黔南惠水下石炭统上司组礁滩灰岩微相
2014-04-28要乐王向东李越郄文昆林巍
要乐,王向东,李越,郄文昆,林巍
1) 中国科学院南京地质古生物研究所,资源地层学与古地理学院重点实验室,南京,210008 2) 中国科学院大学,北京,100049
内容提要: 黔南惠水雅水剖面下石炭统上司组底部含有厚度约7m的生物沉积和建造灰岩,为上下生屑滩夹含5m厚的珊瑚格架礁,微相类型丰富,可划分出4种类型:① 泥粒状灰岩;② 粒泥状灰岩;③ 珊瑚障积岩;④ 层孔虫盖覆岩。在生屑滩沉积生物碎屑泥粒状灰岩中,珊瑚、腕足类、钙藻、介形类、海百合茎等破碎程度高,分选性低,为较强水动力条件远距离或频繁搬运后快速沉积的产物,此时不适宜大型珊瑚原地生长保存,仅见少量而单调的单体四射珊瑚和复体四射珊瑚。珊瑚格架礁以生物碎屑粒泥状灰岩、珊瑚障积岩为主,发育少量层孔虫盖覆岩和藻类粘结作用。腕足类、钙藻、有孔虫、海百合茎、介形类、腹足类较为丰富,苔藓虫以及层孔虫的丰度未达到支撑程度,昭示珊瑚格架礁生长期的水动力条件减弱,为单体四射珊瑚、笛管珊瑚(Syringopora)和丛管珊瑚(Siphonodendron)为主体的珊瑚群落形成原地礁灰岩创造了条件。
泥盆纪弗拉期(Frasnian)—法门期(Famennian)的生物灭绝事件对浅海底栖群落影响力度大且持续时间长(杜远生等,1990;王向东和沈建伟, 2004)。早石炭世全球碳酸盐岩以灰泥沉积最为常见,苔藓虫和海百合碎片堆积加之菌藻类粘结构建的Waulsortian型泥丘或藻礁居多,缺乏后生动物格架礁(Lees and Miller, 1985; West, 1988; Webb, 2002; 王向东和沈建伟, 2004)。与这段特殊时期的全球普遍实例相对应,华南陆表海区后生动物为造礁主体的实例也十分罕见(巩恩普等, 1998, 2010; Gong Enpu et al., 2012)。目前仅在非常局限的地点报道存在维宪期的小型珊瑚礁(陈晓红等, 2013)和苔藓虫—珊瑚点礁(方少仙和侯方浩, 1986)。精确厘定华南赋存早石炭世生物礁地层的时限和礁灰岩特征,并将这些实例纳入全球同期生物礁宏演化的框架,则可为解析这一特定地史期海洋生物圈的群落景观提供证据。本文通过对下石炭统上司组礁滩灰岩微相特征描述,分析礁、滩形成过程中的古生态细节,从而认识华南生物礁滩的沉积和建造方式。
1 礁滩灰岩概况

图1 (a)和(b) 黔南惠水雅水剖面位置,(b)是(a)中实线框放大;(c) 为(b)中虚线框放大的地质图(改自Wu Xianghe, 1987);(d) 华南早石碳世大塘期古地理图(改自冯增昭等, 1998)Fig.1 (a) and (b)are the location of the Yashui section from Huishui, South Guizhou, (b) is the amplification of the solid frame of (a); (c) the geological map of the amplification of the dashed frame of (b) (revised from Wu Xianghe, 1987); (d) the palaeogeographic map of the Datang Stage, early Carboniferous in South China (revised after Feng Zengzhao et al., 1998)
