水、氮控制对短花针茅草原气体交换的影响
2014-05-08李寅龙红梅白文明韩国栋王海明
李寅龙,红梅*,白文明,韩国栋,王海明
1. 内蒙古农业大学生态环境学院,呼和浩特 010018;2. 中国科学院植物研究所,北京 100093;3. 内蒙古农牧科学院,呼和浩特 010031
由于全球气候的变化,引起降雨量的不均匀分布,部分地区高的降雨量而部分地区降雨严重匮乏,而且年内的降雨量与年际间的降雨量发生了大的改变,因而影响了全球陆地生态系统碳的循环和收支平衡。降低的降雨量将影响土壤呼吸和生态系统光合,导致了生态系统净碳吸收的减少。降雨量的减少能降低陆地生态系统总的初级生产力和生态系统净碳交换(Ciais et al, 2005;Granier et al,2007)。Ogaya 和 Peñuelas (2007)在一个 holmoak 森林的研究发现降雨量的降低不仅影响了植物生长和生理生态过程,而且也影响了生态系统碳的循环,同时导致了大量植物物种的大量死亡。在干旱和半干旱地区,相比温度和CO2浓度的增加, 改变的降雨量对于生态系统碳交换有更大的影响,从而导致了生态系统碳交换的减少(Potts et al, 2007;Weltzin et al, 2003)。同样的研究结果在Niu等(2009)的模型学习中和Breshears等(2005)的野外实验研究中被证实。Meyers(2001)研究了美国Oklahoma西南部的草原, 发现干旱引起了土壤呼吸的增加,从而导致了生态系统碳交换的降低。国内,Xia等(2009)对内蒙古典型草原的两年野外研究发现(降雨量差异大的)在干旱的年份碳的释放增加。
由于人类社会工业的发展,工业化速度的加快以及化肥用量的急速增长,近年来,全世界范围内通过大气沉降返回陆地生态系统的氮元素正在急剧增加(Gruber和Galloway, 2008)。目前,国内外对氮素与碳循环方面的研究主要集中于对植物生长与净初级生产力(NPP)方面,而且大多数的研究结果表明氮素添加提高了植物生长与陆地生态系统净初级生产力(NPP)(Xia和Wan, 2008)。例如,近来两个利用 Meta-analysis的综述性研究都表明氮添加显著增加了陆地生态系统净初级生产力(LeBauer和Treseder, 2008)。生态系统碳循环及平衡是由碳吸收与释放这两个主要过程共同决定的,即通过生态系统的光合作用(Gross ecosystem productivity, GEP)吸收大气CO2,然后再通过生态系统的呼吸作用(Ecosystem respiration, ER)返回到大气中。目前为止,尽管全球已经开展了大量植物生长与NPP对氮素添加响应的研究,但有关氮素添加与NEE响应关系的研究却仍然很少。过去的几年中,国内外学者对氮素添加如何影响生态系统两个过程(GEP和ER)进行了一系列的研究,通过这些试验研究期待得出生态系统净 CO2交换(NEE)对氮素添加的响应规律。这些控制试验涉及一系列生态系统类型,包括湿润莎草苔原(Shaver et al, 1998),北方沼泽(Saarnio et al, 2003),北美一年生草地(Harpole et al, 2007)和多年生典型草地(Niu et al, 2010)等。