广东连州田心自然保护区香果树种群及其生境特征
2014-07-16曾庆昌缪绅裕唐志信邓冬梅李春波黄福长
曾庆昌,缪绅裕*,唐志信,邓冬梅,李春波,黄福长
1. 广州大学生命科学学院,广东 广州 510006;2. 广东连州田心省级自然保护区管理处,广东 连州 513400
茜草科植物香果树(Emmenopterys henryi)为落叶大乔木,广泛分布于我国亚热带的浙江、安徽南部、福建北部和中西部、江西、湖北西部和南部、湖南、广东北部、广西北部、贵州、云南、甘肃东南部、陕西、河南、四川东南部和重庆,常生于海拔400~1500 m的深山沟谷阔叶林中,在贵州和云南可在海拔2000 m生长(国家林业局,2009;傅立国,1992)。香果树喜温和或凉爽的气候和湿润肥沃的土壤,分布区内年均温18~22 ℃,年降水量1000~2000 mm,相对湿度70%~85%,土壤为山地黄壤或沙质黄棕壤,pH5~6(国家林业局,2009;傅立国,1992)。
香果树为我国特有的单种属植物,对研究茜草科系统发育和我国南部、西南部的植物区系等均有一定意义(国家林业局,2009;傅立国,1992)。香果树在《广东珍稀濒危植物》中虽有收录,但未见具体分布地点和种群数量。1993年香果树已被列为国家Ⅱ级重点保护野生植物,生境破坏可能是致本种濒危的主要原因(彭少麟和陈万成,2003)。在《南岭植物物种多样性编目》中也仅记载分布于莽山(湖南境内。邢福武等,2012)。笔者在早期对粤北珍稀濒危植物种群的研究中,也未见香果树的自然分布(缪绅裕等,2008)。目前所见文献中,除笔者报道在连州田心保护区有自然分布的1株外(缪绅裕等,2013),仅文献《中国重点保护野生植物资源调查》记载了广东北部有香果树分布,数量为20株,因个体数量太少,所计算的蓄积量为0,所处群落面积2 hm2(国家林业局,2009)。由于天然林的逐渐消失,加上该种的种子萌发率极低,天然更新困难,按照IUCN的评估标准,香果树可被列为“易受害”(易危,VU)等级的植物(国家林业局,2009)。
国内外迄今对香果树的研究涉及多个方面。近2年来的研究主要集中在香果树中2种新的吲哚生物碱(WU等,2013a)、三萜类和细胞毒活性类固醇(MA等,2012)、微卫星标记的发展和特征化(WU等,2013b)、资源的利用与保存(颜健等,2013;王辉等,2013;邵红琼等,2013)、空间分布格局(彭焱松等,2013)、花及胚胎发育(李利平等,2012)及地理新分布(樊卫东等,2012)等。有关香果树群落、种群、生理生态学的研究地主要分布在浙江大盘山、安徽天堂寨、江西庐山、福建武夷山、河南桐柏山、鄂东大别山、湖北九宫山和七姊妹山、湖南大围山等地,未见对广东境内香果树群落或种群的专门研究。本研究在对广东连州田心自然保护区植物资源深入研究的过程中,发现梅树冲村旁山地分布有香果树的天然种群,为了解该保护植物的种群及群落现状,对该种群的生态特征和生境特征进行研究,将有助于了解广东境内香果树最大种群的生存和发展,并丰富广东乃至全国的香果树研究成果。
1 研究地自然条件概况
