甘蔗实生苗根系性状的遗传分析
2014-07-02赵丽萍刘家勇覃伟昝逢刚杨昆赵培方赵俊吴才文
赵丽萍,刘家勇*,覃伟,昝逢刚,杨昆,赵培方,赵俊,吴才文
(1.云南省农业科学院甘蔗研究所,云南 开远 661699;2.云南省甘蔗遗传改良重点试验室,云南 开远 661699)
甘蔗实生苗根系性状的遗传分析
赵丽萍1,2,刘家勇1,2*,覃伟1,2,昝逢刚1,2,杨昆1,2,赵培方1,2,赵俊1,2,吴才文1,2
(1.云南省农业科学院甘蔗研究所,云南 开远 661699;2.云南省甘蔗遗传改良重点试验室,云南 开远 661699)
采用桶栽方式,研究甘蔗实生苗根系性状的遗传效应。结果表明:根长、根交叉数受母本、父本和组合的影响显著,根表面积受母本和父本的影响显著,根系干重受组合的影响显著;亲本及组合的选配方式对根系遗传力大小的贡献率表现为母本、组合、父本依次减小;9个根系性状中,根交叉数、根长、根表面积、根尖数、根干重、根体积的遗传力均大于60%;在所选用的亲本中,用新台糖22号、桂糖92–66作母本,用CP72–1210、川糖 89–103作父本,其后代的根系性状表现较好;相关分析结果表明,根长、根表面积、根直径可作为甘蔗根系育种的选择指标。
甘蔗;根系;实生苗;遗传
1 材料与方法
1.1 材 料
试验杂交后代来源于桂糖92–66×CP72–1210、ROC22×川糖89–103、粤糖00–236×闽糖96–1409、粤糖 96–86×云蔗 02–588、粤糖 00–236×云蔗02–588(分别记为A、B、C、D、E)5个杂交组合。
1.2 试验设计
试验于2012年3—9月在云南省农业科学院甘蔗研究所进行。为了尽量减少清洗时对根系的损伤,桶栽试验基质采用珍珠岩和河沙。基质下层为珍珠岩,上层为河沙,珍珠岩与河沙的体积比3∶1。于实生苗长至4~5叶时期(2012年6月23日)从苗床挑选长势一致的苗移栽至桶内。试验采用3重复设计,即每组合种植9桶(3桶为1个重复,每桶5 株)。植株成活后开始浇营养液,每周1次。营养液选用Hoagland’s(霍格兰氏)经典配方。植株于2012 年9月24日收获。
1.3 测定指标及方法
