舟山群岛2种濒危植物生境特征与迁地保护研究
2014-05-29王凤英彭红玲胡永红
王凤英,田 旗,彭红玲,胡永红
(中国科学院 上海辰山植物科学研究中心/上海辰山植物园,上海 201602)
海洋岛屿生态系统因具有明显的海域地理隔离而区别于陆地生态系统,被誉为生物地理与进化生态学研究的 “天然实验室”[1]。岛屿物种由于分布范围局限、生境脆弱且种群规模较小,较陆地物种具有更高的灭绝风险[2-3]。17世纪至20世纪地球上灭绝的维管植物共计384种,其中岛屿植物达139种,约占36%;渐危或濒危维管植物中岛屿分布种类约占40%[4]。舟山群岛位于长江口以南,杭州湾外缘的东海海域,地史较为年轻,至今为7000~9000 a[5]。舟山群岛自与大陆脱离后,历经数千年海洋性气候的作用,在植物区系性质上已与大陆形成一定差异,历来为中外学者所关注[6]。舟山群岛自然分布的濒危植物达21种,其中普陀樟Cinnamomum japonicum var.chenii和舟山新木姜子Neolitsea sericea为国务院1999年批准的《国家重点保护野生植物名录:第1批》中列入的国家二级重点保护植物[7-8]。普陀樟系樟科Lauraceae樟属Cinnamomum常绿乔木,间断分布于中国东部沿海岛屿,自上海金山县大金山岛西坡延伸至浙江舟山群岛,中国台湾、朝鲜、日本也有分布,常见于海拔200~400 m的低山沟谷常绿阔叶林内[9-10]。舟山新木姜子为樟科新木姜子属Neolitsea常绿木本孑遗植物,分布于浙江舟山群岛(上海崇明佘山屿曾有分布的记录)海拔150~350 m的丘陵谷地,中国台湾、日本以及朝鲜半岛的沿海岛屿也有分布[11]。由于这2种植物具有很高的观赏价值,被广泛开发用于园林绿化。近些年来人为挖掘现象加重,人为干扰可能是导致这2种植物种群数量急剧减少的最主要原因。鉴于岛屿生境的脆弱性所导致的物种濒危状态日趋严峻,对岛屿植物尤其是岛屿濒危植物的保护迫在眉睫。历史的经验和现状告诉我们,稀有濒危物种需要在人类的帮助下才能得到有效的保护,迁地保护就是其中最主要的措施之一[12]。植物园是迁地保护稀有濒危植物最主要的场所。综合气候特点、生境特点、保护区工作的便利等各方面因素,将辰山植物园内的辰山西北坡作为保存物种的迁地保护区,将普陀樟和舟山新木姜子的野外种群迁地到辰山上长期保护及繁衍。
1 材料与方法
1.1 材料
本研究主要以舟山群岛野生群落中的普陀樟和舟山新木姜子为研究对象,对其自然分布、群落组成、土壤等生境特点进行了调查与分析,同时于2008-2009年将这2种植物迁地保护于上海辰山植物园内。
1.2 研究方法
1.2.1 群落调查 在物种所在群落内设置10 m×10 m的样方,调查了普陀岛、桃花岛、朱家尖、大猫岛、悬鹁鸪岛、东白莲岛及西峰岛等分布区以及迁地保护区的群落组成,用每木调查法统计样方内的乔木和灌木植物。在10 m×10 m的大样方内随机选择5个1 m×1 m的小样方,记录样方内草本层植物。