晚古生代联合古大陆的拼接和多冰期气候事件极大地影响了早石炭世的古地理、古气候和古海洋的频繁变化(Fielding et al., 2008; Wicander and Monroe, 2009)。华南早石炭世裂谷强烈活化(郄文昆和王向东, 2012),导致沉积相分异甚强(冯增昭等, 1998)。其中,位于特提斯构造域东端(秦建华等, 1996)的滇黔桂盆地就是裂谷作用的产物,其形成于早泥盆世晚期(杜远生等, 2009, 2013, 彭阳等,2007,2013),石炭纪—二叠纪继承发展(梅冥相和李仲远, 2004; 彭阳等,2009),至晚三叠世消亡(吕洪波等,2003;梅冥相等, 2003)。滇黔桂盆地周缘的滇黔桂湘时见分布广泛的数个碳酸盐岩台地,台地间可识别出斜坡相带(冯增昭等, 1998; 刘超等,2014)。滇黔桂湘碳酸盐岩台地的下石炭统的碳酸盐岩序列厚度偏大,而滇黔桂盆地则频见较深水环境下的暗色细粒混合沉积物(焦大庆等, 2003)。本文论及的礁滩灰岩出露于黔南惠水县雅水镇北部(图1 a, b),层位是下石炭统维宪阶上司组底部(图1 c),该剖面处于滇黔桂盆地北部且远离大陆物源区的滇黔桂湘碳酸盐岩台地沉积区(图 1 d)。
雅水剖面下部为出露连续的下石炭统维宪阶旧司组,以中厚层灰色泥粒状灰岩和粒泥状灰岩与薄层灰黄色泥岩或钙质泥岩互层为特征;上部为下石炭统维宪阶上司组,其底部赋存厚约7m的礁滩灰岩(图2)。上司组最下部为滩相的灰色厚层泥粒状灰岩,含少量单体四射珊瑚和复体四射珊瑚(图2a);其上为厚层礁相灰岩,出露宽度约10m,厚度约5m,主要造礁生物为单体四射珊瑚、笛管珊瑚(Syringopora)和丛管珊瑚(Siphonodendron),珊瑚含量约40%~70%,且达到原地密集生长程度。根据珊瑚类群由下而上的纵向分布特征,可依次划分为灰色厚层单体四射珊瑚建造型灰岩(A);灰色厚层单体四射珊瑚和横板珊瑚建造型灰岩(B);灰色厚层单体四射珊瑚建造型灰岩(C);灰色厚层复体四射珊瑚建造型灰岩(D)(图2b)。礁相灰岩之上为滩相的灰色中薄层泥粒状灰岩(图2)。由下而上对14个点位进行样品采集(样品编号和采集层位如图2所示),并制作定向岩石薄片153片。

图2 黔南惠水雅水剖面旧司组顶部和上司组底部岩性序列和采样编号,右侧照片(a)和(b)对应左侧柱状图中层位Fig. 2 Lithological sequences and sampling no.s from the top of the Jiusi Formation to the base of the Shangsi Formation at the Yashui Section from Huishui, South Guizhou.Horizons of the right side photoes (a) and (b) shown in the left side column