同时,一些研究也通过模型预测(Pepper et al, 2005)和样地研究(Havström et al, 1993)等方法对 NEE如何响应氮素添加进行了研究,但至今仍没有得到一个统一的结论,像Shaver等(1998)的研究结果表明氮素添加对生态系统NEE具有显著正效应,而其他的结果则发现氮素添加对生态系统NEE没有影响(Saarnio et al,2003)。Xia等(2009)对国内温带草原的研究发现氮素的添加显著增加了GEP而没有影响ER,因而导致了整个温带生态系统表现为净碳的吸收(NEE)。另外,由于气候变暖的一个重要机制是通过刺激土壤氮的矿化作用(Rustad et al, 2001)影响陆地生态系统碳循环,因此可能氮素添加与气候变暖下降雨的变化共同影响生态系统 CO2交换,从而削弱大气CO2浓度背景下发生的“N限制”过程(Luo et al,2004)带来的严重影响。
1 试验区自然条件
1.1 研究区概况
试验地位于内蒙古中部、大青山北麓内蒙古乌兰察布市四子王旗王府一队。四子王旗境内地形复杂,由南部的低山丘陵和北部的层状高平原两大地貌单元组成。北部有沙丘、盆地,为荒漠草原,地势较平坦,是较大的天然牧场。整个地形东南高西北低,略呈长方形。四子王旗草原属乌兰察布草原的主要组成部分,草原面积214.3×104hm2,占全旗总面积的89%,天然植被为主。研究点位于四子王旗的内蒙古畜科院研究基地,北纬41°47′17″,东经111°53′46″;海拔 1456 m。地势平坦,地表无沙化和水土流失现象。
1.2 气候条件
该地区属中温带干旱、半干旱大陆性季风气候,具有光资源丰富,日照充足,太阳辐射强的特点。全年降水量较少,春季干旱多风,夏季炎热。多年平均降水量为248 mm,湿润度为0.15~0.30,年均蒸发量2947 mm,为降水量的6~16倍,同时降水多集中在 6—9月份,占全年降水量的60%~70%,年平均气温2~5 ℃,7月均温19~22 ℃,1月均温-15~18 ℃,且年差较大,日差也较大。年均风速4~5 m⋅s-1,≥8级以上大风日可达50~70 d,风季集中在11月至翌年5—6月份,尤以3—5月最强,常形成尘暴、扬沙天气。干旱、低温、风大是该区的气候特点,也是形成风蚀的动力。春季大风与干旱同期出现,是土地沙化的重要气候因素。
1.3 土壤与植被
土壤类型为淡栗钙土。土壤较瘠薄,腐殖质层厚约20~30 cm,有机质质量分数10~18 g⋅kg-1,钙积层分布在20~30 cm,土壤微生物有好气性细菌、放线菌和霉菌等。实验样地所在地区土壤质地为砂壤土,原生植被类型为短花针茅(Stipa breviflora)+冷蒿(Artemisia frigida)+无芒隐子草(Cleistogenes songorica)荒漠草原,植被草层低矮,一般为8 cm,且植被较稀疏,盖度为 17%~20%,种类组成较贫乏,植物群落由 20多种植物组成。建群种为短花针茅(Stipa breviflora),优势种为冷蒿(Artemisiafrigida)、无芒隐子草(Cleistogenes songorica)。主要伴生种有阿氏旋花(Convolvulus ammannii)、阿尔泰狗娃花(Heteropappus altaicus)、栉叶蒿(Neopallasia pectinata)、羊草(Leymus chinesis)、细叶葱(Alliumtenuissimum)、木地肤(Kochia prostrate)、小叶锦鸡儿(Caragana microphylla)等(包秀荣, 2009)。