1.2.2 生境土壤特征 在野外进行2种植物的群落调查时,舟山群岛分布有普陀樟和舟山新木姜子的几个岛屿(普陀岛、桃花岛、朱家尖、悬鹁鸪岛、东白莲岛、西峰岛、大猫岛、梅西岛)及迁地保护区分别进行土壤物理性状调查,在样方内沿样方对角线两端和中间用混合采样法采集0~30 cm土样,混合均匀后带回风干后,送上海市园林科学研究所土壤分析室分析化学性质。土样测定方法:pH值用LY/T 1239-1999《森林土壤pH值的测定》方法测定;电导率用LY/T 1251-1999《森林土壤水溶性盐分分析》方法测定;有机质用LY/T 1237-1999《森林土壤有机质的测定及碳氮比的计算》方法测定;碱解氮用LY/T 1229-1999《森林土壤水解性氮的测定》方法测定;有效磷用LY/T 1233-1999《森林土壤有机磷的测定》测定;速效钾用LY/T 1236-1999《森林土壤速效钾的测定》测定。
1.2.3 物候观测与记录 对新定植的植物定期进行物候观测,主要记录叶变化期 [芽萌动期、新叶出、幼叶期(颜色)、成叶期(颜色)],花变化期[花蕾(颜色)、初花期(颜色)、盛花期、落花期],果变化期[幼果出现(颜色),果实成熟(颜色)、果落期]。
2 结果与分析
2.1 自然生境与迁地保护区生境特征比较
2.1.1 自然环境气候特征 舟山群岛气候属北亚热带南缘海洋性季风气候,年平均气温为15.6~16.6℃,极端最低气温为-7.9℃,年平均降水量为936.3~1330.2 mm,年平均日照时数为1941~2257 h,无霜期为251~303 d,由于受季风不稳定性影响,夏季易受热带风暴(台风)侵袭,冬季多大风,7-8月间常遇干旱[8]。迁地保护区属于亚热带季风气候,受冷暖空气影响,四季分明,气候温和,雨水充沛,日照充足,无霜期长。年平均气温为15.4℃,极端最低日气温-10.5℃,年平均降水量1103.2 mm,年平均日照时数为1997.1 h,年均无霜期为230 d。迁地保护地选在上海辰山植物园的辰山上,辰山山体总高71 m。
2.1.2 群落特征 笔者对舟山群岛普陀樟和舟山新木姜子的几个样地进行了调查与分析。普陀樟所处群落中乔木层以普陀樟,朴树Celtis sinensis,天仙果Ficus beecheyana和红楠Machilus thunbergii为主,它们的重要值分别为73.98,42.48,40.43和30.21,这些物种在群落中占有优势地位;灌木层普陀樟幼苗占优势,其他灌木种类较少;草本层中以灯台莲Arisaema bockii,大吴风草Farfugium japonicum等植物比较丰富[13]。舟山新木姜子所处群落中(表1)乔木层以舟山新木姜子,杉木Cunninghamia lanceolata,野鸦椿Euscaphis japonica,红楠和朴树为主,这些物种的重要值分别为75.10,35.00,31.11,31.01和24.55,在群落中占有优势地位;灌木层舟山新木姜子苗和茶树Camellia sinensis苗略微占优势;草本层以鳞毛蕨属Dryopteris植物及舟山新木姜子小苗占优势。从对辰山迁地保护区的原生境群落调查结果分析(表2),乔木层以刺槐Robinia pseudoacacia,樟树Cinnamomum camphora,构树Broussonetia papyrifera,朴树,冬青Ilex chinensis等为主,灌木层中小蜡Ligustrum sinense占优势,草本层主要以野菊花Dendranthema indicum和蓬藟Rubus hirsutus为主。

