互花米草湿地碳储量及碳汇研究进展*
2014-06-05于彩芬陈鹏飞刘长安韩明辅周胜玲卢伟志
于彩芬,陈鹏飞,刘长安,张 悦,韩明辅,周胜玲,卢伟志
(1.大连海洋大学 大连 116023;2.国家海洋环境监测中心 大连 116023)
在过去的两百年中,大气中的二氧化碳(CO2)含量已经增加了30%多[1]。目前,大气中二氧化碳的增长速率比几百万年前至少快一个数量级[2],并且大气中CO2的浓度远大于地球所经历的80万年[3]。这主要是由于化石燃料燃烧、森林砍伐、其他生物碳储存的破坏和人为排放源等所导致的。据估计,大气中大约1/3的CO2被海洋吸收[4],即海洋大约可以吸收大气中26%的。目前,海洋和陆地从大气中吸收CO2的量几乎差不多[6],滨海栖息地和陆地栖息地巨大的差别在于前者可长期将碳储存在沉积物中[7]。曾有预言表明在本世纪陆地碳汇所吸收的CO2将会减少[8],而海洋从大气中吸收的CO2将会持续增加[9]。因此,海洋将会作为大气中CO2最终重要的碳汇[7]。
湿地经过长期碳积累是巨大的碳库,而且在全球碳循环中扮演着重要的碳汇角色[10]。在某些情况下,一些重要的有机碳,如可以直接或间接被无数海洋生物所利用的来自海洋植物和陆生植物中的碎屑,没有考虑进碳积累率的计算之内,我们很可能低估了长期储存在滨海植被栖息地中的碳[11]。并且,滨海植被转化了大量的碳到沉积物中,导致约有总碳量的1/2封存在海洋沉积物中,甚至可使海平面下降2%,同时,这种高埋藏速率可持续上千年[12]。所以,滨海湿地对滨海地区的高储碳率有着直接或间接的作用。
因此,虽然滨海湿地面积相对较小,但是其长期的碳封存能力是极其宝贵的。互花米草作为我国滨海湿地一种特殊植物,了解其碳储量和碳汇能力,对滨海湿地中的 “蓝碳”在应对气候变化方面的作用有着重要的意义。
1 互花米草湿地的碳储量及碳汇能力
1.1 互花米草湿地碳储量
互花米草湿地的总碳储量由两部分组成:一部分储存于植物体内,包括地上部分植物体、地下根和凋落物;另一部分储存在土壤中。互花米草的生物量从早春开始递增,在生长季内,生长速率远大于死亡速率,花期后,植物器官发育成熟,同时开始衰老。王长永等[13]通过研究米草光合作用速率季节变化及其对初级生产力的影响也证实了这一现象。因此,互花米草的这种生命活动规律决定了其生物量随时间的变化规律 (表1)。互花米草的生物量与纬度有一定的关系,国外学者比较研究了北美沼泽的互花米草揭示了其生产力随纬度增高而减少的规律[14]。另外,不同生态型的互花米草生物量也存在很大差异,钦佩等[15]对比研究了福建罗源湾海滩引种自不同地方的3个生态型互花米草的生物量,发现无论是地上部分,还是地下部分,或是总生物量都是以乔治亚生态型生物量最大,分别是2 745 g/m2、1 366 g/m2和4 111 g/m2,其次为北加罗连那生态型,分别为1 289 g/m2、1 235 g/m2和2 524 g/m2,弗罗里达生态型生物量最小,分别为972 g/m2、928 g/m2和19 00 g/m2。

表1 江苏废黄河口互花米草生物量变化[16]
研究发现我国闽江河口湿地芦苇地上生物量夏季最大,其值为1 524.8 g/m2±78.85 g/m2,互花米草在秋季最大,其值为3 037.2 g/m2±248.78 g/m2,明显高于芦苇[17];再加上大多数互花米草湿地地下部分生物量要超过地上部分[18],如此可观的互花米草地下部分生物量使互花米草总生物量也高于芦苇[17]。高建华等[19]就互花米草引种对苏北潮滩湿地总有机碳 (TOC)、总氮含量 (TN)和总磷含量(TP)分布的影响做了相关研究,发现TOC和TN的水平和垂向平均含量同样以互花米草滩为最高。对江苏王港湿地的研究表明,入侵种互花米草的生物量明显高于本地物种碱蓬,随着入侵年限的增加地上生物量和地下生物量逐渐增大,且都高于本地种碱蓬(表2)[20]。随着入侵时间推移,互花米草总生物量还要持续增长[17]。
