埃迪卡拉纪印痕化石是陡山沱期宏体藻类的固着器吗?——Hiemalora与Gesinella的对比研究
2014-04-22唐烽金幸生王约丁明赵元龙高林志
唐烽, 金幸生, 王约,丁明, 赵元龙,高林志
1) 中国地质科学院地质研究所, 北京, 100037; 2) 国土资源部地层与古生物重点实验室, 北京, 100037; 3) 浙江省自然博物馆, 杭州, 310014; 4) 贵州大学资源与环境工程学院, 贵阳, 550003
内容提要: Hiemalora是一类广为分布的埃迪卡拉(Ediacaran)纪(震旦纪)圆盘状印痕化石,周围具流苏状或触须状突起,曾被解释为似水母动物,或某种叶状生物的固着器官,但以往罕见其与叶状生物体共同保存。本文报道了华南贵州震旦纪陡山沱组一类保存完好丰富的被确认为藻类的碳膜化石——革辛娜藻Gesinella,具有完整的须状固着构造。这一须状构造的形态和大小与Hiemalora非常相似,致密加厚的基部及其周围辐射状分布的丝状或须根状构造,可以分别和后者的中央盘以及周缘突起的“触须”相比较。陡山沱组的这一类碳膜化石同时保存有明显巨大的形同藻类的叶片和固着器,为研究埃迪卡拉生物群的底栖印痕化石提供了新的认识,即:盘状的Hiemalora可能是多细胞藻类的固着器官,而不一定是最初解释的似水母动物或其它叶状体生物的固着构造。不过,证实这一点尚需更进一步地研究埃迪卡拉纪生物的保存方式和埋藏环境。
我国南方埃迪卡拉(震旦)系(注:埃迪卡拉动物群,即Ediacara fauna, 音译为“伊迪卡拉动物群”更准确, 有广泛使用,但国家自然科学名词委员会1993年发布的《地质学名词》中收录的为“埃迪卡拉动物群”)陡山沱组页岩中,保存了以“庙河生物群”为代表的宏体碳质压膜化石群,以大量底栖固着的多细胞藻类为主,还包含推测的后生动物和海绵动物、可疑的遗迹化石等多门类生物化石,化石类型多样、分异显著,与国外埃迪卡拉纪多数保存为大型蕨叶状和圆盘状的印痕化石明显不同(唐烽和高林志, 1998; 唐烽等, 2008; Xiao Shuhai et al., 2002; Narbonne et al., 2005; Xiao Shuhai and Laflamme, 2009)。在近年研究的华南贵州江口的翁会剖面,埃迪卡拉系陡山沱组中、上部保存了较多的庙河生物群常见分子,比如:Baculiphyca、Miaohephyton、Enteromorphites、Doushantuophyton、Glomulus和Longifuniculum,以及数量众多的呈膜片状的Gesinella,被确认为底栖的多细胞藻类化石(赵元龙等, 2004),这些宏体碳质压膜化石均可见盘状、须根状保存的固着构造(王约和王训练, 2006; 唐烽等, 2008)。其中,Gesinella为厘米级以上的大型叶片状碳质压膜化石(唐烽等, 2008),叶状体呈不分枝的膜片状,长度可达14 cm以上,宽至2.4 cm以上。此类化石以往在湘西地区的埃迪卡拉系留茶坡组中也有很多发现(Steiner et al., 1992; Steiner, 1994; 陈孝红等,1999),被定名为湖南革辛娜藻Gesinellahunanensis(Steiner et al., 1992),国外中寒武世布尔吉斯页岩中也保存有类似的碳质压膜化石,但其具体的亲缘关系一直不能肯定(Briggs et al., 1994)。翁会新发现的革辛娜藻标本具有须根状的基部构造和叶状体分节的现象(Wang Yue et al., 2008; 唐烽等, 2008),据此可以推测这类化石可能属于与现代褐藻门中片藻类形态相近的多细胞藻类的祖先类型。其须根状的基部构造一般分为中央盘和外围辐射状须根突起两部分,形态上与国外(如:纽芬兰、白海和其它地区)埃迪卡拉纪典型的圆盘状印痕化石Hiemalora(Narbonne et al., 2005; Fedonkin et al., 2007)非常相似,表明这两类保存方式截然不同的宏体化石可能源于同一生物类别,Hiemalora很可能是底栖的叶片状高级藻类革辛娜藻的固着器官。