表1 舟山新木姜子群落乔木层树种的群落特征Table1 Characteristics of species for tree layer of Neolitsea sericea community
2.1.3 土壤特征 由表3可见:普陀樟原生境砂石少,土层厚度大于30 cm,容重为1.0~1.2 g·cm-3,孔隙度较大。pH 4.70~5.23,典型酸性土壤,电导率为0.04~0.06 mS·cm-1,有机质为 20.67(悬鹁鸪岛)~48.84 g·kg-1(桃花岛),水解性氮质量分数为47.45(朱家尖)~163.79 mg·kg-1(普陀岛),有效磷质量分数为 4.64(西峰岛)~13.37 mg·kg-1(桃花岛),速效钾质量分数为 66.99(东白莲岛)~295.7 mg·kg-1(普陀岛)。舟山新木姜子野生生境砂石较多,土层厚度约30 cm。土壤养分状况良好,舟山新木姜子原生境土壤主要为壤土,其中以黏壤土居多。酸性土壤,pH 4.82~5.38,电导率为0.04~0.10 mS·cm-1,有机质质量分数为 24.18 (朱家尖)~69.31 g·kg-1(桃花岛)。水解性氮质量分数为 45.55(朱家尖)~230.20 mg·kg-1(桃花岛),有效磷质量分数为5.48(大猫岛)~56.09 mg·kg-1(梅西岛),速效钾质量分数为60.80(朱家尖)~371.05 mg·kg-1(桃花岛)。迁地保护地土壤以粉砂质黏壤土和粉砂质壤土为主,个别地方也有砂土和黏土。山体上较厚的枯枝落叶层,成为植株生长所需营养的储存库。大部分土层厚度约60 cm,个别山石多的地方仅20 cm。辰山土壤pH 6.01,电导率为0.05 mS·cm-1,有机质为58.99 g·kg-1,水解性氮质量分数为221.21 mg·kg-1,有效磷质量分数为20.52 mg·kg-1,速效钾质量分数为69.30 mg·kg-1。

表2 迁地保护区群落乔木层树种的群落特征Table2 Characteristics of species for tree layer of Cinnamomum japonicum var.chenii community
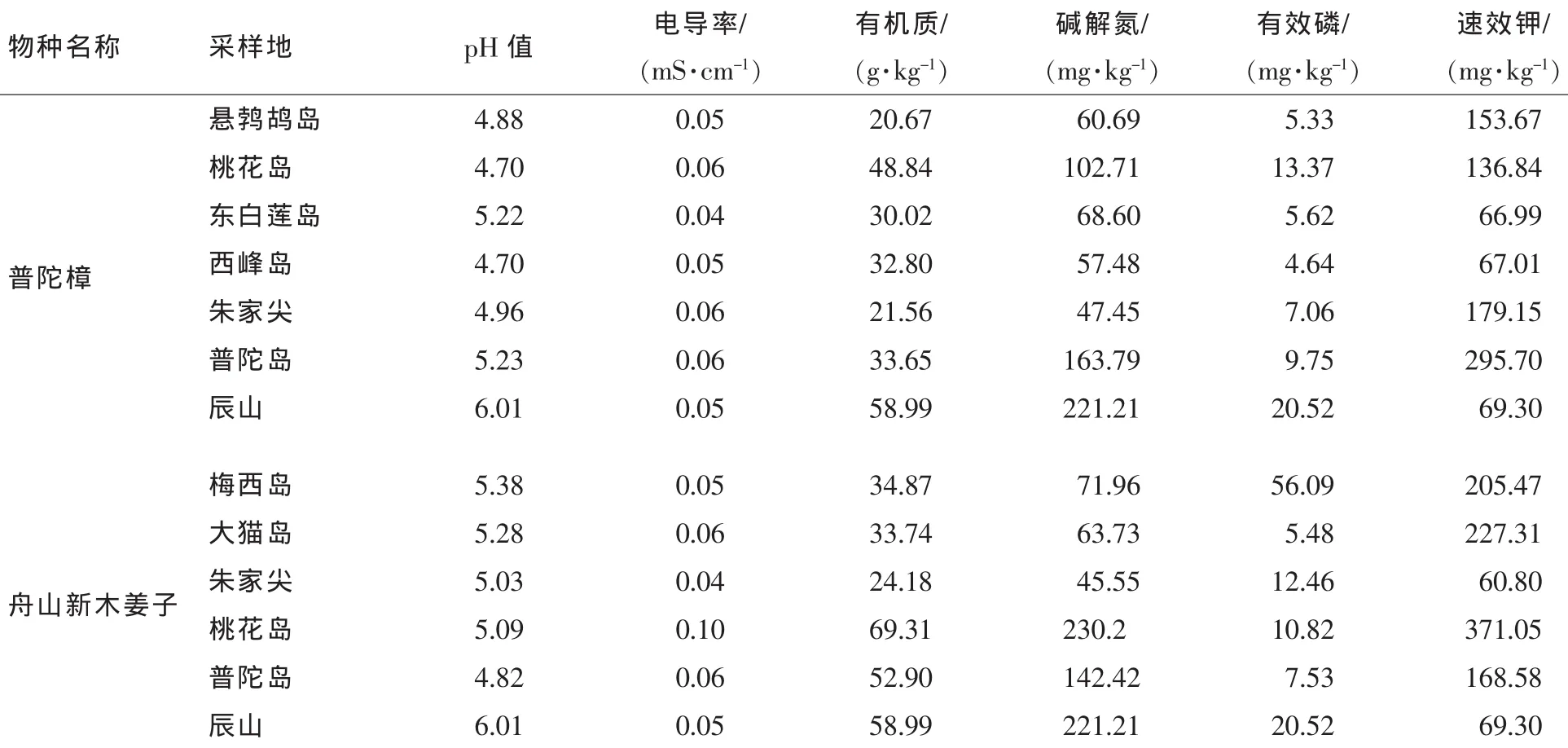
表3 普陀樟和舟山新木姜子原生境和迁地保护区生境土壤特征比较Table3 Soil characteristic indices of Cinnamomum japonicum var.chenii and Neolitsea sericea between the natural and ex-situ habitats
