高致病性猪繁殖与呼吸综合征病毒SHxx13/2013株的分离与鉴定
2014-04-13姜一峰虞凌雪李丽薇童光志周艳君
程 群,姜一峰,虞凌雪,王 康,杨 莘,李丽薇,高 飞,于 海,童 武,童光志,周艳君
(中国农业科学院上海兽医研究所,上海 200241)
高致病性猪繁殖与呼吸综合征病毒SHxx13/2013株的分离与鉴定
程 群,姜一峰,虞凌雪,王 康,杨 莘,李丽薇,高 飞,于 海,童 武,童光志,周艳君
(中国农业科学院上海兽医研究所,上海 200241)
2013年初上海某猪场保育猪群中出现高热、呼吸障碍等临床表现,导致大部分发病猪死亡,为了确定此次猪群中爆发疫病的病原,本研究从发病猪群中随机采集了15份血液样品,利用RT-PCR方法对本次疫病病原进行了检测。结果显示,有11份临床样品呈现猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory syndrome virus, PRRSV)强阳性,且与高致病性猪繁殖与呼吸综合征病毒(Highly pathogenic Porcine reproductive and respiratory syndrome virus, HP-PRRSV)对照大小相一致。随后从阳性样品中选取SHxx13/2013在Marc-145细胞中进行病毒的分离与鉴定,结果显示SHxx13/2013在细胞接种后第1代即出现明显的细胞效应(cytopathic effect, CPE),其病变特征与高致病性PRRSV强毒HuN4株一致。对SHxx13/2013第5代细胞分离毒进行RT-PCR和IFA等特异性鉴定,结果显示该毒株为PRRSV分离株,其蚀斑形态和生长特性与强毒HuN4株相似。分段克隆该毒株全长基因进行测序和序列分析,结果显示,新分离的流行毒SHxx13/2013株全长基因组为15 319 bp,其Nsp2基因特征与HP-PRRSV一致,在第482位和第534~562位发生两处共30个氨基酸的不连续缺失,且该毒株与高致病性PRRSV代表毒株HuN4亲缘关系较近,全长基因的核苷酸同源性为99.6%。本研究结果表明新分离的SHxx13/2013株属于高致病性PRRSV分离株,据此我们推测今年年初上海某猪场爆发的疫情主要是由HP-PRRSV感染所致。
高致病性猪繁殖与呼吸综合征病毒;PCR;分离;鉴定
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种以妊娠母猪流产、死胎、弱仔、木乃伊胎等繁殖障碍及仔猪的呼吸道疾病为特征的传染病[1]。1987年美国首次报道该病发生,随后在世界各国陆续爆发和流行。中国在2006年以前PRRS主要是呈地方流行或散发,其流行的毒株主要是以北美洲型CH-1a为代表的PRRSV经典毒株,然而2006年PRRSV在全国范围内出现爆发和广泛流行。