重庆地区早三叠世蠕虫状灰岩特征及成因
2014-06-27时志强王湘君王艳艳
程 明,时志强,王湘君,王艳艳,金 鑫,段 雄,崔 磊
(1.油气藏地质及开发工程国家重点实验室(成都理工大学),成都 610059;2.大港油田勘探开发研究院,天津 300280)
重庆地区早三叠世蠕虫状灰岩特征及成因
程 明1,时志强1,王湘君2,王艳艳1,金 鑫1,段 雄1,崔 磊1
(1.油气藏地质及开发工程国家重点实验室(成都理工大学),成都 610059;2.大港油田勘探开发研究院,天津 300280)
重庆地区早三叠世沉积中广泛发育一种“错时相”灰岩——蠕虫状灰岩。通过对重庆地区十余条典型剖面飞仙关组及嘉陵江组进行野外观察、采样及室内薄片鉴定、扫描电镜研究,认为蠕虫状灰岩由形似蠕虫的“蠕体”和浅色基质组成,其中蠕体形态多样,常呈条带状、粒状、不规则状发育于基质中,蠕体的主要成分为方解石(质量分数高达95%以上),基质中相对含有较多黏土,但方解石的质量分数依然可达到85%~90%。蠕虫状灰岩中几乎见不到大的生物化石,镜下可观察到黄铁矿颗粒及藻类痕迹。按形态可将蠕虫状灰岩分为顺层线纹状、断续点状和异形杂乱状3类,其形成主要和菌藻类活动有关,并且在其形成过程中风暴作用影响明显,后生动物(如腕足、腹足及双壳类)的扰动会抑制蠕虫状灰岩的发育,因此蠕虫状灰岩的成因是(低等)生物与水动力共同作用的结果。二叠纪末-三叠纪初特提斯洋海水温度高,大气CO2含量极高而O2含量却极低,相应的古海洋是停滞的、缺氧的并且伴随有频繁的风暴,这些都为蠕虫状灰岩的形成提供了条件。
蠕虫状灰岩;早三叠世;风暴作用;特提斯洋
古生代-中生代之交爆发的生物大灭绝是显生宙地质历史上最大也是最具有毁灭性的一次事件,随之发生了(也可能是引起生物大灭绝的原因)全球古环境、古气候的急剧变换[1-3]。之后的生物复苏却经历了约5 Ma的时间。在如此残酷的环境影响下生物的复苏曲折而艰难,但是这种环境却造就了种属低级、单一的海洋微生物和细菌的存活[4-6]。而正是在这个时期,全球沉积体系发生了巨大的改变,不仅出现了“礁缺失”、“煤沉积缺失”和“层状硅质岩缺失”[7-9],而且在浅海环境中沉积了大量特殊的碳酸盐沉积及相关构造,如扁平砾屑灰岩、潮下皱纹构造、微生物岩、海底碳酸盐胶结扇、泥质条带泥晶灰岩和蠕虫状灰岩等。因为这些特殊的沉积及相关构造是在寒武纪或者更早的前寒武海洋环境中广泛发育的,而在奥陶纪之后退缩到少量缺乏后生动物的高压力环境中,如潟湖、深海等。而现在这些沉积及构造在寒武纪之后的正常浅海环境中再次出现被认为是“在时间和空间上发生了错误”,故称之为“非正常”沉积或“错时相沉积”[10]。
在重庆地区的整个异常碳酸盐岩沉积中,蠕虫状灰岩是发育非常普遍并且是极具代表性的“错时相”灰岩之一,部分异常碳酸盐岩及构造反映的环境状态是一方面与二叠纪末的后生动物大灭绝直接相关,另一方面也是三叠纪初生物复苏迟缓的重要原因[4]。对蠕虫状灰岩的研究也是对于“错时相”沉积研究的一种很好补充,更是有助于对二叠纪末、三叠纪初古气候、古海洋等相关问题的探讨。关于蠕虫状灰岩的成因,诸多学者提出不同观点,主流观点有机械成因学说[11-13]、生物扰动学说[10,11,14-16]、化学凝聚学说[10,15]、超盐性学说[17]及复合成因学说[10,18]等,但还没有哪个观点被普遍认同。
笔者对重庆地区诸多剖面进行了野外勘察,在石柱鱼池、丰都雪玉洞、南川水江、万盛平山、万盛关坝、綦江藻渡、北碚文星场、北碚水井坪、北碚芹菜田、重庆歌乐山和重庆凉风垭等10余条剖面均发现有蠕虫状灰岩(图1)。通过野外剖面分层描述、样品采集、薄片镜下观察、样品酸处理以及扫面电镜等实验对蠕虫状灰岩进行了宏观及微观的描述和研究,以期对于蠕虫状灰岩的成因及其所反映的古环境有更为深入的了解。
1 下三叠统蠕虫状灰岩特征
1.1 蠕虫状灰岩的分布
蠕虫状灰岩,又名蠕粒状灰岩、虫管灰岩、虫迹灰岩等,是一种描述性称谓,因其含有诸多形似“蠕虫”的粒状体而得名,到现在为止还没有哪位学者的定义及分类为大家所认可。重庆地区蠕虫