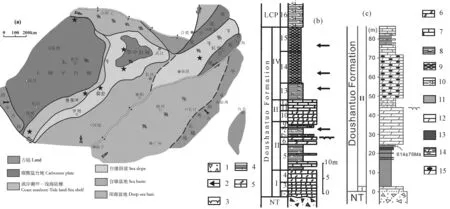
图1 (a) 华南震旦/埃迪卡拉纪岩相古地理略图(星号代表宏体化石群产地,据唐烽等,2009,图1略改);(b) 贵州翁会 剖面地层柱状图;(c) 华南湖北峡东宜昌樟村坪剖面地层柱状图(Liu Pengju et al., 2013,Fig. 7)Fig. 1 (a) Sketch map of the Sinian/Ediacaran facies and paleogeography in South China (modified from Tang Feng et al., 2009; Stars showing the locations of the typical macrofossil biota); (b) Stratigraphic column of Wenghui section, Jiangkou, Guizhou Province; (c) Stratigraphic column of Zhangcunping section, Yichang, Hubei Province (after Liu Pengju et al., 2013,Fig. 7) 陡山沱组中部发现华南多处可见的侵蚀间断面。1—冰碛岩;2—宏体化石层位;3—剥蚀面;4—砂质页岩;5—白云质灰岩;6—白云岩; 7—灰岩;8—钙质页岩;9—泥岩;10—硅质页岩;11—页岩;12—白云岩;13—磷块岩;14—泥质白云岩;15—含燧石结核白云岩 Note the erosional surface in the middle part of Doushantuo Formation.Legend: 1—tillite; 2—macrofossil horizons; 3—erosional surface; 4—silty shale; 5—dolomite limestone; 6—dolomite rock; 7—limestone; 8—calcareous shale; 9—mudstone; 10—siliceous shale; 11—shale; 12—dolomite rock; 13—phosphorite; 14—siltdolomite; 15—dolomite with chert nodule
贵州江口的“翁会生物群”产自陡山沱组中、上部的黑色泥岩及页岩中,埃迪卡拉/震旦纪翁会地区的古地理环境为台地斜坡至深水盆地的过渡地带(唐烽等,2009)。翁会剖面的陡山沱组厚约54 m,下伏Marinoan期的南沱组冰碛岩,上覆留茶坡组下部的深水相黑色硅质页岩(图1b)。自下而上大致分为Ⅰ—Ⅳ岩性层段(图1b),与三峡地区的陡山沱组四个岩性段基本可以对比:Ⅰ为1~4层,底部沉积盖帽碳酸盐岩,向上过渡为灰岩夹薄层页岩;Ⅱ为5~9层,厚层白云质灰岩与黑色页岩互层;Ⅲ为10~12层,灰岩及白云岩夹层;Ⅳ为13~15层,下部黑色钙质页岩,上部黑色页岩、泥岩。丰富多样的宏体藻类化石赋存在第8、13、14、15层中(图1b,箭头),其中占据优势的就是大型叶片状的革辛娜藻,基部保存有完整的须根状固着器,被推测为原位埋藏于松软富水的泥质基底,处于透光的浅水贫氧环境(Wang Yue et al., 2011)。在化石含量最为丰富的第13层,还发现有可能归属浮游的栉水母动物化石——八臂仙母虫Eoandromeda(Tang Feng et al., 2008, 2011; Wang Yue et al., 2008)和少量与国外埃迪卡拉纪圆盘状印痕化石相似的疑似后生动物的压膜化石。三峡地区陡山沱组庙河生物群产出层位的顶部年龄已经确认为551Ma(Condon et al., 2005),其中部第Ⅱ岩性段还普遍出露一个在华南区域分布的剥蚀间断面(Zhu Maoyan et al., 2007; Liu Pengju et al., 2009, 2013),年龄推断为600Ma (Barfod et al., 2002),在翁会剖面的第6层顶部也同样发现了这一侵蚀面(图1b),据此,可以初步推测上述含化石层的年龄大体为580~551Ma之间。