2 研究方法
2.1 试验设计
水分和氮素二因素交互试验2010年8月,采用了裂区设计,6种处理(对照—CK、增雨—W、减雨—R、施氮—N、增雨施氮—WN、减雨施氮—RN),每种处理6次重复,共36个处理, 随机区组排列。共有24个面积15 m × 9 m的小区,小区间隔2 m,12个减雨处理(R和RN处理)是在其他处理的基础上增加减雨架,面积为3 m × 3 m。增雨减雨量为多年各月份平均降雨量的 30%(58 mm)。增雨于每年的雨季(5—8月)的月初进行,使用喷洒装置进行模拟增雨,水滴均匀落在样地内,并保证不形成地表径流;减雨全年都在进行,使用自制的减雨架进行模拟减雨,减雨架遮挡面积为减雨小区面积的30%。氮素添加处理所施氮肥是NH4NO3(简称硝铵,NH4NO3,含氮33%~35%),施肥量为纯氮10 g⋅m-2。在每年6月末或7月初雨季来临时进行喷施,以防止高温干旱的条件导致的NH4NO3挥发。施肥为一次施完。
2.2 取样及样品测定方法
生态系统气体交换(CO2和水分通量)的测定是采用一个自制大叶室(0.5 m×0.5 m×0.5 m),内部固定两个小型风扇,目的是用来搅拌气体使其在最短的时间里尽量的混合均匀,同时也可以使大叶室内部的温度均匀一致,以利于交换气体的测定,在测定时把大叶室和 LI-6400(IRGA;LI-6400,LI-Cor,Lincoln,NE,USA)进行连接,把大叶室密闭的放置在铝合金框上,开启LI-6400就可以进行测定了。根据四子王旗当地的实际情况,每一个样点每次测定时间为120 s,每10 s一起自动记录一次 CO2浓度和水分通量数值,根据时间序列和CO2浓度和水分通量值可以计算出净生态系统交换量(NEE)和生态系统蒸散量(ET)。接下来拿起大叶室使其内部气体扩散,温度和大气温度达一致后,重新放置在铝合金框上并用黑色布遮盖大叶室使光源不能进入,按上面同样的方法测定120 s,记录的CO2浓度数据和时间序列计算所得的就是生态系统呼吸值(ER)。试验数据为2012年采集,总的生态系统生产力(GEP)的计算就是NEE和ER之差。我们这里规定负的NEE为净C的吸收固定,正值为C的释放。
2.3 数据分析
采用 Excel 2003进行数据统计和图表构建,SAS 9.0统计软件进行数据的统计分析。
3 结果与分析
3.1 控制性降雨对生态系统气体交换的影响
水分是荒漠草原生态系统生物生长的重要限制因素,降雨量的多少决定生态系统系统的生产力变化。从图1、图2、图3可知,从整体生长季来看CK、R、W处理的NEE、ER、GEP都呈不对称单峰曲线,NEE和GEP在6月到8月都呈上升趋势,并在8月达到最大值。ER在5月到8月都呈上升趋势,并在8月达到最高值。由表1可知NEE、ER、GEP在R处理中明显低于CK处理和W处理,就NEE而言,CK和R处理之间差异显著,CK和W处理之间差异不显著;在ER中CK、R和W处理之间差异显著;在GEP中R处理与CK、W差异显著,CK和W处理之间差异不显著。
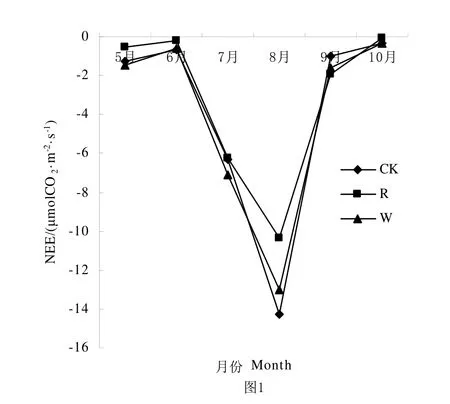
图1 不同降雨量条件下NEE生育期动态变化Fig.1 Seasonal variation of NEE under different rainfall