此次流行是由基因组发生变异PRRSV感染造成的,被命名为高致病性PRRSV,其临床特征主要是高热、高发病率和高死亡率。变异的PRRSV(例如HuN4株与JXwn06株等)[2,3]与经典PRRSV(例如CH-1a株、VR2332株等)相比,致病性明显增强,在其体外传代细胞Marc-145上的适应性强,在基因组的非结构蛋白Nsp2基因中存在不连续的30个氨基酸的缺失[4-6]。已有研究证实Nsp2基因中存在30个氨基酸的缺失与病毒毒力无关[3],究竟是何原因导致变异的PRRSV致病性增强目前尚无结论。对于高致病PRRSV的预防和控制主要是通过疫苗免疫,近几年来,随着一系列高致病性PRRSV弱毒疫苗株的面世和临床应用,使由PRRSV变异株引起的疫情在一定程度上得到了有效地控制[7],但不容忽视的是还有一些地区仍然存在PRRS的散发和地方流行。2013年年初上海市某猪场保育猪群中部分猪出现了发热、呼吸困难、厌食消瘦和死亡等症状,为了持续关注PRRSV目前的流行动态,本研究从该发病猪场中采集了部分发病猪的临床样品,对其进行PRRSV等相关病原的检测、分离和鉴定,对该猪场的疫情控制提供了有力依据。
1 材料和方法
1.1 病毒、细胞与样品来源PRRSV HuN4株、HuN4-F112株、Marc-145细胞以及抗PRRSV N蛋白特异性单抗均由本研究室保存[8],15份发病猪的血液样品采集自上海某发病猪场。
1.2 主要试剂RNA提取试剂盒RNeasy Mini Kit 购自QIAGEN公司;单链cDNA合成试剂盒RevertAidTMFirst Strand cDNA Synthesis Kit 购自Fermentas 公司;pMD18-T 载体、LATaqDNA 聚合酶、DNA Marker、dNTP 均购自TaKaRa 公司;DH5α、DNA提取试剂盒Tissue/Cell gDNA Mini Kit、胶回收小量试剂盒购自上海华舜生物技术有限公司;FITC标记的山羊抗小鼠IgG二抗购自Sigma公司;胎牛血清、胰蛋白酶、DMEM 培养基购于Gibco公司。
1.3 临床样品核酸提取及检测处理本实验室采集的发病猪的血液样品并分别将其命名为SHxx1/2013--SHxx15/2013,按照RNeasy Mini Kit和Tissue/Cell gDNA Mini Kit说明书操作,分别提取15份样品的RNA和DNA,RNA按照单链cDNA合成试剂盒RevertAid First Strand cDNA Synthesis Kit说明书进行反转录,然后分别以合成的cDNA和DNA为模板,利用实验室已有的PRRSV、猪瘟病毒(Classical swine fever virus, CSFV)、乙型脑炎病毒(Japanese encephalitis virus, JEV)、猪流行性腹泻病毒(Porcine epidemic diarrhea virus, PEDV)、伪狂犬病毒(Pseudorabies virus, PRV)、猪圆环病毒(Porcine cireovirus, PCV)和猪细小病毒(Porcine parvovirus, PPV)等特异性鉴定引物进行PCR鉴定,获得的PCR产物经1%琼脂糖凝胶电泳观察。