将植株从培养桶中完整取出,用自来水将根系冲洗干净。将清洗干净的实生苗完整根系从植株上分离下来,用吸水纸吸干其表面水分,称取鲜重后再将根系按单根一条条分开,不重叠,不交叉,并排于专用根盘。利用WinRHIZO根系专用大幅面透视扫描仪扫描根系。把扫描好的图片按群体株系号和重复分类保存在不同文件夹,用根系专用WinRHIZO软件进行分析,计算出根长、根表面积、根系平均直径、根体积、根尖数、根分枝数、根交叉数。把扫描完的根系放入烘箱,105 ℃杀青0.5 h,70 ℃下烘干至恒重,测定其干重。
1.4 数据统计分析
采用R软件和SPSS 18.0进行数据分析。
式中:Tel(SEe)为以设备为主的有效工作时间函数;SEe表示该组织设备中第l台设备的有效工作时间;γel表示第l台设备的工作效率;tel为第l台设备的额定工作时间。
2 结果与分析
2.1 根系性状的差异性分析
由表1可见,根长、根交叉数均受母本、父本和组合的影响显著;根表面积在母本和父本间均表现出显著差异,说明根表面积受母本和父本的影响显著;根系干重在组合间表现出显著差异,说明根系干重受组合的影响显著。

表1 根系性状的方差分析结果Table 1 Variance analysis of root characters
2.2 根系性状的遗传力分析
由表2可见,亲本及组合的选配方式对根系遗传力的贡献率表现为母本(68.30%)、组合(64.1%)、父本(62.5%)依次减小,说明母本对后代的影响大,组合对后代的影响其次,父本的影响相对较小。从后代各性状的平均遗传力看,根交叉数、根长、根表面积、根尖数、根干重、根体积、根直径、根分枝数、根鲜重的遗传力呈依次减小的变化趋势,根交叉数、根长、根表面积的遗传力均大于70%,根分枝数和根鲜重的遗传力相对较低,分别为52.5% 和46.99%,根交叉数、根长、根表面积、根尖数、根干重、根体积、根直径6个性状的遗传力均在55%以上,遗传力较高,能稳定遗传,因此,在选择亲本和配组合时应重视对这6个性状的选择。
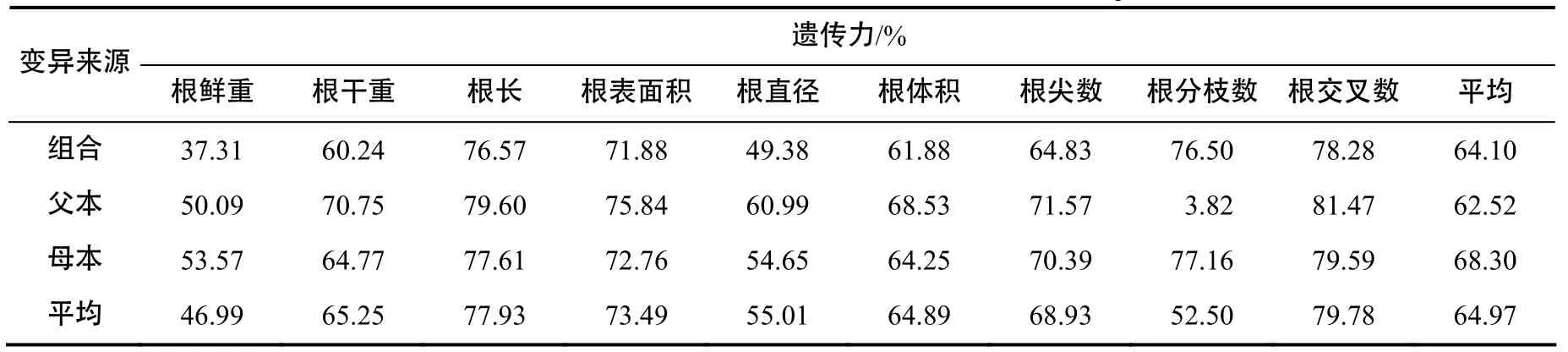
表2 父本、母本及组合根系性状的遗传力Table 2 The contribution rate of male,female and their combination on major root characters