1 大型宏体藻类碳膜化石Gesinella
华南埃迪卡拉系陡山沱组的碳质压膜化石以湖北三峡地区的庙河生物群为代表,大多数都是叶片状、带状和丝状的宏体化石,与现生的底栖多细胞藻类的形态可以对比:上部为分枝或不分枝的叶状体,下部或基部是圆盘状或团块状的固着构造,多数连接有须根状的突起(Xiao Shuhai et al., 2002)。具有须根状突起的固着器、叶状体弯折和分节生长、侧分枝及二歧分枝等特征,表明这些碳质叶状体化石应该归属多细胞藻类(Xiao Shuhai et al., 2002; 唐烽等, 2008)。在贵州江口翁会剖面近年来新发现的大量藻类碳膜化石,叶状体和固着器同样保存得完好精美,丰富多样(赵元龙等, 2004; 王约等, 2007, 2009; 唐烽等, 2008)。依据目前已发现的化石材料来看,叶状体分枝的宏体藻类化石是新元古代埃迪卡拉纪陡山沱组地层中首次出现的类型,在湖北峡东的庙河生物群中占据优势;虽然叶片状藻类化石早在中元古代团山子组中就已经出现(Zhu Shixing and Chen Huiming, 1995),但是厘米级以上的大型叶片状藻类化石则在埃迪卡拉纪陡山沱组中首次出现,在贵州江口的翁会生物群中产出最为丰富(唐烽等, 2008; 本文图2,
图3)。尽管固着器出现了更多的类型,但是根据叶状体形态、大小相似的特征,产自翁会的这些碳膜化石均被鉴定为湖南革辛娜藻(Gesinellahunanensis) (赵元龙等, 2004; 王约等,2007; 唐烽等, 2008)。它们可能是迄今为止在晚前寒武纪地层中所发现的最大的宏体藻类化石,不仅具有巨大的不分枝的膜片状藻体,基部甚至还保存有相当清晰完整的须根状固着构造。
翁会的革辛娜藻化石新发现有三种类型的固着构造:一种呈矛尖状,为上部大型叶状体向基部逐渐缩小形成(图3c,d),大多数附着有密集的须根状突起(图3a,b,f,h),叶状体脱落后,二维保存的压膜标本往往呈小盘长须根的“星射状”固着器(如图3g,m,n)。这一类型的革辛娜藻化石最初由德国的Steiner 命名并图示记述,但其标本基部固着器仅有稀疏的须根突起(Steiner, 1994),与庙河生物群较为丰富的碳膜化石——棒形藻Baculiphycataeniata的固着器形态相似(图3o,q)。另一种为圆盘状或团块状,周缘未见任何突起物,其上通常直接连接较小的叶状体(图3e, i—l)。还有一种在翁会剖面保存较多(图 2a—m),侧压的叶状体在基部碳质明显加厚,基部放射状突起丝状、须状的附着物,与底栖藻类固着器的须根非常相似(图 2a—m;
图3d,h,m),在基部具有明显的压褶界线(如图2c,f—i),推测其直立固着于泥质基底时,中央形成直径较大的圆形或亚圆形的固着盘,四周放射状伸展出密集的须根状突起,呈中央大盘具射线的“日射状”固着器。后两种在翁会地区也曾发现有过渡类型(王约等,2007),代表该属藻类固着器可能存在连续的个体发育过程,即较小的藻类叶状体首先生长出圆盘或团块状固着器,类似吸盘的功能在基底吸附稳固后,再逐渐生长出密集的辐射状须根。而相似的固着器形态如果保存为印痕化石,可以说,与国外多地产出的埃迪卡拉纪圆盘状印痕化石Hiemalora(Narbonne et al., 2005; Hofmann et al., 2008)别无二致。