1.4 病毒的分离将经RT-PCR检测为PRRSV阳性的样品经0.22 μm微孔滤膜过滤除菌处理,取无菌处理后的样品500 μL接种预先制备好的Marc-145单层细胞中,37℃感作1 h,然后加入含有2%的 FBS的DMEM维持液,在37℃、5%的CO2细胞培养箱中培养,观察细胞病变效应(cytopathic effect, CPE),同时进行病毒传代,分离病毒,分装后于-80℃保存备用。
1.5 分离病毒的特异性鉴定取分离传代后第五代病毒储存液提取核酸,对其进行部分重要猪源病毒的特异性PCR鉴定。同时利用抗PRRSV N蛋白特异性单抗对分离病毒进行间接免疫荧光特异性鉴定。
1.6 病毒生长曲线的绘制测定第五代分离病毒的TCID50,然后按0.01 MOI的剂量接种Marc-145细胞,分别于12、24、36、48、60 h收取细胞上清液,绘制病毒生长曲线,同时设强毒HuN4株和疫苗毒HuN4-F112株对照。
1.7 病毒蚀斑形态观察将第五代分离病毒按0.1 MOI接种于预先在六孔板中准备好的单层Marc-145细胞,经低熔点琼脂糖凝胶培养与结晶紫染色后,观察病毒的噬斑形态,同时设强毒HuN4株和疫苗毒HuN4-F112株对照。
1.8 病毒全长基因扩增及序列分析提取第五代病毒的RNA,利用文献报道的方法对其全长基因进行分段扩增、克隆和测序[5],从GenBank中选择具有代表性的PRRSV参考株基因组序列利用MEGA5.0软件进行比较分析。
2 结果
2.1 样品检测结果利用实验室已有的PRRSV、CSFV、JEV、PEDV、PRV、PCV和PPV等特异性鉴定引物对15份临床样品进行PCR鉴定,结果显示15份临床样品中有11份呈PRRSV阳性,其余病原检测结果均为阴性(图略),且PRRSV阳性样品其PCR产物大小与PRRSV变异缺失株大小相一致(图1)。
2.2 病毒分离选择阳性样品SHxx13/2013接种Marc-145细胞,结果显示在接种后72 h后即可观察到明显的细胞病变。其细胞病变主要表现为细胞聚集成簇、圆缩,部分细胞脱落,其CPE形态与PRRSV HuN4强毒株对照相似,不同于疫苗毒HuN4-F112株引起的细胞病变(图2)。
2.3 分离病毒的特异性鉴定将SHxx13/2013的第5代病毒进行RT-PCR鉴定,结果显示仅PRRSV特异性引物能够扩增出约800 bp的特异性目的片段,而其他猪源病毒检测结果均为阴性。进一步利用抗PRRSV N蛋白特异性单抗进行IFA检测,结果显示SHxx13/2013的第5代病毒感染的Marc-145细胞在荧光显微下能够观察到明显的特异性荧光(图3)。