图1 重庆地区下三叠统蠕虫状灰岩剖面分布图Fig.1 Localities of the sections for the Lower Triassic vermicular limestone in the Chongqing area
状灰岩广泛发育于下三叠统飞仙关组和嘉陵江组中(图2);据张杰(2010)统计,蠕虫状灰岩在下扬子地区也广泛发育[19]。
1.2 典型剖面蠕虫状灰岩特征
1.2.1 万盛关坝剖面
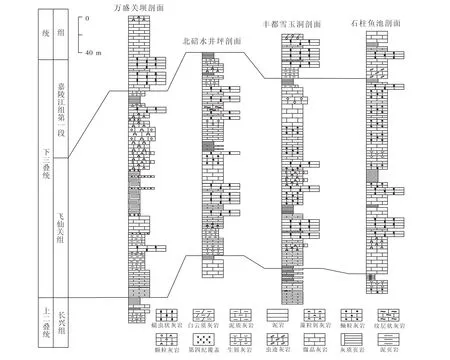
图2 重庆地区部分剖面下三叠统飞仙关组-嘉陵江组第一段对比图Fig.2 Correlation column of the Lower Triassic Feixianguan Formation and Jianlingjiang Formation from several sections in the Chongqing area
剖面位于重庆市万盛区关坝镇,大地构造上属于上扬子地台中部,次级单元为川黔南北向构造带望乡台背斜[20]。区内下三叠统大多出露良好,由下往上依次为飞仙关组和嘉陵江组,蠕虫状灰岩在两者之中均有大量分布。因为此处的蠕虫状灰岩极具研究价值,所以笔者对其进行了较为细致的实测分层,蠕虫状灰岩大多出露于飞仙关组和嘉陵江组第一段中。
蠕虫状灰岩顾名思义是由形似蠕虫状的“蠕体”和基质两部分构成,其中“蠕体”颜色较深,基质颜色较浅(图3-B)。“蠕体”以不同的表现形态呈现于基质中,如:连续条纹状、断续条纹状、斑点状、粒状、透镜状和椭球状等。在万盛关坝剖面,“蠕体”呈现出多种形态,但多数表现为连续或断续条带状。
万盛关坝剖面所见的蠕虫状灰岩具有以下特征:(1)蠕虫状灰岩风化面上“蠕体”与基质区分明显,而新鲜面上不明显。(2)蠕虫状灰岩成层沉积,且同一层中的“蠕体”形态基本一致(为顺层状或粒状)。(3)蠕虫状灰岩中的“蠕体”垂向上的变换有时不是渐变式的,如连续条带状向上或者向下不是渐变为断续条带状,而是直接突变为异形杂乱状(图3-B)。(4)蠕虫状灰岩沉积较厚但层间常夹不含“蠕体”的微晶灰岩(图3-B)。(5)蠕虫状灰岩中仅见极少量的介壳类生物化石。
1.2.2 重庆歌乐山剖面

图3 重庆地区下三叠统蠕虫状灰岩宏观照片Fig.3 Photos showing macro characters of the Lower Triassic vermicular limestone in the Chongqing area
重庆歌乐山剖面飞三段一套连续产出的地层中观察到岩层底部发育连续条带状蠕虫状灰岩,中下部鲕粒灰岩中含有大量介壳,中上部发育微晶灰岩,顶部岩层继续发育连续条带状蠕虫状灰岩。蠕虫状灰岩与生物介壳灰岩或介屑鲕粒灰岩交替产出,其中生物介壳及鲕粒发育的层位不含蠕虫状灰岩,非常直观地表现出当时蠕虫状灰岩的形成与生物相关,这为讨论蠕虫状灰岩的成因提供了较好的证据(图3-F)。
1.2.3 北碚水井坪剖面
该剖面蠕虫状灰岩产于飞三段,基质和蠕体多呈缝合线接触,蠕体边缘凹凸不平,部分有压实、压溶的现象;野外露头上,蠕体多顺层呈断续线状分布,延伸较远。蠕虫状灰岩缺乏生物扰动的标志,为海底静水环境正常沉积的产物。见少量粒泥结构的蠕虫状灰岩,含腕足、腹足、瓣鳃化石碎片,蠕体减少,其展布形态也缺少了连续和断续线纹状,大都杂乱零星点缀于生屑周围。
2 蠕虫状灰岩分类
通过对重庆地区十余条剖面的观察,蠕虫状灰岩中的蠕体具塑性变形特征,常呈连续或断续条带状、透镜状、斑点状、粒状和椭球状等(图3-A,C),蠕体大小不一,大的长轴可达数十厘米,小的仅有几毫米。这些大小不一、形态各异且颜色较深的蠕体发育于颜色较浅的基质中,其中蠕体部分可占到整个岩石的20%~60%(面积分数)之间。将蠕体和基质分别与盐酸反应,发现蠕体与酸的反应较基质剧烈,且基质中会有不参与反应的矿物剩余。显微镜下观察,蠕体与基质的接触类型有渐变式、缝合线式、锯齿式等,甚至有的没有明显的接触界线。镜下蠕体色浅透明,基质色暗,镜下表现恰恰与野外露头相反。蠕体中的方解石结晶较好且晶体颗粒较大;基质的主要成分也是方解石,但结晶程度没有蠕体好,晶体颗粒较小且含有较多黏土(图4-C)。据黄思静(1984)、张杰(2010)所做X射线衍射实验,结果显示蠕体的主要矿物成分为方解石(质量分数>95%)和少量石英;基质成分较复杂,但仍以方解石为主(质量分数为85%~90%),其与酸反应残余的黏土矿物为伊利石、石英及少量绿泥石[7,10]。