成立于 2008年的广东连州田心省级自然保护区,地处北回归线以北,南岭山脉的中段南侧,属中亚热带季风气候区,气候四季分明,年均温19.5 ℃,1月均温 8.8 ℃,极端最低温-6.9 ℃;7月均温28.5 ℃,极端最高温39.8 ℃。年均降雨量1571.8 mm,82%降雨集中于3~8月。年均相对湿度81%,无霜期298.7 d。整个保护区内有珍稀濒危国家保护植物24科30属30种,早期仅记载了1株香果树(缪绅裕等,2013)。后发现梅树冲有香果树群落及其种群,用Garmin12型GPS定位地理坐标为 N25°07′44′,E112°25′59′,海拔 760 m。
2 材料与方法
2.1 气温与大气相对湿度
2013年,在连州田心梅树冲香果树种群所处生境内,用 HOBO-U23-002型温湿度记录仪(美国Onset公司出品)于每日 00:00:00、03:00:00、06:00:00、09:00:00、12:00:00、15:00:00、18:00:00、21:00:00自动记录该地温度和湿度,监测为期1年。
2.2 光照强度
于不同日期同一时刻(16:00),用TES专业级照度计测定群落内和空旷地的光照强度,每点测 6次,求平均值和标准差,以观测香果树对光照强度的适应性。
2.3 土壤理化特性
在种群调查地内每个小样方的中央位置各取 1个样,充分混合阴干、过20目筛后,用TPY-6A浙江托普土壤养分速测仪测定土壤pH值、铵态氮、速效磷、有效钾和有机质含量,土水质量比为 1∶2.5。
2.4 植物叶片生理生态特性
在植物生长季节,定期用日本产SPAD-502测定仪测定活体叶片的叶绿度(SPAD),每次测定颜色相近的成熟叶片 30片,求平均值和标准差。于不同日期同一时刻(16:15),用北京雅欣理仪科技有限公司生产的Yaxin-1102型便携式光合蒸腾仪测定叶片的光合速率、蒸腾速率等,每次测定不同个体的4~6片叶子,求平均值和标准差。
2.5 群落物种多样性
按样方调查法对群落内的乔木层植物进行每木调查,测定乔木层所有个体的胸径和高度。有香果树分布的林地总面积仅为20 m×20 m,将其分隔为4个10 m×10 m的小样方,以计算乔木层各物种的重要值。同时记录样地内的灌木、草本和藤本植物的种类。
2.6 香果树种群数量与结构
用数显的游标卡尺测定每株香果树的基径大小,同时用测高器测定植株高度,绘制香果树种群的所有个体基径的径级结构图和高度结构图。
3 结果与分析
3.1 气温与相对湿度
2013年的大气温湿度自动监测结果表明,香果树生境的年平均温度为 17.315 ℃,其年变化情况见图 1。图 1中显示,1年中的日均极端最低温-0.509 ℃(2月9日,其中0:00:00为所有数据中的最低值-0.902℃),极端最高气温 29.6 ℃(7月 13日,其中15:00:00为33.574 ℃)。年平均相对湿度为84.316%,日平均最大相对湿度为100.0%(出现于多日),最小 30.630%(9月 15日,其中 24:00为所有数据的最低值29.850%)。日均气温波动最大为1~4月份,而大气相对湿度波动最大的是9月份。