2.3 根系性状的配合力
2.3.1 母本根系性状的一般配合力效应
由表3可知,在所选用的4个母本材料中,桂糖92–66和 ROC22后代根系性状的表现优良,尤其是用ROC22作母本,其后代根系性状的GCA均表现为正效应,根鲜重、根干重、根长和根尖数的GCA较高,表现突出,这些根系优良性状容易传递给后代;桂糖92–66根鲜重、根干重表现出负效应,但根表面积、根直径、根体积、根分枝数出现正效应,且GCA较高;用粤糖00–236作母本的后代根系性状表现较差,GCA均呈现出负效应;粤糖96–86仅有根直径和根尖数 GCA呈现出正效应,其余均为负效应,因此,从根系性状上进行选择时,ROC22、桂糖92–66后代根系性状的综合表现较好,可考虑作母本使用,粤糖 00–236后代根系性状的表现差,不适合作母本使用。
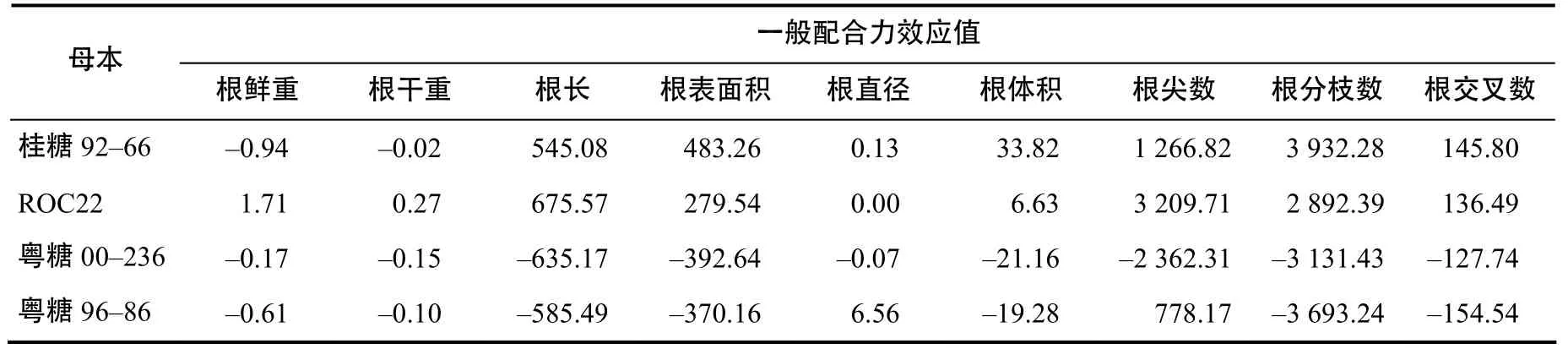
表3 母本根系性状的一般配合力效应值Table 3 Relative effect values of GCA of female root characters
2.3.2 父本根系性状的一般配合力效应
由表4可知,用CP72–1210、川糖89–103作父本的后代根系综合性状表现较好,CP72–1210根鲜重、根干重的GCA均呈现出负效应,其余均为正效应,根表面积、根分枝数、根交叉数的GCA效应值均高于其他父本;川糖89–103作父本的后代根系综合性状表现均呈现出正效应,后代根鲜重、干重、根长、根尖数的GCA效应值较高;闽糖96–1409后代根系性状的GCA效应值较低,且均为负效应;云蔗 02–588后代根直径和根体积均呈现出正效应,GCA效应值较高,分别为34.51和1 167.73,其余均为负效应,因此,从所选用的父本中,川糖89–103、CP72–1210后代根系性状的综合表现较好,可考虑作父本使用;闽糖96–1409后代根系性状的表现差,不适合作父本使用。
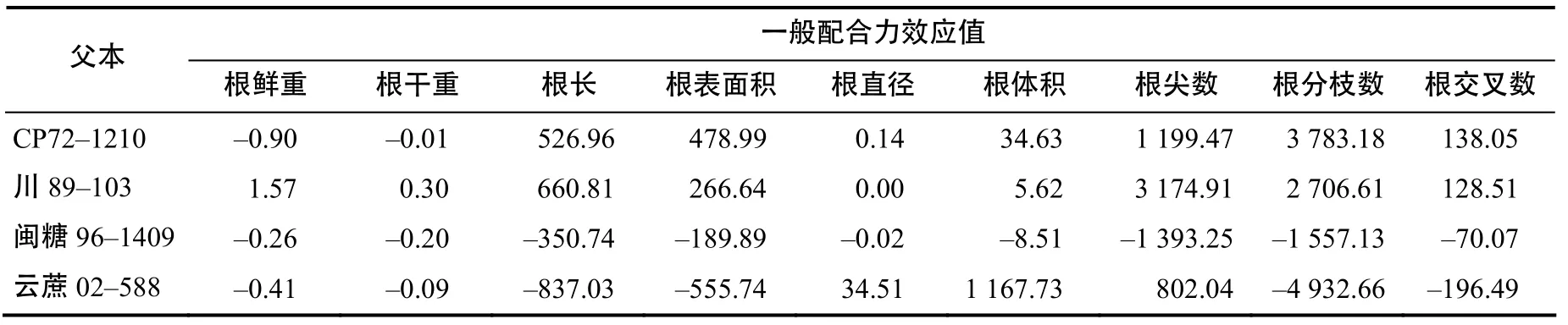
表4 父本根系性状的一般配合力效应值Table 4 Relative effect values of GCA of male root characters