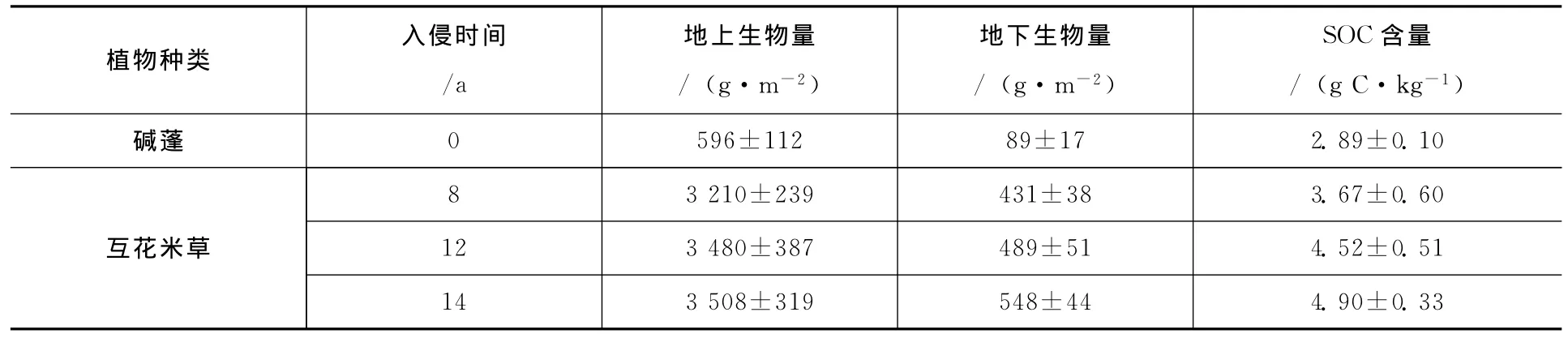
表2 江苏王港湿地互花米草和碱蓬生物学参数[26]
有机碳的积累是由于持续的初级生产力输入和较少的分解,以及碎屑的残留,互花米草高的土壤有机碳含量当然也是高的初级生产力不断通过根输入土壤所引起的。湿地沉积物有机碳含量及储量与植物种类及生物量密切相关[21],互花米草引入盐蒿湿地后,表层土壤有机碳浓度显著增加 (增量达70%),且随着互花米草生长时间延长而明显增加[22]。因此,互花米草的入侵可能加速土壤中有机碳的积累[23]。而且研究表明互花米草入侵闽江河口芦苇湿地增加了土壤碳质量分数和碳储量,芦苇和互花米草0~60 cm根际土壤有机碳储量总和分别为6 794.20 t/km2和8 231.48 t/km2,互花米草明显高于芦苇[24]。另外,土壤有机质的含量与互花米草生长年数相关程度较高,所以随着入侵年限的增加互花米草湿地有机碳储量也会持续增加[25],在江苏王港湿地的研究也发现了类似的结果 (表2)。有关学者研究了长江口湿地芦苇、海三棱藨草和互花米草生态系统的碳储量,发现芦苇湿地的碳储量为19.51 kg/m2,海三棱藨草的现存碳储量为17.07 kg/m2,互花米草的碳储量为 20.94 kg/m2,明显高于前二者[26]。据估计,112 000 hm2的互花米草每年碳固定量可增加23.9 Gg C[27],并且都固存在滨海湿地的土壤里。滨海湿地如此大的碳封存能力一部分来源于优势植被丰富的地下生物量[7],互花米草具有更高的地下生产量,而群落的土壤呼吸速率又较低,因此,互花米草作为滨海湿地的入侵种,其储碳量明显高于我国滨海湿地的土著植物芦苇、海三棱藨草和碱蓬,而且随着时间的推移,其储碳量还会不断增加。
1.2 互花米草湿地碳汇能力
所谓固碳 ,也叫碳封存,指的是以捕获碳并将其安全封存的方式来取代直接向大气中排放CO2的技术[28]。根据碳固定的方式,可将其分为人工固碳减排和植被自然固碳两类,其中植被的自然固碳功能是碳的自然封存过程,是近年来国内外碳汇研究的热点。碳汇是指生态系统中的碳平衡处于不稳定的状态,碳的输入量 (植物生产力与外界输入碳之和)大于输出量(呼吸消耗的碳和输出外界的碳),即碳积累状态。
海洋的植物生境,尤其是红树林、盐沼和海草构成了地球的蓝色碳汇,即 “蓝碳”,其在海洋碳循环中起着重要作用[12]。由于蓝色碳汇生态系统具有非常高的生物量和生产量,植物会产生大量剩余的有机碳[29],另外它们所处的地区能够截取地面物质,增加自己的剩余量,从而提高了碳埋藏率[30],故其具有强大的固碳能力。而各海洋生态系统的固碳能力大不相同,蓝色碳汇的效率也高低不一,其中每一单位面积内盐沼的碳埋藏率最高,为17.2 T C·hm-2·a-1,其次是红树林和海草 (表3)。因此,互花米草湿地作为一种盐沼类型,其碳埋藏率不容小觑。

表3 全球蓝色碳汇覆盖面积以及有机碳年埋藏率[12,29-31]
从互花米草自身的生理特性分析,作为一种C4植物,其光合作用效率和对CO2的利用率都较C3植物高,互花米草可以直接利用空气中的CO2进行光合作用,使环境中的碳含量不再成为光合作用效率的限制因素;另外,互花米草的地下部分由根和根状茎组成,深度有时可达1 m,并且具有较强的繁殖能力,所以互花米草具有较高的初级生产力,能够迅速进行生物量的生产和积累。而互花米草湿地也被认为是具有最高净初级生产力的生态系统之一[18]。