气温和相对湿度的月平均值的变化情况见图2,图 2中月均气温变幅为 7.442 ℃(12月)~27.502 ℃(7月);月均相对湿度变幅为82.358%(9月)~95.735%(5月)。图2显示,气温与相对湿度之间的线性相关关系未达显著水平(相关系数仅0.084),显然气温的变化规律较明显(夏季的6~8月最高,冬季的12~2月最低),而相对湿度的变化幅度相对较小,无明显变化规律(9月和3月较低,4~5月和 11月较高)。气温的季节变化为冬季9.644 ℃<春季(3~5月)的13.039 ℃<秋季(9~11月)17.429 ℃<夏季26.099 ℃。相对湿度的季节变化为秋季89.750%<冬季91.959%<春季92.597%<夏季93.857%,这与其他同纬度地区通常以冬季较干燥有明显区别。
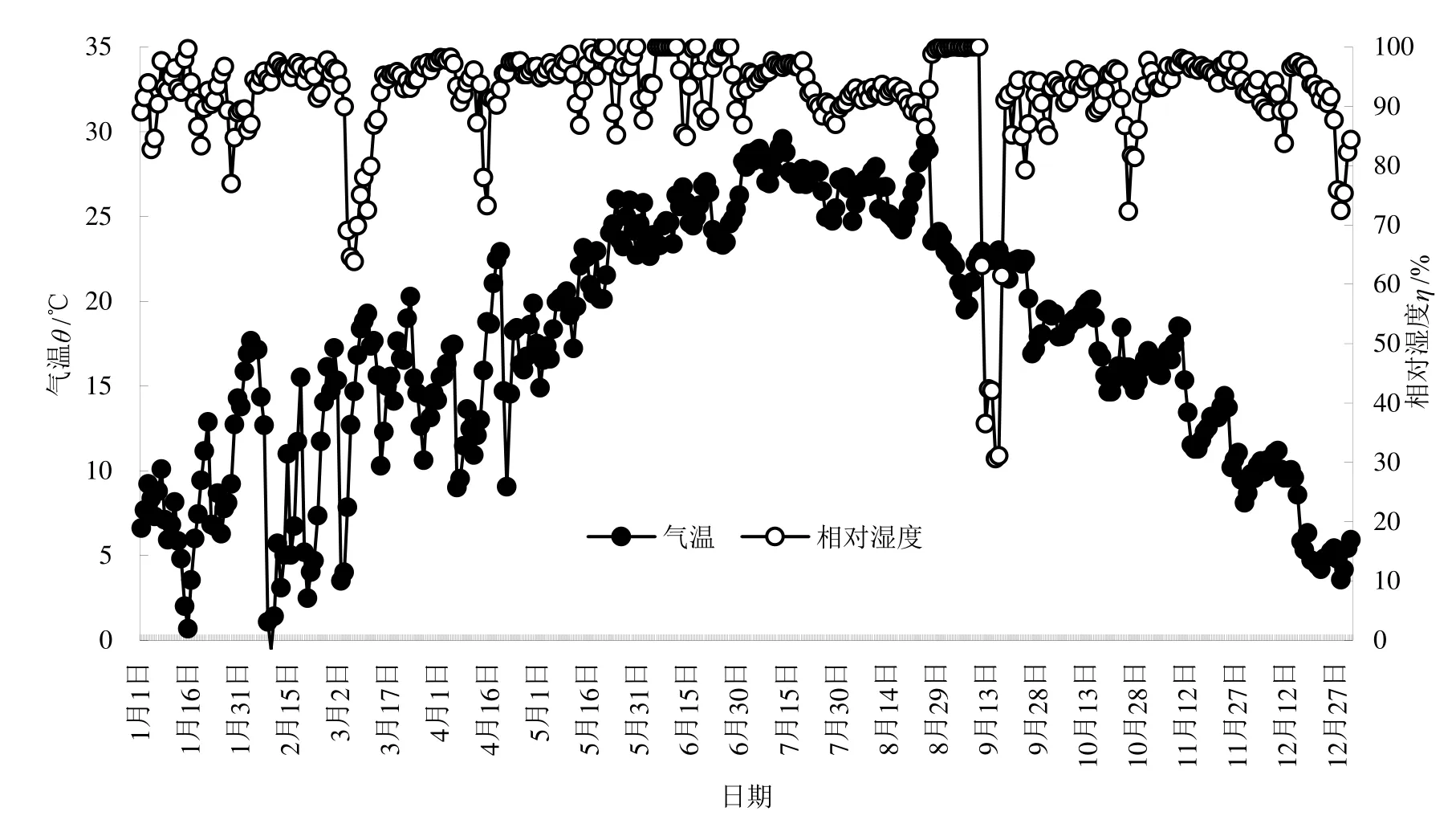
图1 香果树群落气温与相对湿度2013年变化Fig.1 Annual Changes of Air Temperature and Relative Humidity in 2013 at the Emmenopterys henryi Community
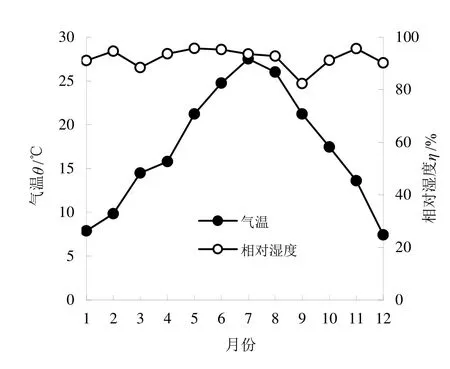
图2 2013年香果树群落气温和相对湿度月变化Fig.2 Monthly Changes of Air Temperature and Relative Humidity in 2013 at the Emmenopterys henryi Community
3.2 光照强度
不同测定日同一时刻(16:00)所测定的植物叶面和旷地光照强度见表1。由表1可知,不同天气条件叶面光照强度与旷地光照强度的比值相差较大,阴天条件下仅 3.59%,多云条件下可达11%~12%。
3.3 土壤理化特性
所测定的土壤理化特性见表2。表2中,土壤pH为 5.64,与所处的中亚热带偏酸性的山地黄壤性质偏酸性一致。
3.4 植物叶片生理生态特性
3.4.1 叶绿度
4月30日、7月16日和8月26日所测叶片的SPAD值分别为36.76±3.56、39.57±4.31和42.68±4.97。3次测定的结果之间无显著差异,但有略微上升的趋势,由于香果树为落叶大乔木,该结果可能是随着叶片的生长,与其叶绿素的合成积累有关。
3.4.2 净光合速率与蒸腾速率
不同季节测定日所测得的净光合速率、蒸腾速率等指标的测定结果见表3。由表3可知,3次测定的生态因子中,大气和叶内的CO2质量浓度、大气和叶内的相对湿度均相差不大,4月底的气温、叶温、光合有效辐射与7月和8月的差异较大,导致4月份的净光合速率和蒸腾速率与7、8月份的差异较大,其中净光合速率在4月的测定值显著小于7月和8月的测定值,而蒸腾速率则显著大于7月和8月测定值。7月与8月测定值之间无显著差异,7月的略高,可能与光合有效辐射较大有关。
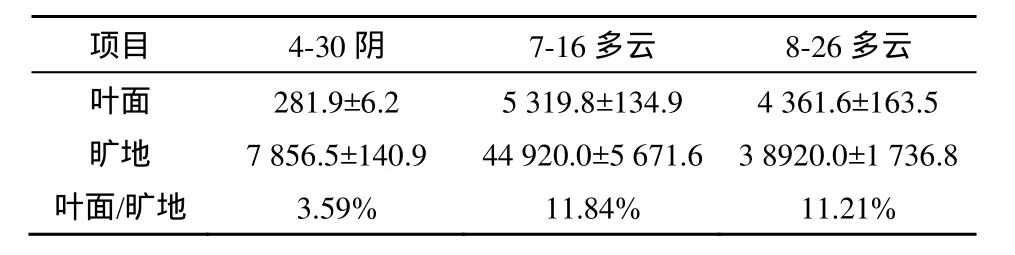
表1 香果树群落内叶面和旷地光照强度(Lx)Table 1 Light Density of Forest Opening and Above the Leaf of Emmenopterys henryi Community

表2 香果树种群土壤理化特性Table 2 Physiochemical Characteristics of Soil at Emmenopterys henryi Population