图2 (a)—(m):具须根状固着器的碳膜化石Gesinella Steiner, 1992Fig. 2 (a)—(m), Carbonaceous compressions of Gesinella Steiner, 1992 with filamenteous holdfasts 所有标本产自翁会剖面,收藏于浙江自然博物馆,编号依次为ZMNH M3361至M3374。标尺最小刻度为1 mm All specimens from Wenghui section, housed in the Zhejiang Museum of Natural History and numbered in turn for each ZMNH M3361 to M3374. Scale bar of minimum is 1mm
2 国外埃迪卡拉纪圆盘状印痕化石Hiemalora
世界各地的埃迪卡拉纪地层,比如:加拿大纽芬兰的Conception和St. John's群、南澳大利亚的Rawnsley石英岩、俄罗斯白海地区的Vendian系、北冰洋西伯利亚的Khatyspyt 组和纳米比亚南部的Nama群等,在其薄层的硅质碎屑岩或碳酸盐岩中都保存有丰富多样的大型印痕化石群,一般通称为“埃迪卡拉生物群”,地质年龄推断大约为580~541Ma (Hofmann et al., 2008)。除了Nama化石群以外,均发现有一类广泛分布的圆盘状化石——HiemaloraFedonkin, 1982,呈扁平的或略凹凸的中央盘及其周围的不规则辐射状突起(MacGabhann, 2007)。这类印痕化石在近30年来的相关研究中曾被广为提及或专文报道(Fedonkin, 1982, 1985, 1990, 1992; Vodanjuk, 1989; Runnegar and Fedonkin, 1992; Narbonne, 1998; Martin et al., 2000; Narbonne et al., 2001, 2005; Dzik, 2003; Serezhnikova, 2007; Hofmann et al., 2008),然而,其亲缘分类问题至今仍是一个难解之谜。Fedonkin (1982)最初发现并命名这类白海地区典型的埃迪卡拉纪化石时,曾归属为单体的水螅类,辐射状突起对比为水螅水母的触须;随后Anderson和Conway Morris (1982)的研究也将在纽芬兰Mistaken Point组发现的同类化石推定为某类具触须的水母状刺胞动物的体化石印痕,尽管他们也提出了Hiemalora可能还具有其它的两类亲缘关系,即归属某种生物的基部固着器,或某类星状的遗迹化石。后者遗迹化石的观点由Martin等(2000)重新评估过;其后不久,Dzik (2003)也认为Hiemalora化石可能为埃迪卡拉纪末期一类典型的底栖绝灭生物“Petalonamae”的固着盘;Serezhnikova (2007)则解释这个圆盘状化石为高级藻类或海绵动物的固着器;Hofmann等(2008)更将此类化石具体确定为一个固着器官属,化石本体是一个类似烛台状分枝的叶状体,连接有柄及Hiemalora式的固着构造,被命名为Primocandelabrum。
在纽芬兰的Mistaken Point砂岩中,呈放射状或触须状突起的Hiemalora标本通常具有一个近乎扁平的中央盘(直径0.3~4.8 cm之间,平均为2.25 cm),盘缘常见较窄的同心环状构造,连接窄环周围的是密集且长短不一的射线状突起物(最大可见长度 5.2 cm, 平均为 3.15 cm);这类射线一般细于2 mm,直线状或略弯曲,未见分枝,少见叠压,向外的末端变细尖灭(Hofmann et al., 2008)。由于在Mistaken Point组偶然发现了一个似烛台状分枝的叶状体与似Hiemalora的固着装置相连接的新属种——Primocandelabrumhiemaloranum,尽管其固着构造的中央盘与周围的长突起相比,明显较小,比纽芬兰已确认的Hiemalora化石形状也小很多,但Hofmann等(2008)仍认为该新类别与Hiemalora可能存在亲缘关系;正如同发现个别圆盘状的Aspidella和蕨叶状的Charniodiscus保存在一起以后 (Gehling et al., 2000),而改变了长期以来对Aspidella圆盘印痕化石的归属意见一样,这一发现也改变了最初的似水母生物的解释,将Hiemalora归于某类叶状生物的固着器官(Hofmann et al., 2008)。