野外剖面观察中并未在蠕虫状灰岩发育层段发现大的生物化石。镜下基质中可见极少量种属单一的生物介壳且为广盐度生物,还可见到些许黄铁矿分散于其中(图4-A,B);藻类痕迹可见,以团状、絮状分布其中,大小不同且形态各异(图4-D)。在蠕虫状灰岩中可见球状钙质微生物化石(图4-E,F),这种微生物钙球在早三叠世叠层石中有报道[5,21,22]。

图4 重庆地区下三叠统蠕虫状灰岩微观照片Fig.4 Microphotos of the Lower Triassic vermicular limestone in the Chongqing area
综合重庆地区各剖面地层中蠕虫状灰岩的形态,参考朱洪发等(1992)的分类方案[11],大致可归为3类:(1)顺层线纹状:蠕体顺层线状连接且与基质层平行相间(图3-D);(2)断续点状:蠕体顺层排列,并各自孤立呈点状,但蠕体顺层大体能相互相连成层(图3-A,C);(3)异形杂乱状:蠕体形态多样,有弯曲状、分叉状、椭球状,外部边缘不圆滑,杂乱且不均匀分布成斑状、点状(图3-B,I)。
3 蠕虫状灰岩成因
3.1 蠕虫状灰岩研究现状
对于蠕虫状灰岩的成因,目前以机械破碎、生物扰动、化学凝聚、超盐性和几种学说的综合最为盛行。
机械成因学说:朱洪发等(1992)认为部分蠕虫状灰岩(如断续点状蠕虫状灰岩)是上部斜坡带沉积物由于重力滑塌原因引起的[11]。钱守荣(1995,1996)认为蠕虫状灰岩是风暴浊流作用的产物,实际上是一种特殊的风暴岩[12,13]。
生物成因学说:许耀明(1982)认为蠕虫状灰岩是在一种环境较闭塞、海水流动较缓及盐度较高的碳酸盐潮滩上由生物扰动作用形成的[14]。黄思静(1984)认为生物潜穴扰动灰岩(即蠕虫状灰岩)是由生物潜穴、扰动作用形成的[15]。朱洪发等(1992)认为异形杂乱状蠕虫状灰岩是受生物作用(扰动、钻穴、觅食等)引起的[11]。赵小明等(2008)认为柱状蠕虫状灰岩中的蠕体是生物潜管和扰动斑点,具生物成因,它的出现意味着底栖生物群落及其活动能力的加强及底层环境的显著改善[10]。宋逢林等(2010)推测蠕虫状灰岩是由微生物主导生态系统活动的产物[16]。
超盐性学说:Piotr等(2010)通过对比反复出现于岩层内的蠕虫状灰岩、白云质灰岩和白云岩中生物遗迹的种类和数量,发现从蠕虫状灰岩到白云质灰岩是逐渐减少的,而到了白云岩则几近消失,进而认为这种表现很有可能是盐度升高而导致生物减少引起的[17]。岩层中重复出现的蠕虫状灰岩反映盐度波动的反复,而盐度的波动是由气候控制的,这与二叠纪末、三叠纪初反复无常的气候是吻合的。
化学凝聚学说:黄思静(1984)认为蠕粒状灰泥岩是由沉积作用及化学凝聚作用形成的[15]。赵小明等(2008)认为粒状蠕虫状灰岩可能是化学凝聚或微生物凝聚形成的[10]。
复合成因学说:即为几种学说的综合,姜月华等(1992)认为由沉积作用及沉积分异作用所形成的条带状灰岩是前提,而后经过生物扰动、水流及压实等作用形成各种形态的蠕虫状灰岩[18]。赵小明等(2008)认为层状、似层状蠕虫状灰岩为低能环境下的成岩分异、化学和机械作用的结果,椭球状蠕虫状灰岩为机械和化学综合作用的结果[10]。
3.2 P/T过渡时期及早三叠世古海洋与古气候条件
Sun等(2012)等经过大量野外及室内的工作,通过牙形石样品,证明二叠纪末海洋表面温度高达40℃;而在陆地上温度更高,也可高达到50℃:如此高的温度已经接近海洋生物死亡及光合作用停止的致命温度。极度的高温有可能是导致二叠纪末特提斯地区古生物灭绝的直接原因,并且也极可能是生态系统恢复、生物复苏迟缓和煤缺失的原因。不稳定的、极高的温度很可能控制着古环境复苏和恢复的进程[6]。