2 礁滩灰岩微相
张园园等(2009)曾对礁灰岩岩石学分类进行总结,并提出生物礁灰岩普遍具有强烈的非均一性。本文的碳酸盐岩微相鉴定以Dunham(1962)的分类方案为基础。根据颗粒与填隙物(基质和胶结物)的组成比例划分灰岩类型,其中原始沉积组分未被粘结的包括:泥状灰岩(mudstone)、粒泥状灰岩(wackestone)、泥粒状灰岩(packstone)和颗粒状灰岩(grainstone);而原始沉积组分被原地生长的各类生物聚集则形成绑结岩(boundstone),即是与原地生物群落构建相关的、特别多见于生物礁相的灰岩类型。Embry和Klovan(1971)依照生物作用类型对绑结岩做了进一步分类,次分为障积岩(bafflestone)、粘结岩(bindstone)。之后,Cuffey(1985)的礁灰岩分类中在绑结岩中引入盖覆岩(coverstone),即平板状生物体(如层孔虫)展布于生物骨架或碎片上加强礁灰岩的固结能力。粘结岩与盖覆岩相似,粘结生物群体整体形态上呈片状,但粘结生物本身形态或许具有不规则性(Cuffey, 1985)。障积岩中生物体形态一般呈枝状或块状(Embry and Klovan, 1971; Cuffey, 1985)。Insalaco(1998)及Riding(2002)分别基于造礁生物形态学、生长—绑结以及充填胶结方式对礁灰岩进行了更加详细的分类,但目前在生物礁古生态学和岩石学研究领域内尚未得以普遍推广使用。
雅水剖面旧司组顶部至上司组底部由下而上的礁滩灰岩微相类型以及特征见表1,微相实例则分别示于图3、图4和图5。自下而上描述如下。
表1黔南惠水雅水剖面上司组礁滩灰岩类型,微相类型,造礁方式和生物颗粒。表中A,B,C和D与图2中的相对应
Table1Limestonecatalogues,microfaciestypes,reefbuildingmodeandorganicparticlesofthereefandbankfaciesfromtheShangsiFormationattheYashuiSectionfromHuishui,SouthGuizhou.HorizonsoftheA,B,C,andDinthetable1correspondtothecolumnoftheFig.2

组灰岩类型微相类型造礁方式生物颗粒上司组滩相灰岩生物碎屑泥粒状灰岩无海百合茎,有孔虫,腕足类,藻类等礁相灰岩D丛管珊瑚障积岩;生物碎屑粒泥状灰岩、泥粒状灰岩丛管珊瑚障积丛管珊瑚,单体四射珊瑚,腕足类,腹足类,钙藻,介形类,海百合茎等C单体四射珊瑚障积岩;层孔虫盖覆岩单体四射珊瑚障积;层孔虫盖覆单体四射珊瑚,钙藻,层孔虫,腕足类等B笛管珊瑚障积岩;生物碎屑粒泥状灰岩笛管珊瑚障积;单体四射珊瑚障积单体四射珊瑚,笛管珊瑚,苔藓虫,介形类,腕足类,钙藻等A单体四射珊瑚障积岩;生物碎屑粒泥状灰岩单体四射珊瑚障积单体四射珊瑚,笛管珊瑚,海百合茎,腕足类,钙藻,介形类,有孔虫等滩相灰岩生物碎屑泥粒状灰岩无珊瑚,腕足类,介形类,海百合茎,钙藻,有孔虫,海胆等旧司组滩相灰岩生物碎屑粒泥状、泥粒状灰岩无腕足类,介形类,钙藻等