通过对比检测长江口九段沙3种植物的固碳、固氮作用,发现互花米草的固碳、固氮作用明显高于本土植物芦苇和海三林藨草。互花米草生态系统的净初级生产力为2.16 kg·m-2·a-1,芦苇和海三林藨草的净初级生产力分别为1.44 kg·m-2·a-1和0.47 kg·m-2·a-1[26],且互花米草地下部分的凋落物数量及其降解速率远低于本土植物芦苇和海三棱藨草,使互花米草对碳的净固定作用更为强劲[32]。
1.3 互花米草湿地碳储量和碳汇的关系
植物是生态系统有机碳库的重要组成部分,植物分解释放是土壤有机碳输入的主要途径之一,植物生物量则可以反映生态系统固碳能力的强弱,另外,植物的有机碳含量主要来源于空气中的CO2,植物地上部分在生长过程中吸收CO2形成光合产物,并作为生物量固定储存起来,所以植物地上部分生物量与植物体的固碳关系非常密切[33]。由于互花米草具有较长的生长季、较大的叶面积指数、较高的净光合作用速率和较大的地上、地下部分生物量,其固碳作用非常明显[26]。另外,互花米草具有极高的地下生物量,而群落土壤的呼吸速率又较低,通过茎和土壤表面释放的碳量很少,因此,互花米草可加速土壤中有机碳的积累,具有更高的固碳能力。
湿地虽然碳储量很高,但是环境的变化极易影响该生态系统中原本稳定储存的碳。我们不得不承认在开阔的滨海湿地,植物入侵对土壤有机碳的动态影响是复杂的,并且还会受到潮汐、土壤有机物例如浮游植物沉淀和土壤质量的影响。滨海湿地碳的汇源机制也较为复杂,潮来潮去的潮间带为潮间带的物种提供了间歇性的还原性环境,在这种环境中生长的互花米草能将基质中产生的甲烷等温室气体通过其通气组织排放出去[34]。互花米草高的碳储量是其碳汇和碳源相互作用的结果,如果遭到破坏互花米草高的碳储量可能成为一个高的碳源。
2 存在的问题及研究趋势
近年来,我们已经认识到互花米草湿地生态系统的碳储量很大,固碳能力很强,但是我国目前关于互花米草湿地碳储量和固碳能力的研究都是集中在如闽江河口、长江口和苏北滨海等区域,忽略了其他地区的互花米草,因此加强其他地区互花米草湿地碳储量和碳汇的研究,有助于全面和正确认识互花米草湿地生态系统的价值和意义。另外,土壤碳储量研究的野外取样深度不统一、取样方法各样,还没有专门的湿地土壤取样设备,导致不同学者对同一地区的碳储量与固碳能力评估不同[26,35-36],因此,研制用于互花米草土壤碳储量取样的仪器设备,制定统一的野外取样方法和相应的行业标准是当务之急,以提高互花米草湿地碳储量的实测精度。目前,对影响互花米草湿地碳循环的因子还未进行系统的、综合的研究,今后应加强该领域的研究。
[1]IPPC(Intergovernmental Panel on Climate Change)Climate Change 2007:The Physical Science Basis,Cambridge University Press,Cambridge,U.K.
[2]DONEY S C,SCHIMEL D S.Carbon and climate system coupling on timescales from the Precambrian to the Anthropocene.Annual Review Environment and Resources,2007,32,31-66.
[3]LUTHI D,FLOCH M L,BEREITER B,et al.High-resolution carbon dioxide concentration record 650,000-800,000 years before present,Nature,2008,453,379-382.
