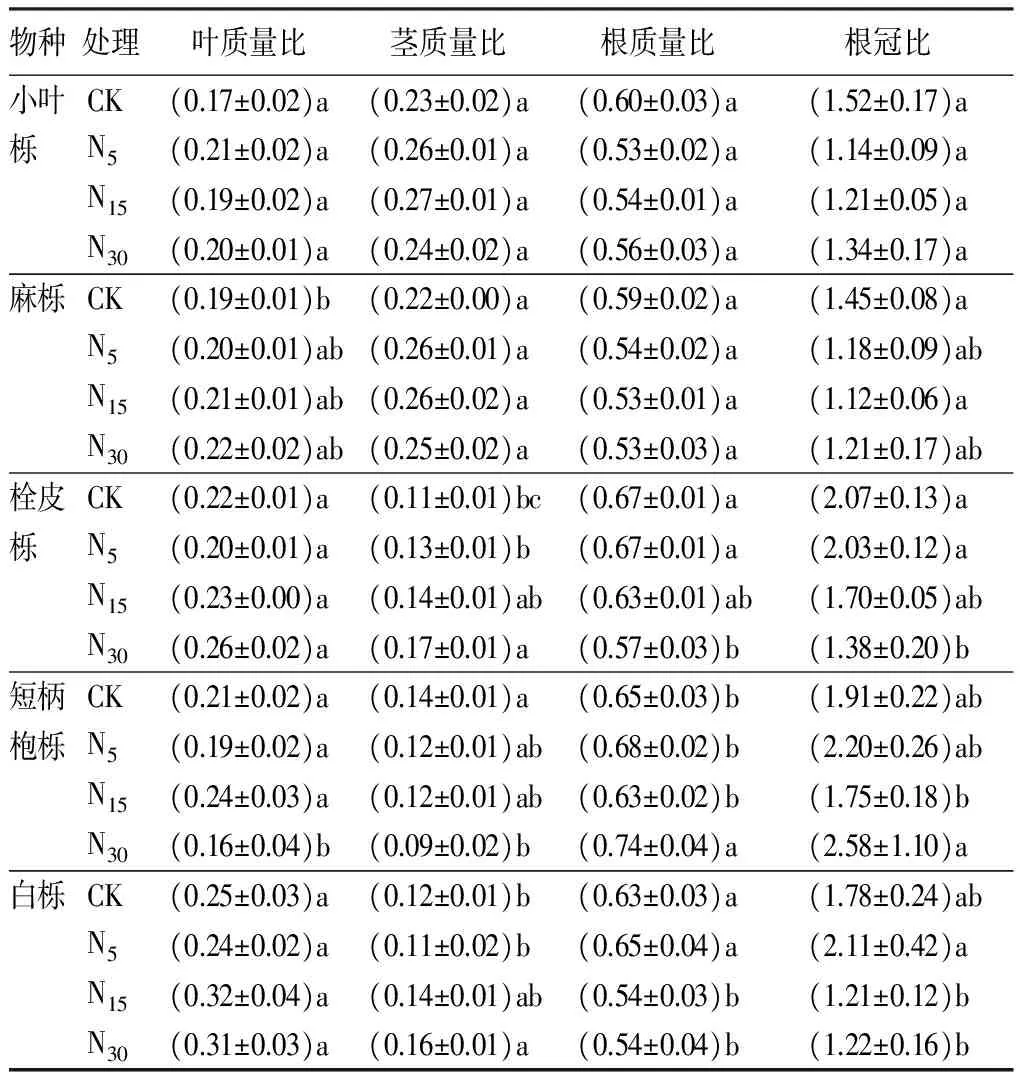
表3 氮素添加对栎属不同树种幼苗生物量分配比例的影响
注:表中数据为平均值±标准差;同列不同字母表示不同氮添加量的影响差异显著(P<0.05)。
4 结论与讨论
以往的研究发现,植物的形态适应可能是植物应对不良环境的最主要适应机制[18]。有效的氮沉降往往引起土壤可利用性氮的增加,直接影响着植物生态发育发生一定变化[17],特别是植物株高、地径、叶数和分枝数等个体形态结构对氮沉降的响应较为敏感,但氮素对植物生长的影响因森林类型、植物种类、氮的输入量以及氮沉降时间长短而不同[7,17],因此研究植物对氮环境变化产生的形态可塑性对于揭示植物对氮沉降的影响机制具有重要意义。本研究中,随着氮素添加浓度的增加,栎属不同树种基径、株高和叶数未表现出规律性的变化趋势,但短期氮处理却在一定程度上增加了幼苗的分枝数。同时,高氮水平促使树种在生长末期的茎相对生长速率高于对照,说明氮添加有利于提高它的生长,或者说在一定程度上延缓了植株的衰落,而对株高生长速率却没有规律性的变化,这与李德军等[4]在研究氮沉降对南亚热带两种乔木幼苗生长发育的结果存在一定差异,而与窦晶鑫等[16]研究小叶章对氮沉降生理生态的影响的结果一致,这可能与树种自身生态学特性以及氮处理时间长短有关。
氮素是影响森林植物生长发育的重要的生态因子,氮沉降适当增加刺激植物生长,提高其初级生产力的累积,特别是在氮限制的生态系统影响更为明显[9,13],而高氮沉降和长期氮沉降反而会产生负面效应,造成氮刺激的植物生长量减少或者无影响。因此,氮沉降将导致植物生产力增加还是减少,完全取决于这些植物所处的生态系统的氮供应情况[4,15,17,21]。大量的研究表明,氮输入的增加会明显促进亚热带森林植物的生长,导致生物量累积的增加[4,10,17],认为亚热带的高氮沉降区域一定氮沉降仍可促进植物幼苗生长,且以中等程度氮处理的效果最佳[4],随着氮素在植物体内的累积,植物生长则会受到抑制作用,而Matson等[22]认为绝大部分亚热带森林植物的生长不受氮的限制,氮沉降的增加可能不会促进植物生长,甚至会引起土壤酸化以及盐基阳离子的可利用性降低,对植物生长产生不利影响。本研究的结果与前者相一致,除白栎高浓度氮沉降对植株生物量积累表现为促进作用外,其他4种均表现为高浓度氮沉降对生物量积累产生了一定的抑制作用,而低中浓度氮处理可以显著促进生物量的积累。