2.3.3 根系性状的特殊配合力效应
由表5可知,综合表现好的为组合A和组合B,组合A根鲜重的SCA为负效应,其余均为正效应,且根表面积、根直径、根体积、根分枝数、根交叉数SCA均比其余4个组合的高,分别为533.84、0.13、35.19、4 377.8、163.41。组合B的SCA为正效应,根鲜重、根干重、根长、根尖数的SCA均比其余4个组合的高,分别为1.20、0.27、763.85、3 262.62。组合C、D、E的SCA值较小,组合C根系直径和组合D根系鲜重的SCA为正效应,其余根系性状均为负效应。
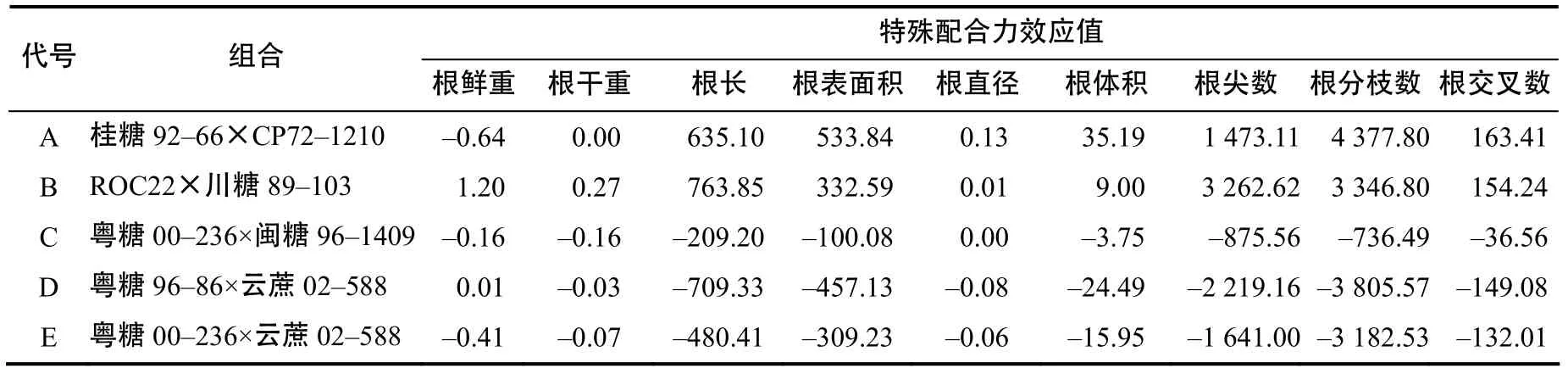
表5 杂交组合根系性状的特殊配合力效应值Table 5 Relative effect values of SCA of combination root characters
2.3.4 根系性状间的相关分析
由表6可见,根系性状间存在不同程度的相关性,根长与根表面积、根尖数、根分枝数、根交叉数的相关性达极显著水平;根表面积与根体积、根分枝数、根交叉数的相关性达极显著水平;根直径与根体积的相关性达极显著水平,说明通过提高根长可有效改良根表面积、根尖数、根分枝数、根交叉数;提高根表面积可有效改良根体积、根分枝数、根交叉数;提高根直径可有效改良根体积,因此,根长、根表面积、根直径可以作为甘蔗根系育种选择的指标。
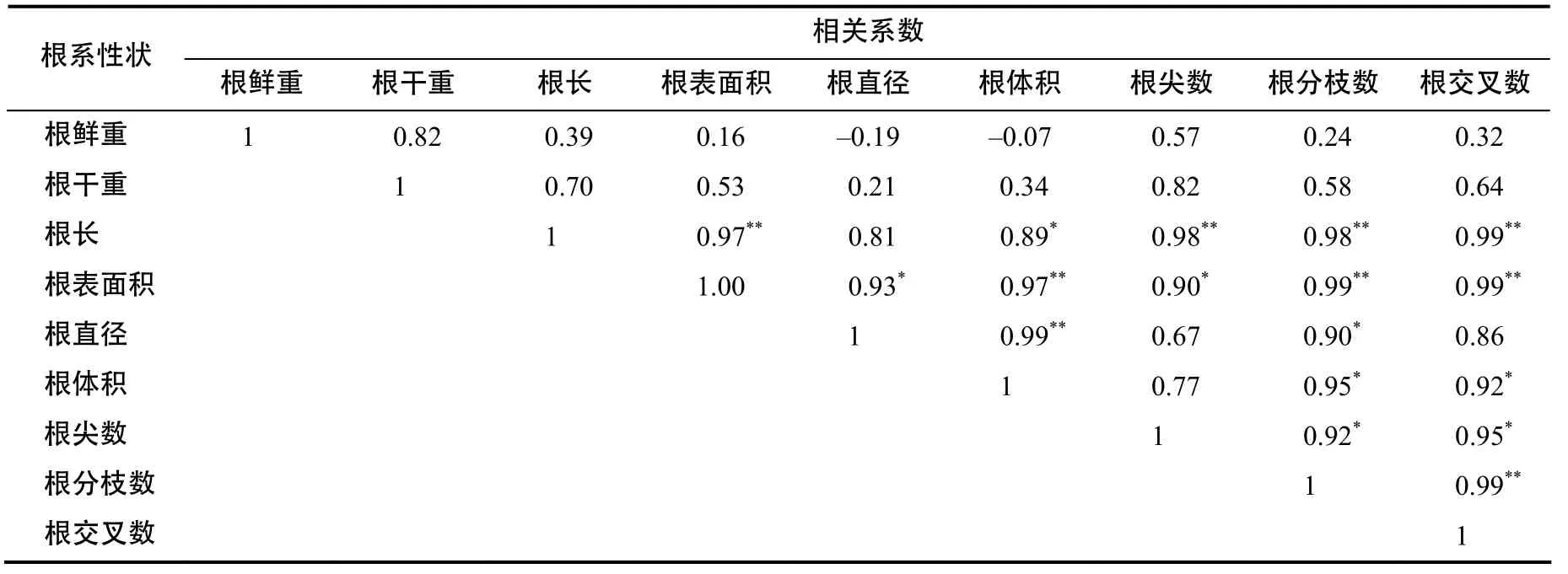
表6 根系性状表型的相关系数Table 6 Correlation coefficients of root characters
3 结论与讨论
不同组合实生苗的根长、根交叉数均受母本、父本和组合的影响;根表面积受母本和父本的影响显著;根系干重受组合的影响显著。遗传分析结果表明,亲本及组合的选配方式对根系遗传力的贡献率母本为68.30%、组合为64.1%、父本为62.5%,后代各性状的平均遗传力表现为根交叉数(79.78%)、根长(77.93%)、根表面积(73.49%)、根尖数(68.93%)、根干重(65.25%)、根体积(64.89%)、根直径(55.01%)、根分枝数(52.5%)、根鲜重(46.99%依次减小。