2.2 迁地保护区的建立
2.2.1 迁地保护区改造 为给迁地保护植物足够的生长空间,对原有群落进行疏伐除杂,主要采取间疏式伐木,重点清理刺槐、构树及带刺类灌木,适当修剪大树树冠,为定植植株提供充足的阳光和合适的生态位。随着这些植物的生长再逐渐去枝、疏伐,以满足植株在不同发育阶段对光照等环境条件的要求。
2.2.2 植物迁地保护群落的组建 群落的组成与结构的设计要满足保护物种及其遗传多样性的要求。不仅要保证不同物种不同发育阶段的植物生长所要求的条件,使其能正常繁育,最终达到从种子到种子的自行繁衍;还需保证整个群落的稳定性及不同植物间的相互协调性,对群落中的主要优势种群能保证其持续稳定的发展,且能基本上保持其遗传组成与遗传结构的稳定[14]。由于大的乔木引种困难,而且成活率低,在群落建造初期,主要以来源于野外实生苗和野外采种播种繁殖的小苗为主。因此,初建成的群落仍以原生境物种为优势种。为保证普陀樟和舟山新木姜子完整的遗传多样性,尽可能收集了多个分部区的植株。普陀樟引自舟山群岛的桃花岛、悬鹁鸪岛、西峰岛、东白莲岛和朱家尖等5个分布区,舟山新木姜子引自舟山群岛的大猫岛、东白莲岛、朱家尖以及桃花岛的大佛岩和对峙山。于2008-2009年共种植了普陀樟76株,舟山新木姜子67株,满足了最小可存活种群50株的要求。根据对这2种植物野外群落调查的结果,栽植以下优势伴生种:红楠、朴树、野鸦椿、天仙果和鳞毛蕨科Dryopteridaceae植物等。苗木定植后,初期对每株植株搭三脚架起支撑防风作用,冬季将树干用麻袋片绑好保温,并且用草帘搭防风墙,在定植后1 a之内,对植株进行了精细养护,及时对植株叶片和树干进行喷水,视具体情况进行浇水,在生长速度最快的季节进行施肥,树穴内及周围经常除草松土,伐去妨碍植株生长的乔、灌枝条,遇到病虫害及时采取措施。2010年的统计结果表明普陀樟和舟山新木姜子在迁地保护区的成活率分别为91%和88%。
2.2.3 物候观测记录 对迁地引种的普陀樟和舟山新木姜子连续2 a的物候观测与记录(表4),基本掌握了它们在迁地保护区的物候期。普陀樟4-6月抽新叶,5月开花,10-11月结果。舟山新木姜子4-6月抽新叶,11-12月开花,第2年10-12月结果。

表4 普陀樟和舟山新木姜子在迁地保护区的物候记录Table4 Phenological period of Cinnamomum japonicum var.chenii and Neolitsea sericea in ex-situ conservation
3 结论与讨论
3.1 自然生境与迁地保护区生境特征比较