2.1 生屑滩(CYS-T)微相
CYS-T-1(图3a):生屑粒泥状灰岩、泥粒状灰岩。含腕足类、介形类、钙藻等碎片,分选性中等且多强烈破碎,大部分颗粒略有定向排列但未达支撑程度,颗粒重结晶明显;灰泥和粉砂屑充填,含微量石英细砂[样品号:CYS-a-1;薄片号:a-1-1(4)]。
CYS-T-2(图3b):生屑泥粒状灰岩、粒泥状灰岩。含海胆刺、腕足类、海绵骨针、有孔虫和钙藻碎片,局部见细条状生屑(多为珊瑚砂级碎片和海绵骨针)略呈包卷式围绕粗生屑排列,可能是定向涡旋水流造成的,大部分颗粒重结晶;颗粒支撑,之间充填灰泥[样品号:CYS-a-2;薄片号:a-2-1-4(3)]。
CYS-T-3(图3c):生屑泥粒状灰岩。含苔藓虫、钙藻、介形类、海百合茎、腕足类以及有孔虫等碎片,颗粒分选较差,除部分钙藻和介形类外,其它生屑强烈破碎,部分颗粒表面发生重结晶作用;生屑颗粒之间充填灰泥或粉砂屑,局部见少量亮晶方解石胶结[样品号:CYS-a-2;薄片号: a-2-1-3(6)]。
CYS-T-4(图3d):生屑泥粒状灰岩。含珊瑚、腕足类、介形类、海百合茎等碎片,多强烈破碎且分选性较低,部分砾级珊瑚碎片周围见粒度均匀、大小约0.2mm的椭圆形灰泥团粒聚集,其特征可类比路凤香和桑隆康(2002)讨论某些无脊椎动物(如蠕虫、腹足类等)的粪团粒成因;生屑颗粒之间充填灰泥,含少量亮晶方解石胶结[样品号:CYS-1层上部;薄片号:1层上部-2(1)]。
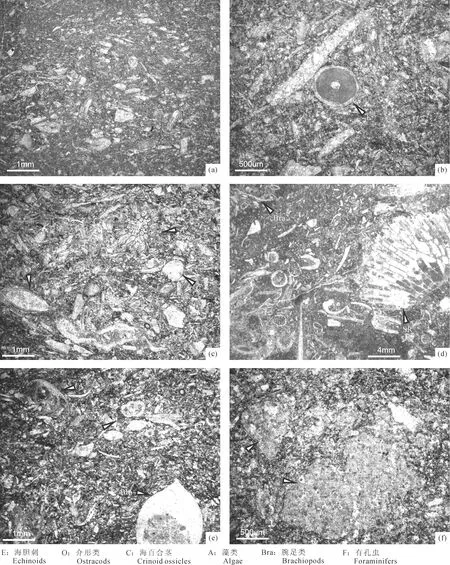
图3 黔南惠水雅水剖面旧司组顶部和上司组底部礁滩灰岩微相Fig.3 Microfacies in the reef and bank limestones from the top of the Jiusi Formation to the base of Shangsi Formation at the Yashui Section from Huishui, South Guizhou (a): a-1-1(4); (b): a-2-1-4(3); (c): a-2-1-3(6); (d): 1层上部-2(1); (e): 2a-2-1(顶)(3); (f): 2b-1-1(2)
2.2 珊瑚格架礁(CYS-J)微相
CYS-J-1(图3e):生屑泥粒状、粒泥状灰岩。含腕足类、钙藻、介形类、有孔虫等碎片,大部分颗粒分选较差并达支撑程度,除有孔虫和部分钙藻外,一般强烈破碎,部分在腕足类壳内见示顶底构造,见链状钙藻碎片,其形态与早古生代的Halysis相似,但这个属从未在晚古生代地层中报道过;颗粒之间充填灰泥,含少量亮晶方解石胶结[样品号:CYS-2a (顶);薄片号:2a-2-1(顶)(3)]。
CYS-J-2(图3f):生屑粒泥状灰岩。见细砾级重结晶的钙藻(可能为粗枝藻)碎片,颗粒分选性中等,因表面多强烈重结晶而难以判断种类;生屑颗粒间充填灰泥,局部见少量亮晶方解石胶结[样品号:CYS-2b-1;薄片号:2b-1-1(2)]。