Isozaki(1994),Wignall(2002)和Kidder(2004)等认为二叠纪末三叠纪初的古海洋是停滞的、缺氧的,且这一停滞缺氧的海洋环境也延伸到了浅海地区[23-25];同时,海水的翻转作用使海底碳酸盐过饱和的贫氧海水上涌更加剧了海洋表面缺氧的现状[26]。诸多学者通过各种实验和模拟,认为诱使早三叠世海洋循环停滞的原因是赤道到极地温度梯度的减小,同时停滞的海洋循环及当时缺O2富CO2的大气可能加剧了古海洋缺氧的现状[27-29]。因为火山剧烈的喷发及同期的温室效应促使大洋底部水溶性有毒气体CO、CO2、H2S和CH4等大量释放,进而又一次加剧了生物集群绝灭的规模及速度[30]。
二叠纪末、三叠纪初大气中的CO2含量极高,而O2含量却处于比较低的水平[29]。同期早三叠世海相地层中广泛发育的纹层状沉积物、少见的底栖生物扰动现象和大量发育的层状黄铁矿等,都指示了早三叠世停滞和缺氧的古海洋环境[31]。与此同时,下三叠统中同样发育了大量指示环境剧烈变化的非正常碳酸盐岩沉积及构造,如角砾状灰岩(即SBB:Shangsi Breccia Bed)、扁平砾屑灰岩等。关于它们的成因普遍认为是风暴或海啸作用的结果[32-34],该时期全球变暖可能导致早三叠世飓风和冬季风暴发生的强度和频率增加[25,35,36]。童金南和殷鸿福(2009)认为部分异常碳酸盐岩反映的环境状态一方面与二叠纪末后生动物大灭绝直接相关,另一方面也是三叠纪初生物复苏迟缓的重要原因[4]。因此,早三叠世晚期异常碳酸盐沉积及相关构造的逐渐减少和消失预示着环境开始逐步改善。万盛平山剖面可以观察到蠕虫状灰岩发育层段之上有虫迹灰岩的出现(图3-H),说明生物活动逐渐开始增强,此时已开始摆脱早三叠世生物活动稀少的局面。因此,蠕虫状灰岩的消失可以作为早三叠世生物开始复苏和环境逐步改善的一个重要标志[19]。
3.3 蠕虫状灰岩成因讨论
重庆北碚水井坪、歌乐山、凉风垭剖面和万盛关坝剖面观察到蠕虫状灰岩常呈现出从顺层线纹状→断续点状→异形杂乱状的垂向沉积变化特征,可以推测出其所反映的水动力条件应该是由弱及强的。断续点状蠕虫状灰岩应是顺层线纹状灰岩受到稍强水动力条件改造的结果;如若水动力条件继续增强,此时断续点状蠕虫状灰岩将继续转变为异形杂乱状蠕虫状灰岩,因此,水动力条件在蠕虫状灰岩的形成过程中扮演了重要的角色。沉积作用、沉积分异作用及之后的成岩过程中所发生的差异压实等在蠕虫状灰岩的形成过程中也起到了一定的作用,如图3-C中可以观察到岩层因受差异压实作用的影响而致使上部和下部的蠕虫状灰岩转变为异形杂乱状蠕虫状灰岩,从岩层中部可以观察到其原始形态为顺层线纹状蠕虫状灰岩。
重庆地区蠕虫状灰岩的薄片中可见藻类痕迹,藻类以团状、絮状分布其中,大小不同且形态各异(图4-D)。在对蠕虫状灰岩进行的SEM实验中发现菌藻类球状微生物化石[37],此类化石在古生代的志留纪、寒武纪地层中已被发现[38]。所以笔者推测蠕虫状灰岩的成因与菌藻存在一定的联系(图4-E,F),可能和高温条件下存活的蓝藻[6]有关。在重庆歌乐山剖面连续的地层中观察到岩层底部发育顺层线纹状蠕虫状灰岩,中下部岩层产出含有大量生物介壳的鲕粒灰岩,中上部发育一套微晶灰岩(既不含蠕虫状灰岩也不含生物介壳),顶部岩层继续发育顺层线纹状蠕虫状灰岩。有意思的是,底部和顶部的顺层线纹状蠕虫状灰岩中看不到介壳,同样,中下部含有大量生物介壳的鲕粒灰岩中也观察不到蠕虫状灰岩发育的痕迹(图3-F)。据此推断,发育有生物介壳的岩层之所以不含蠕体是因为以藻类为食是腹足、双壳等生物对其大量捕食的结果,而底部和顶部含有顺层线纹状的蠕虫状灰岩不含生物介壳是因为以藻类为食的后生捕食生物稀少继而藻类大量繁殖的缘故。
上扬子地区早三叠世早期已受到超级季风的影响[39],因此风暴在同期蠕虫状灰岩的形成过程中也必将起到一定的作用。蠕虫状灰岩主要为生物成因;而异形杂乱状蠕虫状灰岩的成因,笔者认为是以藻类为食的生物在对其进行捕食时于其中活动的结果,或者是相对低能环境中发育的平坦藻席经风暴作用后再沉积的结果。钱守荣(1995,1996)认为蠕虫状灰岩的形成与风暴浊流作用有关,实际上是一种特殊类型的风暴岩[12,13]。在丰都雪玉洞剖面飞仙关组第三段上部的异形杂乱状蠕虫状灰岩中观察到砾屑分布其中,砾屑颗粒较大且磨圆极差(图3-E),因此风暴作用在川东南重庆地区蠕虫状灰岩的形成过程中也是曾经起到过很大作用的。