2.4 病毒生长特性鉴定利用多步法绘制SHxx13/2013-F5生长曲线结果显示,SHxx13/2013-F5在Marc-145细胞上的增殖能力低于弱毒HuN4-F112株,而与强毒HuN4株在Marc-145细胞上的增殖能力相近(图4A)。对SHxx13/2013-F5的噬斑形态观察结果显示,在相同感染条件下,新分离的SHxx13/2013株其蚀斑形态与强毒HuN4株的比较接近,不同于HuN4-F112株感染产生的噬斑形态(图4B)。
2.5 病毒全长基因扩增将SHxx13/2013-F5病毒全长基因分为8个片段后进行PCR扩增,结果显示得到的8个片段均与预期大小相一致,其预期扩增长度依次为1057、2016、1771、1902、2263、2569、3486、3776 bp(图5)。

图1 PRRSV Nsp2鉴定引物RT-PCR检测结果Fig.1 RT-PCR products of PRRSV identif cation diagnostic primers

图2 PRRSV不同毒株感染Marc-145细胞病变效应观察Fig.2 Cytopathogenic effects on Marc-145 cells infected with different PRRSV strains
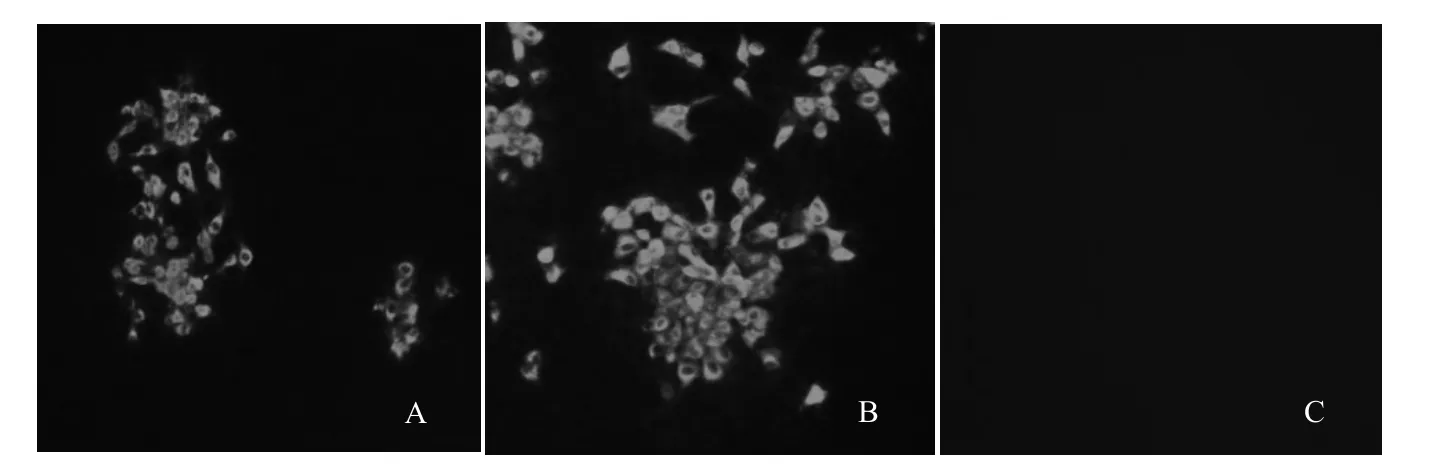
图3 SHxx13/2013的第5代病毒IFA特异性鉴定Fig.3 Identif cation of SHxx13/2013-F5 by immunof uorescence assay (IFA)
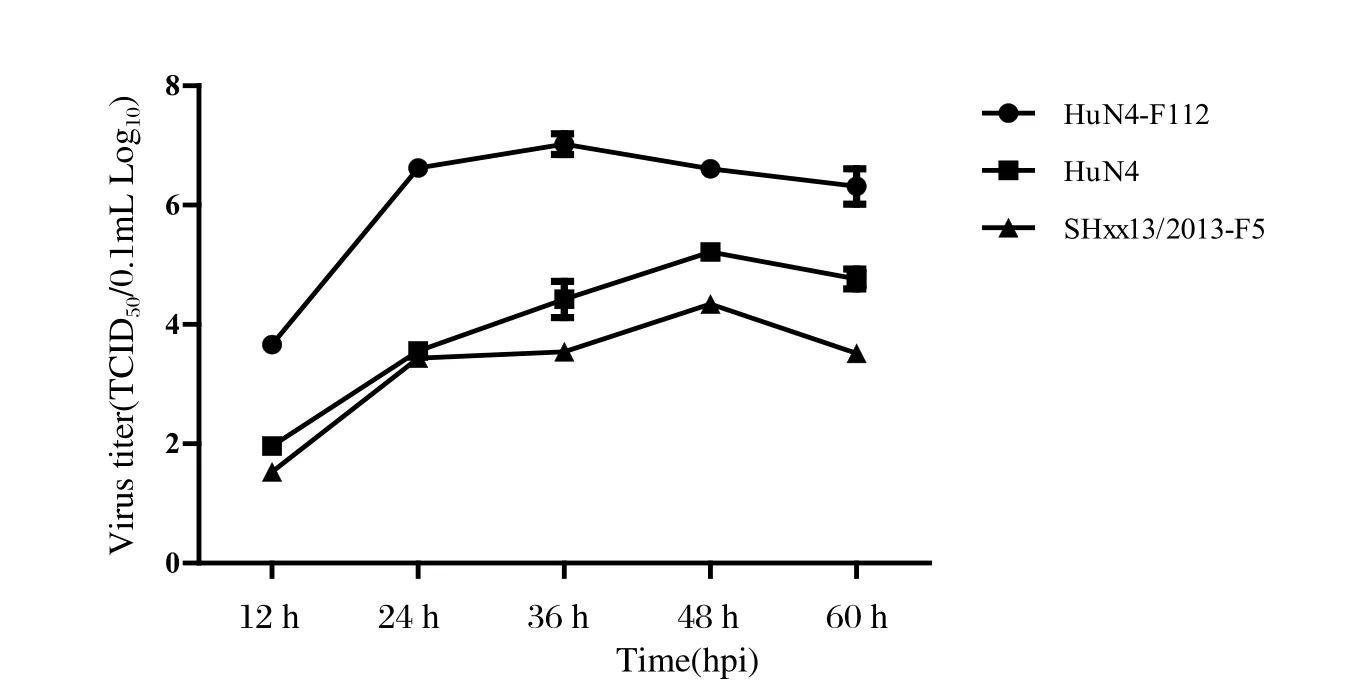
图4A SHxx13/2013-F5与HuN4、HuN4-F112株生长曲线Fig.4A Growth kinetics of PRRSV SHxx13/2013-F5, HuN4 and HuN4-F112

图4B SHxx13/2013-F5与HuN4、HuN4-F112噬斑形态Fig.4B The plaque morphology of SHxx13/2013, HuN4 and HuN4-F112

图5 SHxx13/2013株全长基因RT-PCR扩增结果Fig.5 RT-PCR products of SHxx13/2013 amplif ed by complete genome primers
2.6 SHxx13/2013全长基因序列分析将SHxx13/2013毒株8个基因片段测序拼接后,结果显示SHxx13/2013毒株全长基因为15 319 bp。利用MEGA5.05软件对SHxx13/2013株及参考株序列比较,结果显示新分离的SHxx13/2013株其Nsp2基因与高致病性PRRSV参考株同样在第482位和第534~562位发生两处共30个氨基酸的不连续缺失,主要结构蛋白GP5基因与高致病性PRRSV参考株氨基酸相似性为97.2%~98.0%,绘制的全长基因系统发生进化树结果显示,新分离的SHxx13/2013株属于北美洲型分离株,与2006年以后的高致病性PRRSV参考株有较近的亲缘关系(图6)。