CYS-J-3(图4a):单体四射珊瑚漂砾岩(Rudstone)。含单体四射珊瑚、横板珊瑚、海百合茎、腕足类等生物颗粒,这些生屑颗粒粒度偏粗,大者可达砾级,部分砾级单体四射珊瑚碎片周缘可局部障积灰泥、细砂级生屑、粉砂屑或细砾级生物碎片,单体四射珊瑚和横板珊瑚骨骼空腔中见示顶底构造;大部分生物颗粒间为灰泥和粉砂屑充填[样品号:CYS-2b-2;薄片号:2b-2-2(1)]。
CYS-J-4(图4b):生屑粒泥状灰岩。生屑颗粒和灰泥中见显著的生物扰动潜穴,呈椭圆形,其长轴倾斜或近水平,宽度变化较小,内部被极粗砂级海百合茎、腕足类等碎片以及细砂级生屑和粉砂屑充填,应为某类无脊椎动物的栖居穴;生屑间有大量粉砂屑和少量灰泥充填,可与潜穴周围的灰泥明显区分,潜穴周围聚集有富含有机质的暗色纺锤状球粒,粒度均匀,大小约0.2mm,由于该黑色颗粒边缘清晰,推测穴居生物离开后,其空间为某些无脊椎动物(如蠕虫、腹足类等)的粪团粒所充填[样品号:CYS-2b-3;薄片号:2b-3-3(1)]。
CYS-J-5(图4c):生屑粒泥状灰岩。含苔藓虫、腕足类、钙藻等生物颗粒,部分保存较好的苔藓虫可达砾级,但总体含量较少,不是生物格架岩的主要成分;生物颗粒之间充填灰泥,部分生物颗粒表面有重结晶[样品号:CYS-2b-4;薄片号:2b-4-1(8)]。
CYS-J-6(图4d):笛管珊瑚障积岩。原地生长保存的笛管珊瑚(Syringopora)空腔内充填亮晶方解石或灰泥,其枝体格架间障积介形类、腕足类、钙藻等生物颗粒,并充填细砂级生屑、灰泥和粉砂屑,部分颗粒表面重结晶[样品号:CYS-2b-4;薄片号:2b-4-2(2)]。
CYS-J-7(图4e):生屑粒泥状灰岩。具有明显不分枝的柱形生物潜穴,一般直或略弯曲,水平或倾斜,可互相交插排列,内无衬里,直径2~3.5mm,粗细变化较小,内被粗砂级苔藓虫、腕足类等生屑、细砂级生屑、粉砂屑和灰泥充填,充填物颗粒间见微亮晶方解石胶结,与潜穴周围的灰泥区分明显;推测这类生物潜穴可能是杨式溥等(2004)描记的(Planolite)漫游迹。与图4b中显示的遗迹化石相似,这一潜穴周围也聚集有纺锤状不透明的球粒,粒度均匀,大小约0.2mm,这类生物潜穴也属某些无脊椎动物在灰泥丰富沉积物中的栖居穴,之后被生物碎屑充填[样品号:CYS-2b-4;薄片号:2b-4-4(1)]。
CYS-J-8(图4f):单体四射珊瑚障积岩。单体四射珊瑚通过骨骼相互接触形成珊瑚障积岩,珊瑚空腔内被方解石胶结物和灰泥充填,之间见葛万藻(Girvanella)丝状体粘结在珊瑚外壁之上,可加固珊瑚骨架的抵抗能力[样品号:CYS-2b-5;薄片号:2b-5-1(3)]。
CYS-J-9(图5a):层孔虫盖覆岩。含复体四射珊瑚、层孔虫、腕足类等生物颗粒,见层孔虫利用砾级复体四射珊瑚碎片的硬底生长形成盖覆岩,这片盖覆岩显示原地保存,其边缘沉积物制约层孔虫侧向生长;生物颗粒之间充填灰泥,颗粒表面发生微弱重结晶[样品号:CYS-2b-5;薄片号:CYS-2b-5-3]。