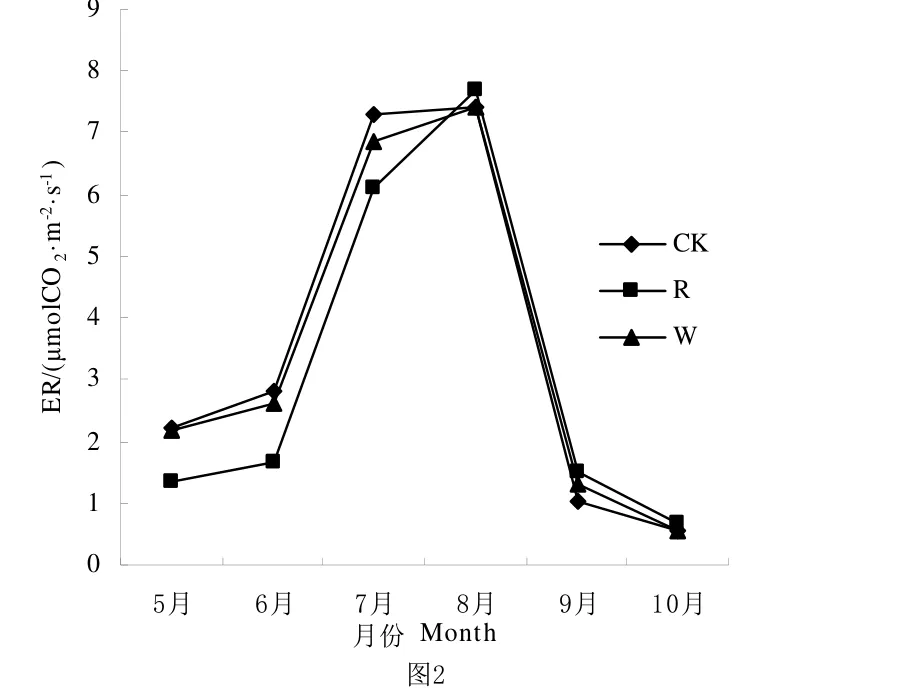
图2 不同降雨量条件下ER生育期动态变化Fig.2 Seasonal variation of ER under different rainfall
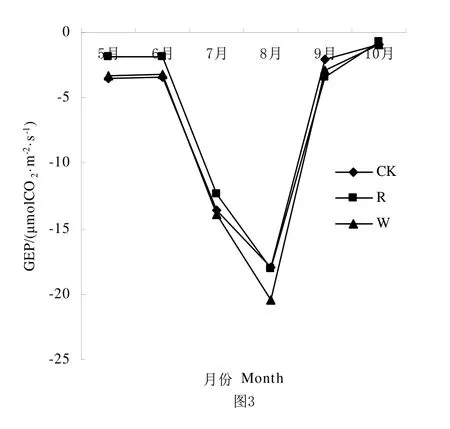
图3 不同降雨量条件下NEE、ER和GEP生育期动态变化Fig.3 Seasonal variation of GEP under different rainfall

表1 短花针茅草原不同水分处理NEE、ER和GEP的变化Tabel 1 The change of NEE、ER and GEP under different rainfall treatments in the Stipa breviflora grassland(µmol·m-2·s-1,以 CO2 计)
3.2 氮素添加对生态系统气体交换的影响
氮素的添加显著的影响了全球碳的收支平衡,同时氮素的增加也能调节气候变化对生态系统碳循环的影响[17]。图4、图5、图6显示整体生长季CK和N处理的NEE、ER和GEP在5月到8月都呈上升趋势,并在8月达到最大值,此后随着温度降低开始下降。由表2可知NEE、ER和GEP之间CK和N处理之间差异都不显著,但N处理明显高于CK处理。

图4 不同水氮条件下NEE生育期动态变化Fig.4 Seasonal variation of NEE under different rainfall and nitrogen conditions
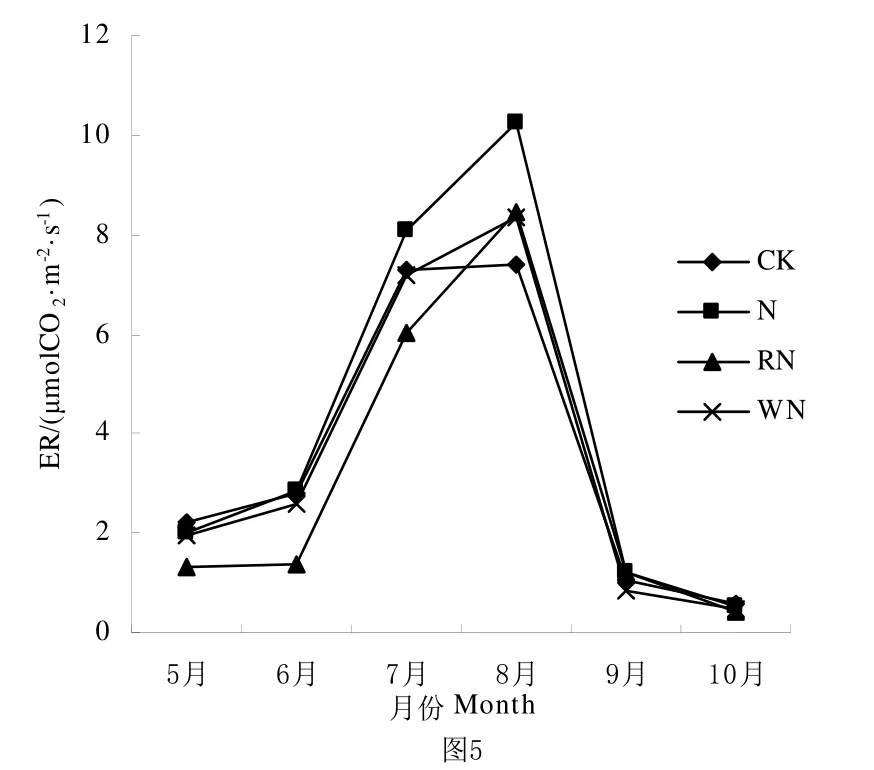
图5 不同水氮条件下ER生育期动态变化Fig.5 Seasonal variation of ER under different rainfall and nitrogen conditions

图6 不同水氮条件下GEP生育期动态变化Fig.6 Seasonal variation of GEP under different rainfall and nitrogen conditions
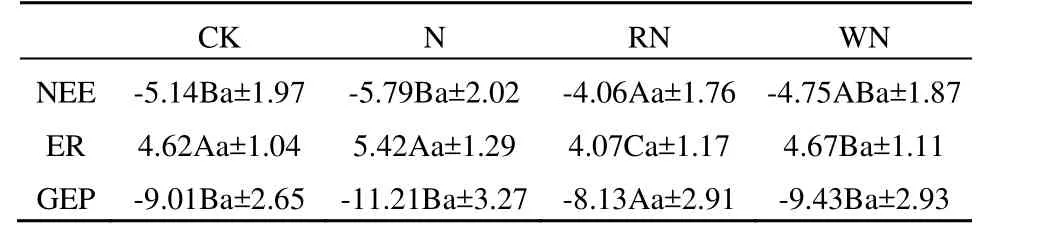
表2 短花针茅草原不同水分和氮素添加处理NEE、ER和GEP的变化Tabel 2 The change of NEE、ER and GEP under different rainfall treatments and N addition in the Stipa breviflora grassland(µmol·m-2·s-1,以 CO2 计)
3.3 氮水交互作用对生态系统气体交换的影响
单独N和W处理分别对NEE有一定的增大(表1、表2)。而RN和WN处理NEE都有降低。但是WN处理的GEP、ER都有所增加;而RN处理的GEP、ER却都下降了。方差分析结果表明,ER各处理之间年度差异达到显著性水平(P<0.05),大小顺序为WN>CK>RN;GEP中RN处理与其他处理之间差异显著。WN和CK处理之间没有显著性差异;NEE中RN和CK之间差异达到显著性水平。
4 讨论
4.1 不同降雨量对生态系统气体交换的影响