2.5 群落物种组成
根据样方调查结果,所计算的乔木层物种重要值见表4。表4中,400 m2的样地中,乔木层共有11科17种植物,以壳斗科4种植物最多,樟科3种次之,体现出亚热带地带性植被——常绿阔叶林的主要特征,种类组成上除了壳斗科和樟科等主要常绿木本植物外,该调查区还包含了较多的落叶树种,如香果树、伯乐树(Bretschneidera sinensis)、君迁子(Diospyros lotus)等。除了香果树为国家Ⅱ级重点保护野生植物外,调查区还有Ⅰ级保护植物和珍稀濒危植物各 1种,分别为伯乐树和银鹊树(Tapiscia sinensis)。因此该群落在植被代表性、种类多样性和稀有性方面均具有重要价值。

表3 香果树叶片的净光合速率和蒸腾速率Table 3 Leaf Net Photosynthetic Rate and Transpiration Rate of Emmenopterys henryi

表4 香果树群落乔木层物种及其重要值Table 4 Species and Their Important Values of Arbor tree in Emmenopterys henryi Community
森林群落的郁闭度达90%,林下灌木草本层植物种类较稀少。其中,灌木种类有9种,包括红锥(Castanopsis hystrix)、网脉山龙眼(Helicia reticulata)、鸭公树(Neolitsea chuii)、白背叶(Mallotus apelta)和八角枫(Alangium chinense)的幼苗,以及茶竿竹(Pseudosasa amabilis)、金粟兰(Chloranthus spicatus)、黄花倒水莲(Polygala fallax)、柘树(Cudrania tricuspidata);草本植物有6种,分别是九头狮子草(Peristrophe japonica)、傅氏凤尾蕨(Pteris fauriei)、黑紫藜芦(Veratrum japonicum)、楼梯草(Elatostema involucratum)、魔芋(Amorphophallus rivieri)、岭南瘤足蕨(Plagiogyria subadnata);藤本仅见楠藤(Mussaenda erosa)1种。本群落中共记录到植物 33种,其中乔木层17种占总种数1/2以上。
2.6 香果树种群数量与结构
香果树种群按基径每间隔 0.2 cm所绘制的径级结构图见图3。图3中,种群共含有97个个体,基径范围为0.50~0.69 cm的个体数最多,有20株;基径大于4.70 cm的个体仅2株,其中包括1株基径达135.05 cm的大树(胸径117.88 cm)。该大树于 2008年初被南岭地区的冰冻灾害所压垮,树高15 m以上部分已枯死。推测梅树冲香果树种群可能由此大树种子繁衍而成。从图3看,由于幼苗数量远超过成熟植株数,故目前该种群处于增长模式。但由于母树已深受冰灾影响,短期内无法正常结实,且未见基径小于0.30 cm的植株,种群更新将在一定年限后受到阻碍,故应加强保护。