蠕虫状灰岩的沉积模式如图5所示。海底蓝细菌-藻席是松软的,容易受到风暴的影响,故蠕虫状灰岩普遍发育于正常浪基面之下、强风暴浪基面之上,可与斜坡地带发育的风暴成因的扁平砾屑灰岩共生[40](图5-A,见于广元上寺剖面)。在一般风暴浪可以影响到的地方,蠕虫状灰岩因受到频繁风暴浪的扰动破坏形成异形杂乱状蠕虫状灰岩(图5-B)。强烈风暴浪基面之上、一般风暴浪基面之下,即受到低频率强烈风暴影响而一般风暴影响不到的地方,常发育连续-断续顺层线纹状蠕虫状灰岩(图5-C)。强风暴浪基面之下,即强烈风暴影响不到的地方水体静止,沉积低能环境的纹层状、薄板状灰岩(图5-D)。在綦江藻渡、万盛关坝、万盛平山等剖面所见丰富的虫迹构造,推测为单分异度、高丰度的软体动物捕食蓝细菌-藻席时的居住迹或活动痕迹(图3-H,I)。综上所述,作者认为蠕虫状灰岩是微生物成因的,并且在其形成过程中生物扰动及外部水动力均发挥了重要作用。断续点状和异形杂乱状蠕虫状灰岩是顺层线纹状蠕虫状灰岩受到生物扰动破坏或外部较强水动力(如风暴)作用的结果。
4 结论
a.蠕虫状灰岩广泛发育于川东南重庆地区下三叠统飞仙关组和嘉陵江组中,是由形似蠕虫的“蠕体”和浅色基质组成的。其中“蠕体”形态多样,常呈条带状、粒状、不规则状发育于基质中。蠕粒的主要成分为方解石(质量分数高达95%以上),基质的成分相对含有较多的黏土,但方解石的质量分数依然可达到85%~90%。蠕虫状灰岩中几乎见不到大的生物化石,镜下可观察到黄铁矿颗粒及藻类痕迹。
b.参考朱洪发等(1992)的分类方案,根据形态将蠕虫状灰岩分为顺层线纹状蠕虫状灰岩、断续点状蠕虫状灰岩和异形杂乱状蠕虫状灰岩3类。
c.蠕虫状灰岩的成因是由微生物主导生态系统活动的产物,继而在其形成过程中伴随有生物扰动作用和其他机械作用,如水动力作用、沉积作用、沉积分异作用和风暴作用等。与此同时,其所处近乎残酷的古气候条件和古海洋环境,如致命高温、停滞的古海洋循环、剧烈风暴及缺O2富CO2的海水和极度贫乏的后生动物共同造就了各种形态蠕虫状灰岩的沉积。
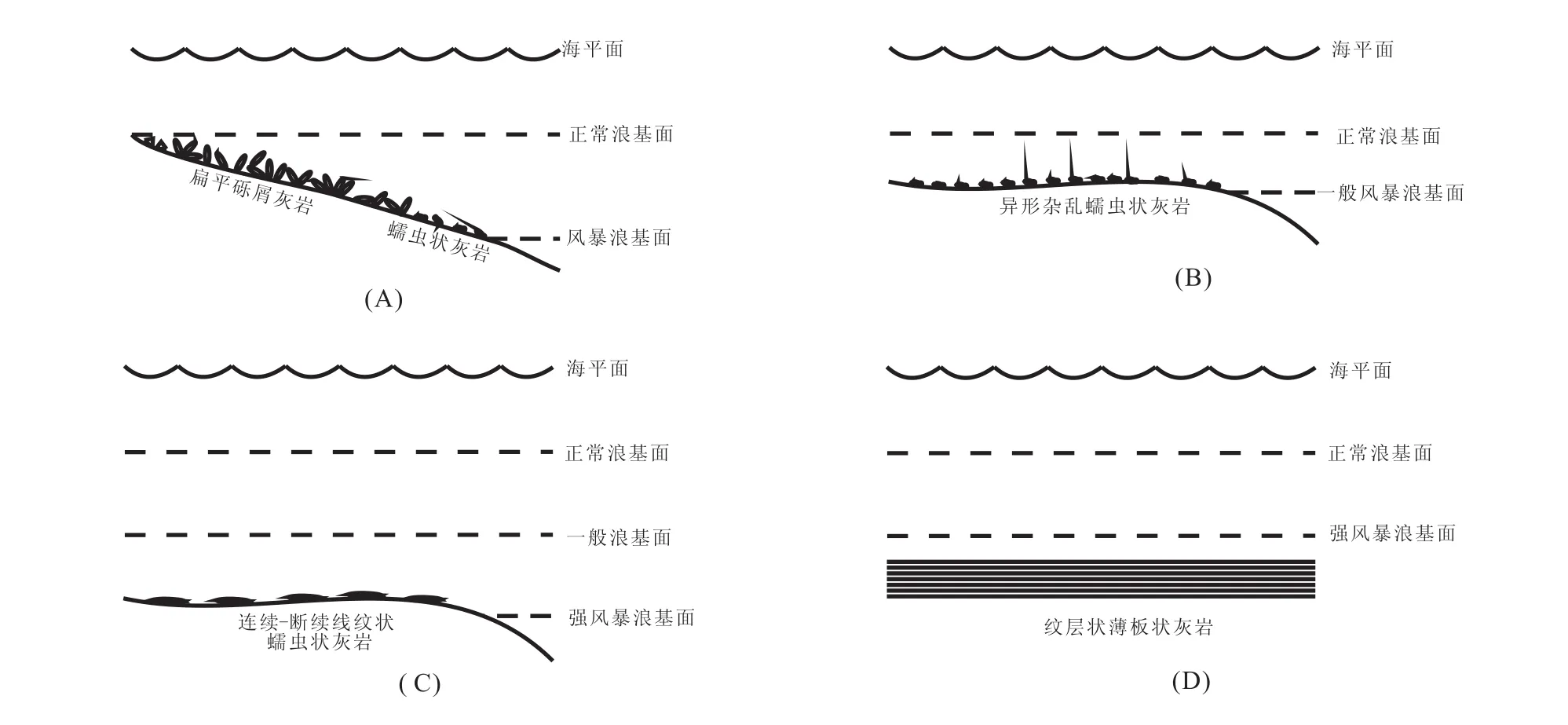
图5 重庆地区下三叠统蠕虫状灰岩沉积模式图Fig.5 Depositional model of the Lower Triassic vermicular limestone in the Chongqing area
[1]Stanley S M.Paleozoic mass extinctions:Shared patterns suggest global cooling as a common cause[J].American Journal of Science,1988,288:334-352.
[2]童金南.华南古生代末大灭绝后的生态系复苏[J].地球科学,1997,22(4):373-376.
Tong J N.The ecosystem recovery after the end-Paleozoic mass extinction in south China[J].Earth Science,1997,22(4):373-376.(In Chinese)
[3]Kozur H W.Some aspects of the Permian-Triassic Boundary(PTB)and of the possible causes for the biotic crisis around this boundary[J].Palaeogeography,Palaeoclmiatology,Palaeoecology,1998,143: 227-272.
[4]童金南,殷鸿福.早三叠世生物与环境研究进展[J].古生物学报,2009,48(3):497-508.
Tong J N,Yin H F.Advance in the study of early Triassic life and environment[J].Acta Palaeontologica Sinica,2009,48(3):497-508.(In Chinese)