用 ROC22、桂糖 92–66作母本,川糖89–103、CP72–1210作父本的后代根系性状综合表现较好。桂糖 92–66×CP72–1210和 ROC22×川糖89–103的后代根系性状SCA综合表现较好。在根系性状的9个指标中,根长、根表面积、根直径可以作为甘蔗根系育种选择的指标。
近年来,大量研究结果表明作物品种基因型及环境条件对根系的生长及分布有明显影响[20]。国内外对水稻[21–23]、玉米[24–26]、大豆[27–29]、马尾松[30]、
甘蓝[31]等根系性状遗传变异的研究也表明根系性状不同程度地存在遗传差异。根长、根表面积、根体积、根干重、根数量、根冠比、根直径、根毛长度、主根长度、根密度和根长密度等都具有较高的遗传力[32–33]。本研究中根交叉数、根长、根表面积、根尖数、根干重、根体积的遗传力均大于60%,这些根系性状能稳定遗传,说明运用这些性状进行早代选择有效。在所选用的亲本中,用ROC22、桂糖92–66作母本,用CP72–1210、川糖89–103作父本的根系性状具有较高的配合力,在甘蔗根系育种中具有重要的利用价值。用ROC22作母本的配合力较高,优良性状遗传给后代的能力强[34]。利用根系育种有可能首先取得突破[35]。目前,根系形态性状、营养元素吸收、耐盐、耐涝等相关基因被成功克隆[36–39],借助分子标记技术和应用基因工程方法进行根系性状研究和根系改良具有广阔的前景。
[1] 梁永书,占小登,高志强,等.超级稻协优9308衍生群体根系与地上部重要农艺性状的相关性[J].作物学报,2011(10):1711–1723.
[2] 史晓江,贺德先,詹克慧,等.作物根系性状的遗传学研究进展[J].河南农业科学,2006(1):12–16.
[3] Saseendran S A,Nielsen D C,Lyon D J,et al.Modeling responses of dryland spring triticale,proso millet and foxtail,millet to initial soil water in the high plains[J]. Field Crops Research,2009,113:48–63.