基于西伯利亚北部Khorbusuonka群灰黑色薄层灰岩中产出的盘状Hiemalora印痕化石材料,Serezhnikova (2007, 2010)也提出了相似的观点,认为此类大型的半浮雕状印痕化石,可能归于海绵动物或两胚层动物级别(如海葵类)的底栖动物基部器官。不象纽芬兰的Hiemalora化石,有更大的中央盘和较长的须状物,甚至共同保存有固着柄和上部的叶状体,西伯利亚的同类化石未见叶状体连接,个体大小与华南翁会大型藻类的须根状固着构造较为相似或略大。2007年Serezhnikova的测量结果显示中央盘的直径在0.5~5.0 cm之间,须状物长度在0.5~3.5 cm之间。其重建的化石形态如同一个厚壁的海绵状底栖生物,具有锥状的固着器,着生有放射状长短不一的须状突起(Serezhnikova, 2007)。不过,她建议的Hiemalora原位保存的放射状突起物的功能,比如在海底软泥中固着及辅助吸收养分等(Serezhnikova, 2007),都显然与藻类的须根状固着器的功能一致。
3 对比和讨论
如前所述,陡山沱组的革辛娜藻化石很多都同时保存有直立的膜片状叶状体和须根状固着器,而固着器官相对较窄,但更富有碳质。据Steiner等(1992)和Steiner (1994)的最初解释,Gesinella藻早期发育时期叶状体基部可能呈管状或棒状的柄与圆盘状或须根状的固着构造相连接,翁会的标本都分别保存了这样的形态(图3a—c, e—g, i—l; 王约和王训练,2006)。而这种多细胞藻类的成体则更可能是具有囊状或膜片状的叶状体,基部连接着更加强壮、富含有机质的圆盘状固着器官,侧压保存的化石基部明显富集碳质(图2, 3),表明生活时的藻类基部有机质丰富,适于分泌胶质吸附在基底上;众多向外辐射的丝状须根或斜交、或垂直地连接于Gesinella叶状体基部的固着盘上(图2a—m),与Hiemalora印痕化石不规则的丝状附着物可以对比;基部的宽度代表这种藻类吸附在陡山沱期海底基质所形成的固着盘的直径,也基本等同于Hiemalora印痕化石中央盘周缘的圆环或圆脊的直径。尽管迄今还没有在页岩相中发现与典型的Hiemalora印痕化石形态(图3p; MacGabhann, 2007)直接对应的大圆盘具须根的碳膜化石,上述这些Gesinella基部构造与Hiemalora印痕化石较为一致的特征引发如下的推测:如果Gesinella直立的大型叶状体脱落,层面上留下的固着盘印痕的形态将与以往描述的Hiemalora中央圆盘及其周边射线状构造非常相似(Serezhnikova, 2007; Fedonkin et al., 2007; Hofmann et al., 2008; Grazhdankin et al., 2008)。
Hiemalora的辐射状“须根”可以划分为三个类型:其长度分别大于、等于、和小于中央盘的直径(Serezhnikova, 2007),相似的三分关系在陡山沱期Gesinella藻类须根状固着器标本中也存在。如同国外的Hiemalora印痕化石,华南翁会这一藻类压膜化石的大多数丝状须根,长度均大于叶片状基部的宽度,须根自基部边缘向外放射分布,至末端逐渐变细尖灭;而与印痕化石略有不同的是,藻类碳膜的须根未见可以确认的分枝,均为互相叠压形成的分叉。须根之间尚保存有很薄的碳质膜,有些碳膜还延展至须根的末端(图2c,f,i,m;