[5]Wang Y B,Tong J N,Wang J S,et al.Calcimicrobialite after end-Permian mass extinction in South China and its palaeo-environmental significance[J].Chinese Science Bulletin,2005,50(7):665-671.
[6]Sun Y D,Joachimski M M,Wignall P B,et al.Lethally hot temperatures during the early Triassic greenhouse[J].Science,2012,338(6105):366-370.
[7]Tong J N,Lin Q X,Shi G R.Evolution of the Permian and Triassic reef ecosystems in South China[J].Proc R Soc Victoria,1998,110:385-399.
[8]Retallack G J,Veevers J J,Morante R.Global coal gap between Permian-Triassic extinction and Middle Triassic recovery of peat forming plants[J].GSA Bull,1996,108:195-207.
[9]Baud A,Richoz S,Pruss S.The Lower Triassic anachronistic carbonate facies in space and time[J].Global and Planetary Change,2007,55:81-89.
[10]赵小明,童金南,姚华舟,等.华南早三叠世错时相沉积及其对复苏期生态系的启示[J].中国科学:D辑,2008,38(12):1564-1574.
Zhao X M,Tong J N,Yao H Z,et al.Anachronistic facies in the Lower Triassic of South China and their implications to the ecosystems during the recovery time[J].Science in China:Series D,2008,38(12):1564-1574.(In Chinese)
[11]朱洪发,王恕一.苏南、皖南三叠纪瘤状灰岩、蠕虫状灰岩的成因[J].石油实验地质,1992,14(4):454-460.
Zhu H F,Wang S Y.The origins of the Triassic nodular and vermicular limestonds in South Jiangsu-South Anhui Provinces[J].Experimental Petroleum Geology,1992,14(4):454-460.(In Chinese)
[12]钱守荣.蠕虫状灰岩成因新解[J].淮南矿业学院学报,1995,15(3):15-19.
Qian S R.A new research on the origin of the vermicular limestone[J].Journal of Huainan Minging in Stitute,1995,15(3):15-19.(In Chinese)
[13]钱守荣.蠕虫状灰岩中的同生变形构造及其成因[J].安徽地质,1996,6(1):38-41.
Qian S R.Contemporaneous deformation structures in vermicular limstone[J].Geology of Anhui,1996,6(1):38-41.(In Chinese)
[14]许耀明.江苏省宜兴县善卷洞附近的青龙灰岩及其岩相特征[J].同济大学学报,1982,7(3):43-55.