CYS-J-10(图5b):生屑粒泥状、泥粒状灰岩。含复体和单体四射珊瑚、腕足类、腹足类、钙藻等生物颗粒,保存较好,可识别褶皱和光滑两种腕足类壳型,生物空腔内被灰泥或粉砂屑充填,在光滑壳腕足类空腔中的充填物具有明显粒序韵律,反映充填过程中的阶段性,腹足类壳内发育示顶底构造,但其所指方向与目前层序方向相反,可能是该腹足类壳体空腔被亮晶方解石充填后再次被搬运翻转后所致;大部分生屑未达到支撑程度,颗粒之间充填灰泥[样品号:CYS-3-3;薄片号:3-1-4-1(1)]。
CYS-J-11(图5c):丛管珊瑚障积岩。原地生长保存的丛管珊瑚(Siphonodendron)空腔间充填亮晶方解石,而枝体间障积丰富的灰泥,并有强烈破碎的珊瑚和腕足类等粗砂—砂级生物颗粒充填,生物颗粒被灰泥和方解石胶结物充填[样品号:CYS-3-1;薄片号:3-1-4-2(1)]。
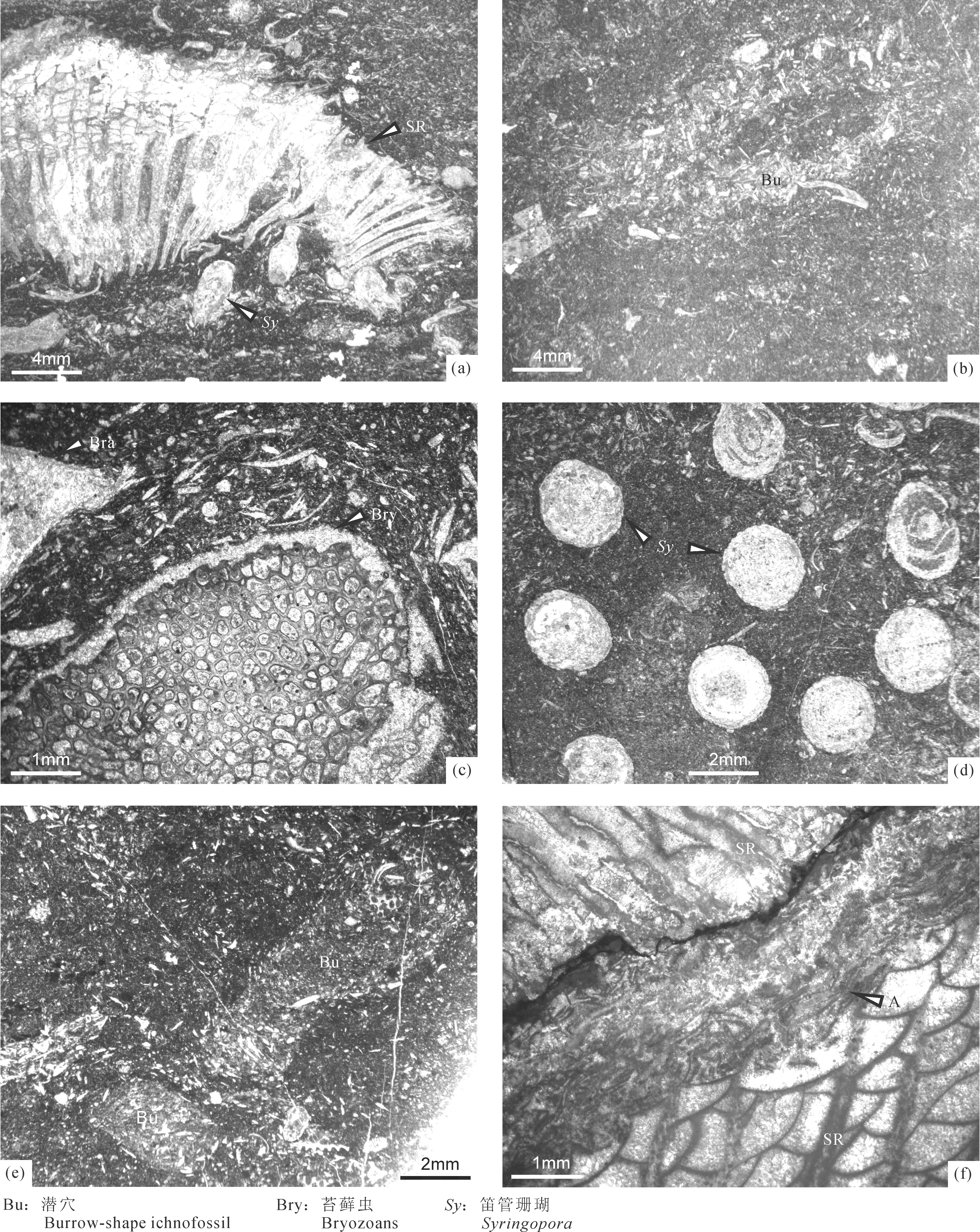
图4 黔南惠水雅水剖面上司组底部礁滩灰岩微相Fig.4 Microfacies in the reef and bank limestones at the base of Shangsi Formation at the Yashui Section from Huishui, South Guizhou(a): 2b-2-2(1); (b): 2b-3-3(1); (c): 2b-4-1(8); (d): 2b-4-2(2); (e): 2b-4-4(1); (f): 2b-5-1(3)
CYS-J-12(图5d):丛管珊瑚障积岩。原地生长保存较好的丛管珊瑚(Siphonodendron)空腔中充填灰泥和亮晶方解石,枝体间充填细砂级腕足类、粉砂屑和灰泥[样品号:CYS-3-3;薄片号:3-3-1(1)]。
CYS-J-13(图5e):丛管珊瑚障积岩。原地生长保存较好的丛管珊瑚(Siphonodendron)格架,在侧部以无性出芽繁殖方式生长出新的珊瑚个体,这样可以使珊瑚具有更大表面积,占据更大空间;局部珊瑚枝体折断,空腔中充填亮晶方解石和灰泥,枝体间充填大量灰泥,含微量的介形类、海百合茎、腕足类碎片和粉砂屑[样品号:CYS-3-3;薄片号:CYS-3-3-2]。
2.3 生屑滩微相
CYS-T-5(图5f):生屑泥粒状灰岩。含海百合茎,有孔虫,腕足类,藻类等碎片,除有孔虫外,一般强烈破碎,分选性中等,部分颗粒表面重结晶;颗粒之间被灰泥充填[样品号:CYS-4-1;薄片号:4-1-1(4)]。
3 珊瑚格架礁群落的历史定位和礁滩灰岩构建方式
泥盆纪弗拉期—法门期(F—F)生物大灭绝之前的中泥盆世大气CO2浓度是当今的14~24倍(Berner and Kothavala, 2001),超温室效应导致海洋中大量碳酸钙沉淀和钙质生物(珊瑚、层孔海绵、钙质微生物菌类和藻类等)高度繁盛,由这些生物造礁形成的礁相碳酸盐岩在全球广泛分布,涵盖北美、俄罗斯地台、西伯利亚地台、西欧、澳大利亚以及华南等陆块(Copper, 2002),主要为珊瑚—层孔虫海绵礁(Yu Changmin and Shen Jianwei, 1998; 王向东和沈建伟, 2004)。