对于陆地生态系统在全球碳循环中到底扮演着怎样的角色这个问题,长期以来一直处于激烈争论之中。早期的研究认为温带生态系统在全球碳循环中主要以碳汇存在于陆地生态系统中(Myneni et al, 2001),而近期的一些研究认为这些生态系统在全球碳循环中的作用并不确定(Aires et al, 2008)。事实上,这些争论之所以出现是因为近期的研究把气候因子的变异考虑在内。大气中不同纬度CO2浓度季节的变化在全球尺度上出示为一个清晰的NEE(Fung et al, 1987)变化图。在北半球高纬度地区,由于夏季温度高,光合作用高于呼吸那导致了大气中CO2浓度的降低,即NEE表现为正效应。相反,在冬天,植物枯黄和死亡,呼吸作用显著的高于光合导致了大气中 CO2浓度的增高,即 NEE表现为负效应(Keeling et al, 1996)。这些研究和我们对内蒙古荒漠草原一年的研究结果一致,即高纬度的荒漠草原样地表现为净碳吸收,Niu等(2008)认为水分的利用率对于调整温带典型草原年内生态系统谈交换扮演着至关重要的角色。我们研究中,生态系统碳的变化与季节的降雨量呈现类似变化的规律,高的生态系统碳的交换出现在降雨量最高的季节,而生态系统碳交换的最低值出现在植物返青期和植物生长的晚期,即降雨量最低的季节。这个研究结果与内蒙古典型草原的研究有相似的结果(Xia et al, 2009;Rustad et al, 2001),同样其他的温带草原系统也得到同样的结果(Aires et al, 2008)。含水量的高低是制约生态系统碳交换的重要因素。Wu等(2010)认为增加的降雨量对生态系统碳循环的影响明显高于减少降雨量对生态系统的影响,这与我们的结果一样。高的降雨量导致了 NEE的增加,同时也增加了植物生物量和生产力。Niu等(2008)的研究发现在降雨量增加的情况下,生态系统总的生产力(GEP)增加量显著高于生态系统呼吸(ER),从而导致了生态系统的净碳吸收增加。同样,降雨量的增加提高了草地生态系统碳的吸收已经在许多的研究中呈现(Rustad et al, 2001;Luo et al,2004)。
4.2 氮素添加对生态系统气体交换的影响
氮素的添加显著的影响了全球碳的收支平衡,同时氮素的增加也能调节气候变化对生态系统碳循环的影响(Niu et al, 2010)。例如,一些研究认为在气候变化的情况下氮素的利用率和氮素在生态系统的运动显著影响了生态系统长期碳的收支(Luo et al, 2004)。在我们的研究中,在植物生长的中期(7、8和 9月)NEE的值显著高于植物生长的早期(5和6月)和晚期(10月)。研究显示在整个植物生长的季节氮的添加对增加了 NEE的值不显著。一个Lolium perenne群落的研究(Zhou et al,2007)和两个半干旱草原的研究(Xia et al, 2009)跟我们的研究结果是一致的。在植物生长季节,氮素的添加增加了生态系统的净碳吸收。氮素是植物生长的重要限制性元素(Vitousek和Howarth, 1991),尤其是在中国北方的荒漠草原。氮素的添加对植物地上部分生产力有直接的影响。氮的添加显著增加植物的地上部分生产力,从而显著提高了 GEP的值。Campbell等(1993)的研究已经证实氮素的利用显著增加了植物根的呼吸从而加速了植物的生长,导致了高的地上部分生产力。在我们的实验研究中,氮素的添加增加了植物根和土壤微生物的呼吸那可能导致了ER值的升高。我们的研究结果支持了氮素的添加增加了生态系统的净碳吸收。