图3a,f—h,n)。这可能由于沉积环境不同的原因,陡山沱组黑色页岩代表的泥质基底更容易保存碳质膜;而印痕化石保存的多数硅质碎屑岩中有机质均淋滤消失。除了多数具较宽基部和边缘须根的固着器以外,翁会化石中尚有少部分基部呈锥尖状附着须根的Gesinella标本可能“扎根”在软沉积物中,一旦扁平膜状的叶状体脱落,层面上就有可能保存很小的中央盘和周围细长柔曲的放射状须根,如同上文所述的“星射状”(Steiner, 1994; 王约和王训练, 2006);湖北峡东陡山沱组的庙河生物群中同样产出小盘长须根的星射状化石,但多被确认为棒形藻Baculyphyca的固着构造(图3q;Xiao Shuhai et al., 2002);北美纽芬兰产出的烛台状或灌丛状Primocandelabrum印痕化石也连接有类似Heimalora的星射状固着器(Hofmann et al., 2008),表明叶状体形态不同的宏体化石可能具有形态相似的固着构造。
二维压膜的Gesinella藻类化石几乎都有厚实、碳质富集的基部,还有一部分连接着团块状、无须根、边缘平滑的椭圆状底盘(图3e,i—l),可能是曾被图示的须根生长之前的蒂状固着器(王约等, 2007)。如果上部的叶状体脱落,在层面上原位保存为压膜或印痕化石,与Heimalora略凹凸的中央盘印痕形态基本是一致的,甚至与埃迪卡拉纪广泛出现的圆盘状宏体化石的形态也可以对比。可以说,在晚前寒武纪冰期后以底栖叶状体多细胞生物占优势的埃迪卡拉生物群中,出现的圆盘状化石中至少有一部分可能属于藻类的固着器,生活于浅水透光带。
国外的埃迪卡拉生物群包含有大型蕨叶状的印痕化石,也发现有少数标本与圆盘状无放射纹的印痕化石共同保存,由此被推断为底栖的亲缘不明的后生生物(Gehling et al., 2000),而分布广泛、出露众多的圆盘状化石也有一部分被认为是软躯体底栖多细胞生物的固着构造(Fedonkin et al., 2007)。以庙河生物群为代表的华南埃迪卡拉系陡山沱组化石群中比较常见星射状或日射状如Heimalora印痕形态的碳膜标本,只是个体较小,还保存有更多的碳质膜,并多与Gesinella或Baculyphyca形态属的大型藻类叶状体基部连接保存,生活时显然具备固着的功能。而纽芬兰星射状的固着构造同样被确认为Heimalora,偶见其连接着较为模糊的烛台状或丛状的Primocandelabrum,这种扇状轮廓、具柄和分枝的叶状体印痕化石在纽芬兰地区比较罕见,与蕨叶状的印痕化石明显不同(Hofmann et al., 2008),而与华南庙河型生物群的分枝藻类叶状体形态则更为相似(Xiao Shuhai et al., 2002)。近年来,同样发现有硅质碎屑岩相的埃迪卡拉纪印痕化石的形态与页岩相庙河型碳膜保存的宏体化石相同(Eoandromeda, Tang Feng et al., 2008, 2011; Zhu Maoyan et al., 2008),表明庙河页岩型生物群尽管较为独特,地方性属种较多,但也存在与不同环境保存的埃迪卡拉生物群之间的交流,翁会地区陡山沱组化石层位之上即覆盖普遍被认为是深水相的留茶坡组硅质岩,可能与纽芬兰深水沉积的Avalon型化石群也存在一定联系。
纽芬兰Avalon半岛的埃迪卡拉系长期以来都被认为是属于透光带以下的深水浊流和前三角洲沉积(Narbonne et al., 2005; Hofmann et al., 2008; Wilby et al., 2011),然而其中产出的埃迪卡拉纪最早期的化石组合——Avalon生物群的空间辐射和类别分异度都可以媲美寒武纪“生命大爆炸”(Shen Bin et al., 2008),这与现代深海底栖生物组合低丰度、低分异度的现象不太吻合(Graham and Wilcox, 2000)。再者,Hiemalora形态的固着器在浅水相的华南地区与Gesinella的藻类叶状体可见较多的连接保存,而在深水环境的纽芬兰地区常见单独保存,罕见与分枝的Primocandelabrum叶状体印痕化石共同保存,这种固着器和叶状体低比例共同出露的现象,表明非原位保存、特异埋藏的可能性极大,似Hiemalora的固着器化石更可能来自略深水的透光带。依据现生底栖海藻的垂直分布规律,大型不分枝叶片状藻类(如Gesinella)更适宜生存在潮下带的较深水位(Graham and Wilcox, 2000),可能被浊流或三角洲前缘泥砂流瞬时搬运至斜坡—深水盆地的边缘过渡带,使膜片状或薄囊状的叶状体从固着构造上脱落、漂浮并腐解于远洋水体中,而更富集有机质的须根状固着器会更多保存在浊流或三角洲前缘砂质沉积中,相比在页岩相泥质沉积中基本保存为碳膜化石,这些砂质沉积中的固着器有机质更加容易被淋滤消失而形成印痕化石。