[4] Smith D M,Inman-Bamber N G,Thorburn P J.Growth and function of the sugarcane root system[J].Field Crops Research,2005,92:169–183.
[5] 刘坤,陈新平,张福锁.不同灌溉策略下冬小麦根系的分布与水分养分的空间有效性[J].土壤学报,2003,40(5):697–703.
[6] 苗果园,高志强,张云亭,等.水肥对小麦根系整体影响及其与地上部相关的研究[J].作物学报,2002,28(4):445–450.
[7] 朱德峰,林贤青,曹卫星.水稻深层根系对生长和产量的影响[J].中国农业科学,2001,34(4):429–432.
[8] Manschadi A M,Hammer G L,Christopher J T,et al. Genotypic variation in seedling root architectural traits and implications for drought adaptation in wheat (Triticum aestivum L.)[J].Plant and Soil,2008,303:115–129.
[9] 李鲁华,陈树宾,秦莉,等.不同土壤水分条件下春小麦品种根系功能效率的研究[J].中国农业科学,2002,35(7):867–871.
[10] Zhu J M,Mickelson S M,Kaeppler S M,et al.Detection of quantitative trait loci for seminal root traits in maize (Zea mays L.)seedlings grown under differential phosphorus levels[J].Theoretical and Applied Genetics,2006,113:1–10.
[11] 朱维琴,吴良欢,陶勤南.作物根系对干旱胁迫逆境的适应性研究进展[J].土壤与环境,2002,11(4):430–433.
[12] 蓝立斌,陈超君,米超,等.不同施氮方式对甘蔗生理生化性状的影响[J].南方农业学报,2011,42(1):26–29.
[13] 徐林,黄海荣,黄玉溢,等.地下滴灌条件下甘蔗根系和蔗地土壤速效养分分布规律的研究[J].广东农业科学,2011(1):78–80.
[14] 黄杏,杨丽涛,李杨瑞,等.固氮菌接种对甘蔗根系生理特性的影响[J].广西农业科学,2009,40(3):233–237.
[15] 周桂,邹成林,丘立杭,等.聚乙二醇胁迫下甘蔗根系蛋白质差异表达图谱分析[J].华中农业大学学报,2010,29(4):408–412.
[16] 周桂,丘立杭,邹成林,等.水分胁迫对甘蔗根系蛋白质差异表达的影响[J].西南农业学报,2010,23(5):1455–1459.
[17] 王威豪,罗永明,梁强,等.水分胁迫下甘蔗根系蛋白质和核酸对外源乙烯的响应[J].西南农业学报,2008,21(6):1579–1582.
[18] Aitken K S,Jackson P A,McIntyre C L.Quantitative trait loci identified for sugar related traits in a sugarcane (Saccharum spp.) cultivar×Saccharum officinarum population[J].Theor ApplGenet,2006,112:1306–1317.
[19] Patricia Battie Laclau,Jean-Paul Laclau.Growth of the whole root system for a plant crop of sugarcane underrainfed and irrigated environments in Brazi[J].Field Crops Research,2009,114:351–360.
[20] 王法宏,王旭清,刘素英,等.根系分布与作物产量关系的研究进展[J].山东农业科学,1997(4):48–51.
[21] 丁平,贺立源,李德华,等.低磷胁迫下不同水稻基因型苗期根系性状的遗传研究[J].安徽农业科学,2008,36(1):96–97.
[22] 张成良,陈大洲,肖叶青,等.东乡野生稻苗期根系形态性状耐冷性遗传分析[J].江西农业学报,2010,22( 2):1–3.
[23] 陈明明,董伟,胡中立,等.水稻苗期根系性状的数量遗传分析[J].武汉植物学研究,2000,18(5) :431–434.
[24] 陈俊意,徐莉.低磷土壤下玉米根系质量的混合遗传分析[J].西南师范大学学报:自然科学版,2009,34(1):63–66.
[25] 刘宇,于海秋,李兴涛,等.耐低钾玉米自交系主要根系性状的遗传分析[J].沈阳农业大学学报,2010,41(2):199–202.
[26] 陈俊意,蔡一林,徐军,等.玉米基因型磷效率与相对根系性状的最优回归模型及育种学启示[J].玉米科学,2008,16(6):58–61.