5 结论
在不同降雨量处理下,随着降雨量的变化NEE、GEP、ER都呈先升高再降低的趋势;NEE和GEP下R处理与CK和W处理之间达到显著性水平;ER下各处理之间达到显著性水平(P<0.05),大小顺序为CK>W>R。在施氮处理下,随着降雨量的变化NEE、GEP、ER都呈先升高再降低的趋势;CK与N处理之间差异不显著,NEE、ER和GEP都有所增加。氮水交互作用下,随着降雨量的变化NEE、GEP、ER都呈先升高再降低的趋势;ER各处理之间差异达到显著性水平(P<0.05),大小顺序为WN>CK>RN;GEP中RN处理与其他处理之间差异显著。WN和CK处理之间没有显著性差异;NEE中RN和CK之间差异达到显著性水平。
AIRES L, PIO C, PEREIRA J. 2008. Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years[J]. Global Change Biology, 14: 539-555.
BRESHEARS D D, COBB N S, RICH P M, et al. 2005. Regional vegetation die-off in response to global-change-type drought [J].Proceedings of the National Academy of Sciences of the United States of America, 102(42): 15144-15148.
CAMPBELL B D, STAFFORD SMITH D M, GCTE P. 2000. A synthesis of recent global change research on pasture and rangeland production:reduced uncertainties and their management implications[J].Agriculture, ecosystems & environment, 82: 39-55.
CIAIS P, REICHSTEIN M, VIOVY N, et al. 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003[J].Nature, 437: 529-533.
FUNG I Y, TUCKER C J, PRENTICE K C. 1987. Application of advanced very high resolution radiometer vegetation index to study atmosphere-biosphere exchange of CO2[J]. Journal of Geophysical Research, 92: 2999-3015.
GRANIER A, REICHSTEIN M, BREDA N, et al. 2007. Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year: 2003[J]. Agricultural and Forest Meteorology, 143: 123-145.