Xu Y M.The Qinglong limestones and their facies features at the Shanjuan caverns,Yixing,Jiangsu[J].Journal of Tongji University,1982,7(3):43-55.(In Chinese)
[15]黄思静.蠕虫状灰岩及其成因[J].成都地质学院学报,1984,11(3):60-68.
Huang S J.Vermicular limestone and its origin[J].Journal of Chengdu College of Geology,1984,11(3):60-68.(In Chinese)
[16]宋逢林,杨浩,王钦贤.湖南慈利礁相二叠系—三叠系界线地层及其对大灭绝事件的启示[J].华南地质与矿产,2010,15(3):51-56.
Song F L,Yang H,Wang Q X.Reef facies strata of Permian-Triassic boundary in Kangjiaping section,Cili area,Hunan Province and its implications for biological mega-deracination[J].Geology and Mineral Resources of South China,2010,15(3):51-56.(In Chinese)
[17]Piotr Jaglarz,Alfred Uchman.A hypersaline ichnoassemblage from the Middle Triassic carbonate ramp of the Tatricum domain in the Tatra Mountains,Southern Poland[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2010,292:71-81.
[18]姜月华,岳文浙,业治铮,等.蠕虫状灰岩特征和成因新探[J].矿物岩石,1992,12(1):1-7.
Jiang Y H,Yue W Z,Ye Z Z,et al.The new approach on the characteristics and origin for vermicular limestone[J].Mineralogy and Petrology,1992,12(1):1-7.(In Chinese)
[19]张杰,童金南.下扬子地区下三叠统蠕虫状灰岩及其成因[J].古地理学报,2010,12(5):535-548.Zhang J,Tong J N.Vermicular limestone of the Lower Triassic and its origin in lower Yangtze region[J].Journal of Palaeogeography,2010,12(5):535-548.(In Chinese)
[20]谭先锋,李祖兵,夏敏全.关坝地区的地质特征与地质实习教学研究[J].重庆科技学院学报:社会科学版,2012,14(3):170-172.
Tan X F,Li Z B,Xia M Q.Geology and geological research practice teaching in Guanba area[J].Journal of Chongqing University of Science and Technology(Social Sciences Edition),2012,14(3):170-172.(In Chinese)
[21]Jiang H X,Wu Y S,Cai C F.Filamentous cyanobacteria fossils and their significance in the Permian-Triassic boundary section at Laolongdong, Chongqing[J].Chinese Science Bulletin,2008,53(12):1871-1879.
[22]Wu Y S,Jiang H X,Yang W,et al.Microbialite of anoxic condition from Permian-Triassic transition in Guizhou,China[J].Science in China:Series D,2007,50(7):1040-1051.
[23]Isozaki Y.Superanoxia across the Permo-Triassic boundary:Record in accreted deep-sea pelagic chert in Japan(C)//Pangea:Global Environments and Resources.Calgary:Canadian Society of Petroleum Geologists,1994:805-812.
[24]Wignall P B,Twitchett R J.Extent duration and nature of the Permian-Triassic super anoxic event[C]//Catastrophic Events and Mass Extinctions:Impacts and Beyond.Special Paper-Geological Society of America,2002,356:395-413.
[25]Kidder D L,Worsley T R.Causes and consequences of extreme Permo-Triassic warming to globally equable climate and relation to the Permo-Triassic extinction and recovery[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2004,203:207-237.
[26]赵小明,童金南,姚华舟,等.华南下三叠统蠕虫状灰岩与生物复苏[C]//中国古生物学会第十次全国会员代表大会暨第25届学术年会论文摘要集.南京:中国古生物学会,2009:150-151.
Zhao X M,Tong J N,Yao H Z,et al.Vermicular limestone and biological recovery in the Early Triassic South China[C]//Abstract Volume,The 10thNational Congress of Palaeontological Society of China(PSC)-The 25thAnnual Conference of PSC.Nanjing:The Palaeontological Society of China,2009:150-151.(In Chinese)
[27]Wignall P B,Hallam A.Anoxia as a cause of the Permian/Triassic mass extinction:facies evidence from northern Italy and the western United States[J].Palaeogeogr,Palaeoclimatol,Palaeoecol,1992,93:21-46.
[28]Wignall P B,Twitchett R J.Oceanic anoxia and the end Permian mass extinction[J].Science,1996,272:1155-1158.
[29]Berner R A.A combined model for Phanerozoic atmospheric O2and CO2[J].Geochimicaet Cosmochimica Acta,2006,70:5653-5664.
[30]李玉成,周忠泽.华南二叠纪末缺氧海水中的有毒气体与生物集群灭绝[J].地质地球化学,2002,30(1):57-63.
Li Y C,Zhou Z Z.Massive dissociation of gas hydrate during oceanic anoxia as a cause of mass extinction at the end of Permian[J].Geology-Geochemistry,2002,30(1):57-63.(In Chinese)
[31]时志强,安红艳,伊海生,等.上扬子地区早三叠世异常碳酸盐岩的分类与特征[J].古地理学报,2011,13(1):1-10.