[4]SABINE C L,FEELY R A,GRUBER N,et al.The oceanic sink for anthropogenic CO2,Science,2004,305(5682),367-371.
[5]QUERE C L,RAUPACH M R,CANADELL J G,et al.Trends in the sources and sinks of carbon dioxide.Nature Geoscience,2009,2,831-836.
[6]BENDER M L,HO D T,HENDRICKS M B,et al.Atmospheric O2/N2changes,1993—2002:Implications for the partitioning of fossil fuel CO2sequestration,Global Biogeochemical Cycles,2005,19.
[7]The Management of Natural Coastal Carbon Sinks(2011):Carbon Sequestration by Coastal Marine Habitats:Important Missing Sinks,Emily Pidgeon.
[8]FRIEDLINGSTEIN P,COX P,BETTS R,et al.Climate-carbon cycle feedback analysis:Results from the C4MIP model intercomparison,Journal of Climate,2006,19(14),3337-3353.
[9]ORR J C,MONFRAY P,AUMONT O,et al.Simulations of historical and future anthropogenic CO2uptake from 12 global ocean models.In Sixth International Carbon Dioxide Conference,Sendai,Japan,2001,151-1054.
[10]张莉,郭志华,李志勇 .红树林湿地碳储量及碳汇研究进展[J].应用生态学报,2013,24(4):1153-1159.
[11]ODUM E P,CRUZ A A.Particulate organic detritus in a georgia salt marsh estuarine ecosystem,1967,383-388.in G,H,Lauff(ed.)Estuaries.AAAS.Washington,D.C.
[12]DUARTE C M,MIDDELBURG J J,CARACO N.Major role of marine vegetation on the oceanic carbon cycle,Biogeosciences,2005,2:1-8.
[13]王长永,钟崇信,钦佩 .米草光合作用速率季节变化及其对初级生产力的影响[J].农村生态环境,1994,10(4):14-17.
[14]TURNER R E.Geographic variation in salt marsh macrophyte production:a review.Contributions in Marine Science,1976,20:47-68.
[15]钦佩,经美德,谢民 .福建罗源湾海滩三个生态型互花米草群落生物量的比较[C]//南京大学学报编委会 .南京大学学报—米草研究论文集,南京:南京大学学报编委会,1985:222-236.