GRUBER N, GALLOWAY J N. 2008. An Earth-system perspective of the global nitrogen cycle [J]. Nature, 451: 293-296.
HARPOLE W S, POTTS D L, SUDING K N. 2007. Ecosystem responses to water and nitrogen amendment in a California grassland [J]. Global Change Biology, 13: 2341-2348.
HAVSTRöM M, CALLAGHAN T V, JONASSON S. 1993. Differential growth responses of Cassiope tetragona, an arctic dwarf-shrub, to environmental perturbations among three contrasting high- and subarctic sites [J]. Oikos, 66: 389-402.
KEELING C D, CHIN J F S, WHORF T P. 1996. Increased activity of northern vegetation inferred from atmospheric CO2measurements[J].Letters To Nature, 382: 146-149.
LEBAUER D, TRESEDER K K. 2008. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed [J].Ecology, 89: 371-379.
LUO Y, SU B O, CURRIE W S, et al. 2004. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide [J].Bioscience, 54: 731-739.
MEYERS T P. 2001. A comparison of summertime water and CO2fluxes over rangeland for well watered and drought conditions [J].Agricultural and Forest Meteorology, 106: 205-214.
MYNENI R B, DONG J, TUCKER C J, et al. 2001. A large carbon sink in the woody biomass of Northern forests[J]. Proceedings of the National Academy of Sciences of the United States of America, 98:14784-14789.
NIU S, WU M, HAN Y, et al. 2008. Water mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe [J]. New Phytologist, 177: 209-219.
NIU S, WU M, HAN Y, et al. 2010. Nitrogen effects on net ecosystem carbon exchange in a temperate steppe [J]. Global Change Biology, 16:144-155.
NIU S, YANG H, ZHANG Z, et al. 2009. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe[J]. Ecosystems, 12: 915-926.
OGAYA R, PEñUELAS J. 2007. Tree growth, mortality, and above-ground biomass accumulation in a holm oak forest under a five-year experimental field drought [J]. Plant Ecology, 189: 291-299.
PEPPER D A, DEL GROSSO S J, MCMURTRIE R E, et al. 2005.Simulated carbon sink response of short grass steppe, tall grass prairie and forest ecosystems to rising [CO2], temperature and nitrogen input[J]. Global Biogeochemical Cycles, 19: GB1004.
POTTS D L, HUXMAN T E, ENQUIST B J, et al. 2006. Resilience and resistance of ecosystem functional response to a precipitation pulse in a semi-arid grassland [J]. Journal of Ecology, 94: 23-30.
RUSTAD L, CAMPBELL J, MARION G, et al. 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming [J].Oecologia, 126: 543-562.
SAARNIO S, JäRVIö S, SAARINEN T, et al. 2003. Minor changes in vegetation and carbon gas balance in a boreal mire under a raised CO2 or NH4NO3 supply [J]. Ecosystems, 6: 46-60.
SHAVER G R, JOHNSON L C, CADES D H, et al. 1998. Biomass and CO2flux in wet sedge tundra’s: responses to nutrients, temperature, and light [J]. Ecological Monographs, 68: 75-97.
VITOUSEK P M, HOWARTH R W. 1991. Nitrogen limitation on land and in the sea: how can it occur?[J] Biogeochemistry, 13: 87-115.
WELTZIN J F, LOIK M E, SCHWINNING S, et al. 2003. Assessing the response of terrestrial ecosystems to potential changes in precipitation[J]. Bioscience, 53: 941-952.
WU Z, DIJKSTRA P, KOCH G W, et al. 2011. Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation[J]. Global Change Biology, 17:927-942.
XIA J, NIU S, WAN S. 2009. Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe[J]. Global Change Biology, 15:1544-1556.
XIA J, WAN S. 2008. Global response patterns of terrestrial plant species to nitrogen addition [J]. New Phytologist, 179: 428-439.
ZHOU X, WAN S, LUO Y. 2007. Source components and interannual variability of soil CO2efflux under experimental warming and clipping in a grassland ecosystem[J]. Global Change Biology, 13: 761-775.
包秀荣. 2009. 控制性增温和施氮肥对荒漠草原土壤的影响[D]. 内蒙古农业大学.