[27] 刘莹,盖钧镒,吕慧能,等.大豆耐旱种质鉴定和相关根系性状的遗传与QTL定位[J].遗传学报,2005,32 (8):855–863.
[28] 刘莹,盖钧镒,吕慧能.黄淮海地区大豆耐旱根系性状的遗传分析[J].华北农学报,2007,22(5):31–35.
[29] 刘莹,盖钧镒,吕慧能.黄淮海地区大豆品种苗期根系性状的遗传变异及与耐逆境胁迫的关系[J].华北农学报,2008,23(1):114–118.
[30] 杨青,张一,周志春,等.异质低磷胁迫下马尾松家系根构型和磷效率的遗传变异[J].植物生态学报,2011,35(12):1226–1235.
[31] 陈娟妮,梁颖.长江流域主要甘蓝型油菜品种苗期耐湿性鉴定[J].中国生态农业学报,2011,19(3):626–630.
[32] Caradus J R.Genetic control of phosphorus uptake and phosphorus status in plants[C]//Genetic Manipulation of Crop Plants to Ehance Integ rated Nutr ient Management in Cropping System.Patancheru,India:ICRISAT Asia Centre,1995:55– 74.
[33] 王秀全,陈光明,何丹,等.玉米根系性状的遗传及相关分析[J].西南农业学报,2002(3):24–27.
[34] 覃伟,吴才文,曾千春,等.用新台糖甘蔗品种作母本培育强宿根后代的潜力评价[J].湖南农业大学学报:自然科学版,2012,38(1):1–7.
[35] 严小龙,廖红,年海.根系生物学:原理与运用[M].北京:科学出版社,2007:73–115.
[36] Ren Z H,Gao J P,Li L G,et al.A rice quantitative trait locus for salt tolerance encodes a sodium transporter[J].Nat Genet,2005,37:1141−1146.
[37] Inukai Y,Miwa M,Nagato Y,et al. Characterization of rice mutants deficient in the formation of crown roots[J].Breed Sci,2001,51:123−129.
[38] Yao S G,Mushika J,Taketa S,et al.The short-root mutation srt5 defines a sugar-mediated root growth in rice (Oryza sativaL.)[J].Plant Sci,2004,167:49−54.
[39] Zhao Y,Hu Y F,Dai M G,et al.The WUSCHEL-related homeobox gene WOX11 is required to activate shootborne crown root development in rice[J].Plant Cell,2009,21:736−748.
责任编辑:王赛群
英文编辑:王 库
Heredity analysis of sugarcane seedling root characters
ZHAO Li-ping1,2,LIU Jia-yong1,2*,QIN Wei1,2,ZAN Feng-gang1,2,YANG Kun1,2,ZHAO Pei-fang1,2,ZHAO Jun1,2,WU Cai-wen1,2
(1.Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences, Kaiyuan, Yunnan 661699, China; 2.Yunnan Province Key Laboratory of Sugarcane Genetic Improvement, Kaiyuan, Yunnan 661699, China)
The root inherited characters of sugarcane seedlings were conducted by a potted trial. The results indicated that root length, root crossings were significantly affected by male, female and their combination; root surface areas were significantly affected by male and female, and root dry weights were significantly affected by their combination. The contribution sequence of heritability on offspring decreased as female, their combination and male in turn. The heritability of root crossings, root length, root surface areas, number of root tips, root dry weight and root volume were all greater than 60%. The root characters were much better for these springs by taken ROC22 and GT92–66 with female and CP72–1210 and CT 89–103 with male. The correlation analysis showed that the root length, root surface areas and root diameter might be used as the index for selecting roots in sugarcane breeding.
sugarcane; roots; seedling; heredity
S566.1
A
1007−1032(2014)05−0470−06
10.13331/j.cnki.jhau.2014.05.004
投稿网址:http://www.hunau.net/qks
2014–06–21
国家现代农业产业技术体系建设专项基金项目(CARS–20–1–1);云南省重点新产品开发计划项目(2011BB005,2012BB014)
赵丽萍(1981—),女,云南沾益人,助理研究员,主要从事甘蔗育种及配套技术研究,zlp_alyma@126.com;*通信作者,lljjyy1976@163.com