南澳大利亚典型的浅水相印痕化石群(包含Rawnsley石英岩中Hiemalora式的圆盘状化石)多数就是保存在波基面下三角洲砂质沉积和风暴事件沉积中(Gehling et al., 2013)。其他浅水硅质碎屑岩相(如加拿大西北部和东欧地区)的埃迪卡拉纪沉积模式也同样易于保存半浮雕状的印痕化石群(Hofmann et al., 2008)。西伯利亚北部产出的Avalon型化石群也是保存于碳酸盐补偿面(CCS, carbonate compensating surface)之上的浅水瘤状灰岩薄层中(Grazhdankin et al., 2008),其中包含更多更精美的Hiemalora印痕化石。这些都表明,具有流苏状或须根状的圆盘印痕化石Hiemalora不仅出现在较深水的浊流和三角洲前缘的事件沉积,而且也出现在浅水的碳酸盐沉积中,与藻类富集生长的潮下透光带可能会有密切的联系。因而,将保存方式截然不同的两类化石,即碳膜化石的Gesinella藻类固着器和印痕化石的Hiemalora,进行对比研究后表明:Hiemalora印痕化石也许代表不同的生命类型,部分仍有可能解释为水母型的后生动物(Bykova, 2011),也有部分可能解释为分枝状生物Primocandelabrum的固着器(Hofmann et al., 2008),更有大部分可能归属于浅水透光带生活的膜囊状底栖多细胞藻类Gesinella的固着构造。印痕化石解释的局限性和不确定性需要在其它埋藏环境发现有更多更完美的形态相似的化石,和更加深入的沉积古地理对比研究。
4 结论
Hiemalora是典型的埃迪卡拉生物群印痕化石,曾被解释为似水母动物,现在解释为叶状体生物的固着器官。本文研究创新性地认为Hiemalora可能与贵州震旦纪陡山沱组碳质压膜保存的底栖藻类化石(革辛娜藻Gesinella)的固着器类似,并获得了以下几点新的认识:
(1) 尽管Hiemalora与Gesinella的固着器存在差别,正面保存的Hiemalora具有明显的内部环圈和外部放射状结构之分,而大多数侧面压膜保存的Gesinella固着器一般与藻体紧密相连,没有明显的内外之分;本文首次识别出叶状体归属Gesinella的三类固着器,其中的部分标本(如图示)同样可见中央盘和周围须根状突起的区分,若叶状体脱落后单独保存的话,形态上完全可以和Hiemalora对比;
(2) 在加拿大纽芬兰报道命名的,与Hiemalora连接保存的分枝状叶状体,保存得较为模糊,本文研究认为显然不是埃迪卡拉动物群中典型的蕨叶状分子,更可能与陡山沱期首现的分枝藻类叶状体相似;
(3) 虽然华南页岩相中的Gesinella叶状体是首次出现的最大的,但其固着器最大的也就几个厘米,而纽芬兰的Hiemalora印痕化石(连须根状突起物算上)大小不一,甚至有大到数十厘米的标本,但其平均值与西伯利亚北部记述的Hiemalora印痕标本类似,基本为5厘米左右,它们的平均大小都可以和Gesinella的固着器大小进行对比。
(4) 单独保存的Gesinella的固着器在华南包括翁会的化石点保存不多,在湖南桃源相近层位的黑色硅质页岩中出露有单独保存的Gesinella的固着器,并经Steiner (1994)素描图示,这首先表明Gesinella叶状体粗壮,基部有机质富集,不易脱离固着器,且页岩相环境沉积平稳缓慢,其中更易于保存连接叶状体的标本;其次更说明大小保存悬殊的Hiemalora印痕化石的埋藏环境是深水相的异地保存(如纽芬兰,Hofmann et al., 2008),叶状体容易脱落降解;或者是浅水相的事件沉积等(如南澳大利亚,Gehling et al., 2013),叶状体易于分离,而且事件沉积的较粗碎屑岩层中,更可能保存更大的同类底栖生物化石。