首先,根据20世纪初期德国慕尼黑大学迈依尔提出的 “气候相似论”,木本植物引种成功的最大可能性在于树种原生地和新栽培地区的气候条件有相似的地方。舟山群岛和上海辰山植物园均隶属于华东东部地区,经纬度十分接近,均属于亚热带气候,年平均温度约为15℃,年平均降水量约1000 mm,年平均日照时数约2000 h,温度、降水、日照等气候参数都很接近,满足了气候相似的原则。其次,普陀樟和舟山新木姜子野外所处的群落各层次物种种类有一定程度的相似性,与迁地保护区群落物种相似性不高,但我们在引种普陀樟和舟山新木姜子的同时,还引种了原生境的伴生种,营造了与野生群落相似度较高的群落环境。此外,在生态条件相似的迁地保护过程中,土壤条件也是必不可少的因素,普陀樟、舟山新木姜子原生境与迁地保护区土壤均呈酸性。普陀樟、舟山新木姜子原生境所有样点有机质质量分数均高于20 g·kg-1(园艺发达国家最低标准),说明普陀樟和舟山新木姜子原生境土壤有机质丰富,而迁地保护区辰山上的土壤有机质质量分数达到58.99 g·kg-1,足以能保证迁地植物的正常生长。土壤速效养分是衡量土壤养分供应能力的现实指标。从样点分布来看,桃花岛样地各速效养分质量分数整体处于较高水平,朱家尖较其他样地来说偏低,迁地保护区除速效钾质量分数稍偏低外,其他2项指标均处于较高水平。以上土壤理化特征显示普陀樟和舟山新木姜子原生境与迁地保护区土壤酸碱性一致,原生境土壤相关肥料指标与迁地保护区也基本相似,足以满足普陀樟、舟山新木姜子的生长。综上所述,普陀樟和舟山新木姜子原生境与迁地保护区生境中气候条件、群落特征、土壤特征均有很大程度的相似性,有利于迁地保护的成功。
3.2 迁地保护区的建立
迁地保护群落垂直结构的形成是一个动态的过程。在群落建造初期,原生境中的物种起着至关重要的作用,随着群落的动态变化,迁地保护植物在群落中的作用和地位会逐渐凸显。采用逐步疏伐的方法是根据不同植物的生态生物学特性为各物种选择合适的生态位,并在植株生长发育过程中及时去枝或疏伐,以满足植株在不同发育阶段对光照等环境条件的要求。经适当疏伐后,基本上保留着原生境群落的垂直结构不变,新栽植的小苗在群落中只是充当了灌木层和草本层的角色。引进乔木植物的种群结构在群落建造初期因无法满足不同年龄级的个体要求,故需要采用分期分批定植的方案,在植物不断生长过程中逐步完善各种群的年龄结构。
迁地保护的最终目的是要保护多样性的物种遗传基因。普陀樟和舟山新木姜子各引种了5个种群,多个种群的引入不仅能够更完整地保存物种的遗传多样性,还有利于基因交流,突破了传统的引种来源单一问题。
3.3 迁地保护有效性评价
迁地保护的有效性评价需综合考虑数量、材料来源、适应性等多方面因素。本研究迁地保护了普陀樟和舟山新木姜子分别为76和67株,满足了迁地保护最小种群大于50株的理论要求[16]。分别引入5个不同来源地的种群,满足了保存物种遗传多样性的要求。此外,经后期观察与物候记录,普陀樟和舟山新木姜子已经在迁地保护正常开花与结果。从目前的理论研究及植物实际的长势来观察,迁地保护获得了一定的成效。
[1]魏娜,王中生,冷欣,等.海洋岛屿生物多样性保育研究进展[J].生态学杂志,2008,27(3):460-468.WEI Na,WANG Zhongsheng,LENG Xin,et al.Conservation of oceanic island biodiversity∶a review [J].Chin J Ecol,2008,27(3)∶460 - 468.