以往研究证明,外源施氮增加了土壤中可利用氮的含量,必然会引起植物体内氮的积累,引起植物体内的生物量分配格局发生变化[23],叶质量比、茎质量比和根质量比则反映了生物量在叶、枝和根不同器官之间分配的比例[17]。本研究中,不同氮处理水平对栎属各树种叶质量比、茎质量比和根质量比均具有不同程度的影响作用,短期氮添加对叶生物量和茎生物量的分配比例具有促进作用,反而降低了根质量比,但不同树种生物量分配存在一定差异,这与很多研究结果相一致[4,16]。随着氮浓度的增加,对栓皮栎和白栎的茎质量比存在明显的促进作用,其他3种的茎质量比在不同处理间没有表现出显著性差异(P<0.05),仍在一定程度上促进了茎生物量的增加。这是因为氮素是植物光合色素合成的重要元素,而地上植物叶和枝是光合合成竞争资源的有效构件,短期氮添加刺激了植物光合产物的产生,使得叶和茎生物量积累发生增加[17,24]。本研究中小叶栎、栓皮栎、麻栎和白栎根冠比降低的现象与Persson等[24]的研究结果相一致,他们认为氮沉降导致森林植物根冠比下降的原因主要在于氮添加改变了土壤的理化性质,而土壤的理化性质又与根的生长和结构显著相关。当土壤有效氮不足时,根系作为植物获取地下资源(养分和水分)的主要构件,必须通过扩大根系生长来吸收更多的养分,而当土壤有效氮比较充足时,且能满足植物生长和发育所需的养分,光合产物向根分配的比例就会减少,而向地上部分分配的比例就会增加[17]。同时,本研究也发现氮沉降使得短柄枹栎的根冠比显著增加,短期氮添加浓度的增加不会限制短柄枹栎根部的生长。可见森林植物对土壤养分状况应答策略存在不同,导致其生物量分配比例的不同,反映了植物应对氮沉降响应存在一定的差异。
本研究中,栎属不同树种生长特征对短期氮素添加响应不够敏感,没有表现出显著的变化趋势,而生物量积累则反映出了一定的变化,说明短期适量氮素添加可以使得树种干物质累积增加,过量氮素反而明显发生抑制作用,且不同树种对氮素增加的响应存在差异。然而,本模拟试验时间相对较短,相对树木较长的生命周期而言,试验结论还不能够充分揭示中亚热带栎属树种对氮沉降的响应,仅可以代表处于幼苗阶段对氮增加的结果。同时,所采取施氮方式是以土壤表面氮添加来模拟,而地上植物营养部件直接受到的影响没有直观的反映。因此,未来氮沉降对幼苗幼树阶段的研究应该以生长在自然环境中的林分进行长期定位研究,尽量结合当地降水变化进行外源氮的施加,所得结果会更加合理。
[1] 赵亮,周国逸,张德强,等.CO2浓度升高和氮沉降对南亚热带主要乡土树种及群落生物量的影响[J].应用生态学报,2011,22(8):1949-1954.
[2] 张蕊,王艺,金国庆,等.氮沉降模拟对不同种源木荷幼苗叶片生理及光合特性的影响[J].林业科学研究,2013,26(2):207-213.
[3] 常运华,刘学军,李凯辉,等.大气氮沉降研究进展[J].干旱区研究,2012,29(6):972-979.