按植株高度以33 cm为1个单位划分香果树种群,得梅树冲香果树个体的高度结构如图4所示,其中高度为0.20~2.03 m的香果树单株有77株,占总数的79.38%。图4中的图像与图3有一定的相似性,体现出植株的基径生长与高度生长有一定的相关性。
综合图3和图4,若按常规的划分标准对乔木层数目胸径的径级结构进行划分,高度<33.0 cm为Ⅰ级幼苗,胸径<2.5 cm为Ⅱ级幼苗,胸径2.5~7.4 cm之间为小树,胸径7.5~22.4 cm为中树,胸径≥22.5 cm大树的标准划分,则梅树冲的香果树种群中,有Ⅰ级幼苗11株(占11.34%),Ⅱ级幼苗76株(占78.35%),小树9株(占9.28%),中树0株,大树 1株(1.03%),整个种群整体上呈增长趋势,但因能产生种子的母树受冰灾严重破坏,无法产生新的繁殖体,且Ⅰ级幼苗数量少于Ⅱ级幼苗,故该种群发展到一定阶段将趋于衰退。

图4 香果树种群高度结构Fig.4 Height Structure of Emmenopterys henryi Population
4 讨论与结论
4.1 讨论
作为广东香果树的自然分布地,连州田心自然保护区梅树冲有得天独厚的气候条件,也是迄今在广东境内发现的香果树最大种群分布地。2013年测定的该境地气温在-0.902~33.574 ℃,年平均17.315 ℃(略高于浙江多地香果树生境平均气温的12.3~16.2 ℃(康华靖等,2007b);相对湿度29.850%~100.000%,平均84.316%;土壤为山地黄壤,pH5.64,符合其自然分布区年均温18~22 ℃,相对湿度 70%~85%,土壤 pH5~6(国家林业局,2009;傅立国,1992)的条件。本研究中,香果树生境土壤的有机质质量分数为 1.95%、速效磷为7.47 mg·kg-1和有效钾为 110.5 mg·kg-1,均低于大小兴安岭林区的蒙古栎(Quercus mongolica)林的有机质 3.29%、26.73 mg·kg-1和 212.96 mg·kg-1(王芳等,2013),有效钾质量分数也低于浙江大盘山香果树林土壤总钾质量分数(7.02%)(陈子林等,2009),但略高于我国黄棕壤、红壤和赤红壤有效钾质量分数(<100 mg·kg-1)(杨振明等,1999)。本研究中,香果树生境土壤的有机质、速效磷和有效钾含量相对较低,但种群的绝大多数植株生长状况良好,生活力较强,即使主干受到损毁也能萌生出较多枝条,表明香果树对生境土壤的要求并不十分严格。此研究结果可为该香果树的迁地保护提供科学依据。
香果树为偏阳性树种,但幼苗和10 a树龄以内的幼树能耐荫蔽,通常散生于以壳斗科为主的常绿阔叶林、常绿与落叶阔叶混交林中,一般树龄在30 a以上的壮龄树才能开花结实(国家林业局,2009;傅立国,1992)。本研究的香果树种群,也同样散生于以壳斗科为主的森林中(乃梅树冲村村头基本未被人类破坏的风水林),幼苗耐阴,其所处的内部环境光照仅有空旷地的11.5%左右。有研究表明,高光强(5000 lx)对香果树幼苗生长极其不利,而低(1000 lx)和中光强(2200 lx)下生长较好(刘鹏等,2008)。本研究在多云条件下测定的群落内部叶面光照强度4300~5300 lx,接近上述研究的强光,但实际上,5000 lx的光照强度仅为阳光直射条件下50000 lx的1/10左右,对于植物光合作用而言亦非强光。