Shi Z Q,An H Y,Yi H S,et al.Classification and characters of the Early Triassic anomalous carbonate rocks in Upper Yangtze Area[J].Journal of Palaeogeography,2011,13(1):1-10.(In Chinese)
[32]Wignall P B,Twitchett R J.Unusual intraclastic limestones in Lower Triassic carbonates and their bearing on the aftermath of the end-Permian massextinction[J].Sedimentology,1999,46:303-316.
[33]冯增昭,王英华,李尚武.中国南方早中三叠世岩相古地理[J].地质科学,1997,32(2):212-220.
Fen Z Z,Wang Y H,Li S W.Lithofacies palaeogeography of the early and middle Triassic of south China[J].Scientia Geologica Sinica,1997,32(2):212-220.(In Chinese)
[34]赵小明,牛志军,童金南,等.早三叠世生物复苏期的特殊沉积/错时相沉积[J].沉积学报,2010,28(12):314-323.
Zhao X M,Niu Z J,Tong J N,et al.The distinctive sediments in the early Triassic recovery time“Anachronistic Facies”[J].Acta Sedimentologica Sinica,2010,28(12):314-323.(In Chinese)
[35]Barron E J.Severe storms during Earth history[J].Geological Society of America Bulletin,1989,10(5):601-612.
[36]Ito M,IshigakiA N T,Saito T.Temporal variation in the wavelength of hummocky cross-stratification:Implications for storm intensity through Mesozoic and Cenozoic[J].Geology,2001,29:87-89.
[37]陈孟莪,李菊英,陈其英.黔中晚震旦世微生物岩及其磷的富集[J].岩石学报,1999,15(3):446-452.
Chen M E,Li J Y,Chen Q Y.The Late Sinian microbiolite and its phosphorus enrichment in Central Guizhou Province[J].Acta Petrologica Sinica,1999,15(3):446-452.(In Chinese)
[38]张廷山,沈昭国,兰光志,等.四川盆地早古生代灰泥丘中的微生物及其造岩和成丘作用[J].沉积学报,2002,20(2):2-6.
Zhang T S,Shen Z G,Lan G Z,et al.Microbial fossils and their biosedimentation and buildup in Paleozoic mud mounds,Sichuan Basin[J].Acta Sedimentologica Sinica,2002,20(2):2-6.(In Chinese)
[39]时志强,曾德勇,熊兆军,等.三叠纪巨型季风在上扬子地区的沉积学记录[J].矿物岩石地球化学通报,2010,29(2):164-172.
Shi Z Q,Zeng D Y,Xiong Z J,et al.Sedimentary Records of Triassic Megamonsoon in Upper Yangtze Area[J].Bulletin of Mineralogy,Petrology and Geochemistry,2010,29(2):164-172.(In Chinese)
[40]Shi Z Q,Duan X,Wang Y Y,et al.Lower Triassic Griesbachian in Shangsi Section,NW Upper Yangtze Region,China:implications for the formation of anachronistic carbonates[J].Acta Geologica Sinica,2013,87(Z1):915-918.
Characteristics and origin of Early Triassic vermicular limestone in Chongqing area,China
CHENG Ming1,SHI Zhi-qiang1,WANG Xiang-jun2,WANG Yan-yan1,JIN Xin1,DUAN Xiong1,CUI Lei1
1.State Key Laboratory of Oil&Gas Reservoir Geology and Ex ploitation,Chengdu University of Technology,Chengdu 610059,China;
2.Exploration and Development Research Institute,Dagang Oilfield Company of PetroChina,Tianjin 300280,China
The vermicular limestone is one of the anachronistic limestone and widespread in the Lower Triassic Feixianguan Formation and Jialingjiang Formation in the Chongqing area.In terms of detailed field observation,sampling,thin-section identification,SEM,etc.for more than 10 sections,it is considered that the vermicular limestone consists of dark vermicuform bodies and light-coloured tint matrixes,among which the vermicuform bodies are in varied shapes.The main composition of the vermicuform bodies is calcite,whose content is above 95%.The matrix has more clay content,whose calcite unexpectedly reaches 85%~90%.There are few metazoan fossils that can be found in vermicular limestone,but pyrite and algae are observed under the microscope.According to the shape,the vermicular limestone is divided into three types,that is,laminar-like limestone,spot-like limestone and messy-like vermicular limestone.The formation of the vermicular limestone is related to cyanobacteria or algae activities,and the storms play a very important role in the process of its formation.The disturbance of metazoans(e.g.brachiopods,gastropods and bivalves)will inhibit the development of the vermicular limestone,thus the genesises of the vermicular limestone are biological and mechanical.The formation of the vermicular limestone is owing to such extremely grim conditions of the Early Triassic Tethys Ocean such as the high seawater temperature,high content of CO2,low content of O2,stagnant anoxia environments and frequent storms,etc.
vermicular limestone;Lower Triassic;storm;Tethys
P588.245
A
10.3969/j.issn.1671-9727.2014.03.11
1671-9727(2014)03-0347-09
2013-05-23
国家自然科学基金资助项目(40872082;41272131)
程明(1984-),男,硕士研究生,研究方向:沉积学,E-mail:cm851@163.com。
时志强(1972-),男,博士,教授,从事沉积学的教研及科研工作,E-mail:szqcdut@163.com。