[4] 李德军,莫江明,方运霆,等.模拟氮沉降对南亚热带两种乔木幼苗生物量及其分配的影响[J].植物生态学报,2005,29(4):543-549.
[5] 王小治,朱建国,高人,等.太湖地区氮素湿沉降动态及生态学意义:以常熟生态站为例[J].应用生态学报,2004,15(9):1616-1620.
[6] 周国逸,闫俊华.鼎湖山区域大气降水特征和物质元素输入对森林生态系统存在和发育的影响[J].生态学报,2001,21(12):2002-2012.
[7] 吕超群,田汉勤,黄耀.陆地生态系统氮沉降增加的生态效应[J].植物生态学报,2007,31(2):205-218.
[8] Reay D S, Dentener F, Smith P, et al. Global nitrogen deposition and carbon sinks[J]. Nature Geoscience,2008,1(7):430-437.
[9] 李德军,莫江明,方运霆,等.氮沉降对森林植物的影响[J].生态学报,2003,23(9):1891-1900.
[10] 李德军,莫江明,方运霆,等.模拟氮沉降对三种南亚热带树苗生长和光合作用的影响[J].生态学报,2004,24(5):876-882.
[11] Zhao C S, Hu C X, Huang W, et al. A lysimeter study of nitrate leaching and optimum nitrogen application rates for intensively irrigated vegetable production systems in Central China[J]. Journal of Soils and Sediments,2010,10(1):9-17.
[12] 陈琳,曾杰,徐大平,等.氮素营养对西南桦幼苗生长及叶片养分状况的影响[J].林业科学,2010,46(5):35-40.
[13] Matson P, Lohse K A, Hall S J. The globalization of Nitrogen deposition: Consequences for terrestrial ecosystems[J]. Ambio,2002,31(2):113-119.
[14] Neff J C, Townsend A R, Gleixner G, et al. Variable effects of Nitrogen additions on the stability and turnover of soil Carbon[J]. Nature,2002,419:915-917.