[2]FRANKHAM R.Do island populations have less genetic variation than mainland populations [J]Heredity,1997,78∶311-327.
[3]RAVEN P H.Plant conservation in a changing genetics of endangered island plants [M]//AVISE J C,HAMRICK J.Conservation Genetics:Case Histories from Nature.New York∶Chapman and Hall,1998:305 - 334.
[4]REID W V,MILLER K R.Keeping Options Alive:The Scientific Basis for Conserving Biodiversity [R].Washington D C∶World Resources Institute,1989.
[5]LI Yiming,NIEMELA J,LI Dianmo.Nested distribution of amphibians in the Zhoushan archipelago,China∶can selective extinction cause nested subsets of species[J].Oecologia,1998,113∶557 - 564.
[6]万利琴,丁炳扬,郭水良.舟山群岛主要岛屿间种子植物区系的差异及其影响因素[J].浙江大学学报:农业与生命科学版,2008,34(6)∶677-683.WAN Liqin,DING Bingyang,GUO Shuiliang.Difference of spermatophyte flora among main islands of Zhoushan Archipelago and its influencing factors [J].J Zhejiang Univ Agric&Life Sci,2008,34(6)∶677 - 683.
[7]傅立国.中国植物红皮书——稀有濒危植物:第1册[M].北京:科学出版社,1991.
[8]俞慈英,陈叶平,袁燕飞,等.舟山群岛普陀樟等3种特有树种种质资源清查[J].浙江林学院学报,2007,24(4):413 - 418.YU Ciying,CHEN Yeping,YUAN Yanfei,et al.Resource s of Cinnamomum japonicum var.chenii,Neolitsea sericea and Tilia miqueliana in Zhoushan archipelago [J].J Zhejiang For Coll,2007,24(4)∶413 - 418.
[9]王景祥.浙江植物志:第2卷[M].杭州:浙江科学技术出版社,1992:352.
[10]张若蕙.浙江珍稀濒危植物[M].杭州:浙江科学技术出版社,1994:156.
[11]王中生,安树青,冷欣,等.岛屿植物舟山新木姜子居群遗传多样性的RAPD分析[J].生态学报,2004,24(3):414 - 422.WANG Zhongsheng,AN Shuqing,LENG Xin,et al.Population genetic diversity of the insular plant Neolitsea sericea based on random amplified polymorphic DNA (RAPD)[J].Acta Ecol Sin,2004,24(3)∶414 - 422.
[12]康明,叶其刚,黄宏文.植物迁地保护中的遗传风险[J].遗传,2005,27(1):160-166.KANG Ming,YE Qigang,HUANG Hongwen.Genetic risks in plant ex-situ conservation [J].Hereditas,2005,27(1)∶160-166.
[13]田旗,黄姝博,胡永红.中国特有植物普陀樟群落特征及物种多样性[J].植物分类与资源学报,2010,32(增 17):81- 88.TIAN Qi,HUANG Shubo,HU Yonghong.Characteristics and species diversity of Cinnamomum japonicum var.chenii(Lauraceae) community in China [J].Plant Diversity Resour,2010,32(supp 17)∶81 - 88.
[14]叶其刚,王畅,王诗云.三峡库区稀有濒危植物异地保护群落设计的初步研究[J].武汉植物学研究,2000,18(1):33 - 41.YE Qigang,WANG Chang,WANG Shiyun.A preliminary study on the design of a community for ex-situ conservation of the rare and endangered plants in the Three Gorge Reservoir Area [J].J Wuhan Bot Res,2000,18(1)∶33 - 41.
[15]许再富.稀有濒危植物迁地保护的原理与方法[M].昆明:云南科技出版社,1998.