F—F事件期间出现的缺氧(Joachimski and Buggisch, 1993; Bond and Wignall, 2005)、海退(Sandberg et al., 2002; Chen Daizhao and Tucker, 2003)和降温(Copper, 1986; Averbuch et al., 2005; Chen Daizhao et al., 2005; Xu Bing et al., 2012)造成浅海区壳相底栖动物,特别是喜暖型珊瑚—层孔虫动物群急剧消失,而以叠层石或凝块石、Waulsortian泥丘、藻礁等原始类型群落呈现(Lees and Miller, 1985; Webb, 2002)。晚泥盆世—早石炭世经历了全球生物礁群落的复苏—重建过程(王向东和沈建伟, 2004),在杜内期无后生动物礁发育,随后以珊瑚—苔藓虫—海绵等为主的后生动物礁复苏和发展,并在维宪晚期达到繁盛阶段(Webb, 2002),与该期全球气候逐渐变暖相一致(Bruckschen et al., 1999; Mii et al., 1999; Isbell et al., 2003; Buggisch et al., 2008; Fielding et al., 2008; Grossman et al., 2008)。与全球早石炭世生物碳酸盐岩建造相对应,华南早石炭世以后生动物或藻类为主造礁形成的礁或礁丘,以及Waulsortian泥丘实例也十分罕见(巩恩普, 1997; 巩恩普等, 2010; Gong Enpu et al., 2012)。目前仅有广西柳州龙殿山杜内晚期Waulsortian相碳酸盐岩泥滩(Aretz et al., 2012),广西田林下垌维宪期珊瑚礁(陈晓红等, 2013)和甘垌子维宪期苔藓虫—珊瑚点礁(方少仙和侯方浩, 1986),以及广西来宾叠层石礁丘(Shen Jianwei and Qing Hairuo, 2010)报道。黔南维宪期上司组底部珊瑚格架礁的发育与全球早石炭世生物礁的演化可能具有同时性,反映了该期海洋处于温暖时期。
通过对黔南雅水下石炭统维宪阶上司组底部中的礁滩相碳酸盐岩建造的微相研究,可识别其沉积微相包括下列类型:
(1) 生屑滩有2种微相类型:① 泥粒状灰岩;② 粒泥状灰岩。
(2) 生屑滩之间的珊瑚格架礁有4种微相类型:①粒泥状灰岩;②泥粒状灰岩;③珊瑚障积岩;④层孔虫盖覆岩(表1)。
珊瑚格架礁中绑结岩以珊瑚障积岩为主,原地生长埋葬的珊瑚障积岩由丛状或块状近直立原地生长的单体四射珊瑚、笛管珊瑚(Syringopora)和丛管珊瑚(Siphonodendron)枝体与障积比例不等的灰泥、生屑或骨架相互支撑形成,可以使礁体具有更坚硬骨架而抵抗风浪。在珊瑚格架礁中还发育少量的层孔虫盖覆岩和藻类粘结作用,它们属华南石炭纪礁体中常见的微相类型(巩恩普, 1997)。珊瑚格架礁中的珊瑚一般以无性出芽生长方式繁殖(图5e),这是显生宙以来重要的礁体构建方式(Fagerstrom and West, 2011),在加勒比海活珊瑚礁中也起到重要作用(Foster et al., 2007)。