湖北九宫山的香果树群落样方中(400 m2)有18种植物,其中乔木7种,主要为山茶科、壳斗科植物,灌木10种、草本层3种,藤本仅有扶芳藤(Euonymus fortunei)1种(徐小玉等,2002)。湖南大围山香果树群落样方中(400 m2)有35种植物,其中乔木8种,灌木11种,草本16种(刘成一等,2011)。浙江大盘山香果树群落主要伴生种为樟科和山茶科植物,生活型以高位芽植物占绝对优势(陈子林等,2007)。田心梅树冲香果树群落400 m2样地有植物33种,其中乔木层17种(以壳斗科、樟科植物最多,但缺乏山茶科植物),灌木9种,草本植物6种,藤本植物1种,与湖北九宫山、湖南大围山、浙江大盘山比较,有一定的相似性,但各具特点。
浙江境内多个生境(康华靖等,2007a;康华靖等,2007b)、安徽天堂寨(杨开军等,2007)的香果树种群长期演替均呈衰退趋势。种群的空间分布格局主要为集群分布(康华靖等,2007b),香果树的萌生能力较强,尤其是当母树受到破坏时,将产生大量的萌生苗,来维持种群的生存,使得幼树阶段呈聚集分布(康华靖等,2007a;康华靖等,2007b)。而萌条多见于倒木、树桩、根际等处。梅树冲的香果树种群繁衍于1株母树,其树龄在30 a以上,但2008年遭受严重的冰灾,导致无法继续开花结实。群落中有不萌生苗,目前的种群Ⅰ级幼苗数量少于Ⅱ级幼苗数量,属增长种群,但因多数个体年龄低于10 a,故在20 a内难以有新的幼苗加入,种群将逐渐趋于衰老。因此,加强对连州田心自然保护区香果树种群的就地保护是非常必要的。同时由于该保护区的地理位置和气候条件适宜,应开展更深入的研究,包括对该种群每1个体的长期跟踪定位观测,以及从国内其它地方获得种子后进行人工繁殖和迁地保护等。此外, 应加强对当地村民的宣传保护教育,以防止人为对香果树种群产生破坏。
4.2 结论
(1)广东连州田心自然保护区拥有迄今发现的广东境内的香果树最大天然种群,为研究国家Ⅱ级重点保护植物香果树的种群动态提供了宝贵的自然资源。本研究通过对2013年该生境的气温、相对湿度、土壤和森林内部的光照条件等生长因子的监测调查,认为该地是香果树自然分布的适宜分布地。
(2)梅树冲的香果树种群目前虽为增长种群,但因受到冰冻灾害的严重影响,尤其是能开花结实的成体植株严重受损,种群将难以在短期内得到更新,并将逐渐演变为衰退种群,为此应加强对该种群的保护和长期定位观测。
致谢:广东连州田心省级自然保护区的欧阳玉光、桂广来;广州大学的王厚麟、黄金玲、梁桂滔、梁劲章、陈炽军、关博等同志参加了野外调查工作,美国Louisiana州立大学R.D.DeLaune教授和A.Jugsujinda博士为英文摘要润色,特致谢意。
MA Wei-Wei,ZHAI Sheng-Nan,ZHANG Yong-Hua,et al. 2012.Development and characterization of microsatellite markers forEmmenopterys henryi(Rubiaceae), a rare tree from China[J].American Journal of Botany, 99(4):E179-E181.