[16]钦佩,谢明,王长永 .米草的应用研究[M].北京:海洋出版社,1992,41-45.
[17]张林海,曾从盛,仝川 .闽江河口湿地芦苇和互花米草生物量季节动态研究[J].亚热带资源与环境学报,2008,3(2):25-33.
[18]关道明,刘长安,左平,等 .中国滨海湿地米草盐沼生态系统与管理[M].北京:海洋出版社,2009.
[19]高建华,杨桂山,欧维新 .互花米草引种对苏北潮滩湿地TOC、TN和TP分布的影响[J].地理研究,2007,2(4):799-807.
[20]ZHANG Y H,DING W X,LUO J F,et al.Changes in soil organic carbon dynamics in an Eastern Chinese coastal wetland following invasion by a C4plantSpartinaalterniflora.Soil Biology & Biochemistry,2010,42:1712-1720.
[21]贾瑞霞,仝川,王维奇,等 .闽江河口盐沼湿地沉积物有机碳含量及储量特征[J].湿地科学,2008,6(4):492-499.
[22]张耀鸿,张富存,周晓东,等 .互花米草对苏北滨海湿地表土有机碳更新的影响[J].中国环境科学,2011,31(2):271-276.
[23]姜丽芬 .互花米草(Spartinaalterniflora)入侵对长江河口湿地生态系统生产过程的影响:入侵种与土著种的比较研究[D].上海:复旦大学,2005.
[24]王宝霞,曾从盛,陈丹,等 .互花米草入侵对闽江河口芦苇湿地土壤有机碳的影响[J].中国水土保持科学,2010,8(2):114-118.
[25]沈永明,刘咏梅,陈全站 .互花米草盐沼土壤有机质分布特征[J].海洋通报,2003,22(6):43-48.
[26]LIAO C Z,LUO Y Q,JIANG L F,et al.Invasion ofSpartinaalternifloraenhanced ecosystem carbon and nitrogen stocks in the Yangtze estuary,China.Ecosystems,2007,10:1351-1361.
[27]AN S Q,GU B H,ZHOU C F,et al.Spartina alterniflorainvasion in China:implications for invasive species management and future research,Weed Research,2007,47:183-191.
[28]李新宇,唐海萍 .陆地植被的固碳功能与适用于碳贸易的生物固碳方式[J].植物生态学报,2006,30(2):200-209.
[29]DUARTE C M,CEBRIAN J.Plant growth-rate dependence of detrital carbon storage in ecosystem.Limnology and Oceanoqraphy,1996,41:1758-1766.
[30]BOUILLON S,BORGES A V,CASTANEDAMOYA E,et al.Mangorove production and carbon sinks:A revision of global Budget estimates.Global Biogeochemical Cycle,2008,22:GB2013.
[31]CEBRIAN J,DUARTE C M.Plant growth-rate dependence of detrital carbon storage in ecosystem.Science,1995,268(5217):1606-1608.
[32]LIAO C Z,LUO Y Q,FANG C M,et al.Litter pool sizes,decomposition,and nitrogen dynamics inSpartinaalterniflora-invaded and native coastal marshlands of the Yangtze estuary.Oecologia,2008,156:289-600.
[33]董洪芳,于君宝,孙志高,等 .黄河口滨岸潮滩湿地植物—土壤系统有机碳空间分布特征[J].环境科学,2010,31(6):1594-1599.
[34]DING W X,ZHANG Y H,CAI Z C.Impact of permanent inundation on methane emissions from aSpartinaalternifloracoastal salt marsh,Atmospheric Environment,2010,44:3894-3900.
[35]梅雪英,张修峰 .长江口典型湿地植被储碳、固碳功能研究:以崇明东滩芦苇带为例[J].中国生态农业学报,2008,16(2):269-272.
[36]梅雪英,张修峰 .长江口湿地海三棱藨草(Scirpus mariqueter)的储碳、固碳功能研究:以崇明东滩为例[J].农业环境科学学报,2007,26(1):360-363.