这种无性出芽繁殖具有较高的生长速率,能够快速增加珊瑚体(Jackson et al., 1986),不仅快速地扩大格架岩体积以占据有利的空间生态位,还通过障积海底生物碎片和部分灰泥使礁体结构更加致密以增强抗浪能力(Fagerstrom and West, 2011)。珊瑚格架礁中的珊瑚障积岩、层孔虫盖覆岩、藻类粘结作用和珊瑚出芽生长方式的微相特征均昭示礁相特征。
生屑滩微相类型主要为生屑泥粒状灰岩,局部含少量生屑粒泥状灰岩(表1),珊瑚、腕足类、钙藻、介形类、海百合茎等生物颗粒破碎程度高而分选性低,据余素玉(1989)和Blomeier 等(2009)对生屑破碎程度和环境指标的识别依据,上司组的生屑滩为经历过水动力条件较强远距离或频繁搬运后的快速沉积产物,其所在的强水动能区堆积速度通常偏高,而这一地带短期堆积的颗粒在成岩固结之前被下一次风浪冲蚀搬运的几率也偏大,所以最终的净保存量也非异常偏大,在一定时限内此滩相堆积区是不适宜珊瑚定殖且达到大型格架生长阶段,仅见少量而单调的单体四射珊瑚和复体四射珊瑚。
珊瑚格架礁则以生物碎屑粒泥状灰岩、珊瑚障积岩为主,次为少量层孔虫盖覆岩,局部含少量生屑泥粒状灰岩(表1),腕足类、钙藻、有孔虫、海百合茎、介形类、腹足类、苔藓虫以及层孔虫多未达到支撑程度,这反映了珊瑚大量繁殖乃至达到造礁时的古环境重要指标,即格架岩生长与水动力条件减弱有关(Morin et al., 1994),另一重要的岩相特征显示,整个珊瑚密集生长的序列中鲜见颗粒灰岩,可佐证该剖面是大量生屑经频繁搬运后到达水动力条件降低的区域与灰泥共同堆积所在,无风暴级强水流冲蚀的海域为单体四射珊瑚、笛管珊瑚(Syringopora)和丛管珊瑚(Siphonodendron)群落发展创造了条件。在珊瑚格架礁之上的生屑滩中,有孔虫含量明显增加(图5f),而在礁相环境中很少,这说明有孔虫不是主要的附礁生物,而是滩相环境中的栖居者。
4 结论
黔南雅水早石炭世维宪期上司组珊瑚格架礁的发育与全球早石炭世生物礁的演化具有同时性,反映了该期浅海区处于温暖状态。该组底部礁滩灰岩的微相类型丰富,可划分出4种类型:① 粒泥状灰岩;② 泥粒状灰岩;③ 珊瑚障积岩;④ 层孔虫盖覆岩,其中,生屑滩有2种微相类型:① 泥粒状灰岩;② 粒泥状灰岩,生屑滩之间的珊瑚格架礁中有4种微相类型:① 粒泥状灰岩;② 泥粒状灰岩;③ 珊瑚障积岩;④ 层孔虫盖覆岩。
上司组中生屑滩以生物碎屑泥粒状灰岩为主,珊瑚、腕足类、钙藻、有孔虫、海百合茎等生物颗粒破碎程度大,分选性不好,为经历过水动力条件较强远距离或频繁搬运后的快速沉积产物,该环境不适宜珊瑚大量繁殖,仅见少量单体四射珊瑚和复体四射珊瑚。上司组中珊瑚格架礁则以生物碎屑粒泥状灰岩、珊瑚障积岩为主,发育少量层孔虫盖覆岩和藻类粘结作用,腕足类、钙藻、有孔虫、海百合茎、介形类、腹足类、苔藓虫以及层孔虫多未达到支撑程度,反映了水动力条件减弱,为单体四射珊瑚、笛管珊瑚(Syringopora)和丛管珊瑚(Siphonodendron)为主体的珊瑚群落形成原地礁灰岩创造了条件。