[15] Nordin A, Strengbom J, Witzell J, et al. Nitrogen deposition and the biodiversity of boreal forests: implications for the Nitrogen critical load[J]. Ambio,2005,34(1):20-24.
[16] 窦晶鑫,刘景双,王洋,等.小叶章对氮沉降的生理生态响应[J].湿地科学,2009,7(1):40-46.
[17] 吴茜,丁佳,闫慧,等.模拟降水变化和土壤施氮对浙江古田山5个树种幼苗生长和生物量的影响[J].植物生态学报,2011,35(3):256-267.
[18] 刘洋,张健,陈亚梅,等.氮磷添加对巨桉幼苗生物量分配和C∶N∶P化学计量特征的影响[J].植物生态学报,2013,37(10):933-941.
[19] Villagra M, Campanello P I, Bucci S J, et al. Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species[J]. Tree physiology,2013,33(12):1308-1318.
[20] Santiago L S, Wright S J, Harms K E, et al. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition[J]. Journal of Ecology,2012,100(2):309-316.
[21] Patterson T B, Guy R D, Dang Q L. Whole-plant nitrogen-and water-relations traits, and their associated trade-offs, in adjacent muskeg and upland boreal spruce species[J]. Oecologia,1997,110(2):160-168.
[22] Matson P A, Mcdowell W H, Townsend A R, et al. The globalization of N deposition: ecosystem Consequences in tropical environments[J]. Biogeochemistry,1999,46(1/3):67-83.
[23] Berger T W, Glatzel G. Response of Quercus petraea seedlings to nitrogen fertilization[J]. Forest Ecology and Management,2001,149(1/3):1-14.
[24] Persson H. Ahlström, A and Clemensson-Lindell. Nitrogen addition and removal at Gårdsjön-effects on fine-root growth and fine-root chemistry[J]. Forest Ecology and Management,1998,101(1/3):199-205.
Short-term Effects of Nitrogen Addition on Seedling Growth and Biomass Allocation with Different Tree Species ofQuercusin the Mid-Subtropics of China/Wang Xiaorong, Pan Lei, Tang Wanpeng, Pang Hongdong, Zheng Lanying
(Forestry Institute of Hubei Province, Wuhan 430075, P. R. China)//
Journal of Northeast Forestry University.-2014,42(6).-24~28
We studied the seedling growth and biomass allocation of subtropicalQuercusspecies in response to short-term nitrogen deposition and plantedQ.variabili,Q.acutissima,Q.chenii,Q.glanduliferavar.brevipetiolataandQ.fabriwith four nitrogen treatments (CK, N5, N15, N30) to the soil with NH4NO3by simulating the N depositions of 0, 5, 15 and 30 g·m-2·a-1. After four months of nitrogen addition treatment, short-term nitrogen treatment has no significant effect on basal diameter, height and leaves ofQuercusseedlings, but it can promotes significantly to increase the branch number of seedlings. ExceptQ.variabilis, stem relative growth rate of the other species decrease after an initial increase, but the height relative growth rate has no significant variation pattern. The biomass accumulation can be promoted in low content of nitrogen treatment, while inhibited by high content.Quercusseedlings biomass accumulation shows a different trend response to nitrogen deposition. The leaf weight ratio and stem weight ratio are increased, but mostly not significant exceptQ.variabilisandQ.acutissima. Both root weight ratio and the ratio of root to shoot significantly increase forQ.glanduliferavar.brevipetiolataseedlings, but significantly decrease for other four species.
Mid-subtropics;Quercus; Nitrogen addition; Seeding growth; Biomass allocation
1) “十二五”国家科技支撑计划项目(2011BAD38B04)资助。
王晓荣,男,1984年10月生,湖北省林业科学研究院,助理研究员。
潘磊,湖北省林业科学研究院,研究员。E-mail:panlei2008@126.com。
2013年9月18日。
Q948.1
责任编辑:潘 华。