WU Xing-De,HE Juan,LI Xing-Yao,et al. 2013a. Triterpenoids and steroids with cytotoxic activity fromEmmenopterys henryi[J].Planta Medica,79 (14):1356-1361.
WU Xing-De,WANG Lei, HE Juan, et al. 2013b. Two New indole alkaloids fromEmmenopteryshenryi[J].Helvetica Chimica Acta,96(12):2207-2213.
陈子林,康华靖,刘鹏,等. 2007. 大盘山自然保护区香果树群落结构特征[J].云南植物研究, 29(4):461-466.
陈子林,康华靖,周楠,等. 2009. 香果树主要金属元素分布及其与土壤养分之间的关系[J].中国野生植物资源, 28(1):30-36.
樊卫东,沈茂才,刘军,等. 2012. 香果树属——秦岭北坡茜草科一新分布属[J].西北植物学报, 32(4):819-820.
傅立国. 1992.中国植物红皮书——稀有濒危植物(第一册)[M].北京:科学出版社:568-569.
国家林业局. 2009. 中国重点保护野生植物资源调查[M].北京:中国林业出版社:153-154.
康华靖,陈子林,刘鹏,等. 2007a. 大盘山自然保护区香果树种群结构与分布格局[J].生态学报, 27(1):389-396.
康华靖,刘鹏,陈子林,等. 2007b. 不同生境香果树种群的径级结构与分布格局[J].林业科学, 43(12):22-27.
李利平,李争艳,王玉兵,等. 2012. 香果树花及胚胎发育的细胞学研究[J].植物研究, 32(6):646-650.
刘成一,廖建华,陈月华,等. 2011. 湖南大围山香果树群落特征及物种多样性分析[J].中南林业科技大学学报, 31(11):110-113.
刘鹏,康华靖,张志详,等. 2008. 香果树(Emmenopterys henryi)幼苗生长特性和叶绿素荧光对不同光强的响应[J].生态学报, 28(11):5656-5664.
缪绅裕,黄金玲,唐志信,等. 2013. 广东连州田心保护区珍稀濒危保护植物研究[J].广州大学学报:自然科学版, 12(5):29-34.
缪绅裕,王厚麟,黄金玲,等. 2008. 粤北和粤东北若干珍稀濒危野生植物的种群特征[M].热带亚热带植物学报, 16(5):397-406.
彭少麟,陈万成. 2003. 广东珍稀濒危植物[M].北京:科学出版社:66.
彭焱松,张晓波,桂忠明,等. 2013. 庐山香果树毛竹混交林空间格局研究[J].广西植物, 33(4):502-507.
邵红琼,樊春华,杨子慧. 2013. 香果树种质资源保存库建设要点[J].黑龙江农业科学, (1):160-161.
王芳,黄玫,孙希华,等. 2013. 大小兴安岭林区不同林型土壤养分综合评价[J].水土保持通报, 33(1):182-187.
王辉,陈丽文. 2013. 豫南山区珍稀树种香果树的开发利用[J].林业实用技术, (4):46-48.
邢福武,陈红锋,王发国,等. 2012. 南岭植物物种多样性编目[M].武汉:华中科技大学出版社:164.
徐小玉,姚崇怀,潘俊. 2002. 湖北九宫山香果树群落结构特征研究[J].西南林学院学报, 22(1):5-8.
颜健,邱頔,陆璐,等. 2013. 高含游离脂肪酸的香果树籽油制备生物柴油的方法[J].植物分类与资源学报, 35(1):89-94.
杨开军,张小平,张中信,等. 2007. 安徽天堂寨保护植物香果树群落现状分析[J].植物资源与环境学报, 16(1):79-80.
杨振明,周文佐,鲍士旦,等. 1999. 我国主要土壤供钾能力的综合评价[J].土壤学报, 36(3):377-386.