3 讨论
猪繁殖与呼吸综合征是对全球养猪产业危害较为严重的传染病[9,10]。2006年高致病性PRRSV的爆发,曾一度给我国养猪业造成了巨大的经济损失。PRRSV 分离株分为北美洲型和欧洲型2种基因型,目前我国流行的PRRSV毒株主要是北美洲型毒株[11,12],在我国由于2006年高致病性猪蓝耳病的爆发,又习惯的将其分为经典毒株(其代表株为CH-1a株、HB-2(sh)/2002株等)和高致病性毒株(代表株有HuN4株、JXwn06株等)。二者之间在病毒基因水平存在的典型特征是Nsp2基因在第482位和第534~562位发生两处共30个氨基酸的不连续缺失,虽然现在已经证实该缺失与PRRSV病毒高致病性之间不存在必然联系,但此缺失仍被认为是区分中国高致病性PRRSV和其他美洲型毒株的标志。现在我国猪群中流行的PRRSV主要为缺失变异的高致病性PRRSV毒株[13]。
本研究从2013年年初上海某发病猪场采集临床样品,进行猪源病毒检测,结果表明在发病猪临床样品中仅检测到了PRRSV的存在,样品中PRRSV阳性率为73.3%。对其中阳性样品进行病毒分离后,经过RT-PCR、IFA等特异性鉴定,证实我们分离获得了一株PRRSV流行毒SHxx13/2013,该分离株对传代细胞Marc-145表现高度易感,第一代即可出现具有明显特征性的CPE。对流行毒SHxx13/2013生物学特性鉴定结果表明,该毒株在Marc-145细胞上的增殖特性与高致病性PRRSV强毒HuN4株相似,且形成蚀斑的形态也与强毒HuN4株基本相一致,与疫苗毒HuN4-F112株的生长特性相差较大。推测新分离的SHxx13/2013株并非是疫苗毒感染,对发病猪场进行背景调查结果证实该发病猪场一直未使用PRRSV疫苗免疫猪群。我们对新分离的SHxx13/2013株进行全长基因测序分析,结果显示该毒株Nsp2基因在第482位和第534~562位发生两处共30个氨基酸的不连续缺失,且与高致病性PRRSV缺失变异株具有较近的亲缘关系,证实新分离的SHxx13/2013株属于高致病性PRRSV变异毒株。综合上述研究我们分析认为该猪场由于对保育仔猪群未及时进行疫苗免疫,一定程度上增加了感染高致病性PRRSV的几率,进而导致在保育仔猪群中发生猪蓝耳病疫情,并在猪群中迅速传染和蔓延,造成了大批保育猪的发病和死亡。本研究结果提示我们目前尽管高致病蓝耳病的爆发流行处于较低水平,但是仍需要密切关注该病的流行和遗传变异趋势,不能放松针对该病的科学诊断、预防和控制。

图6 基于SHxx13/2013株全长基因进化树分析Fig.6 Phylogenetic tree of complete genome of SHxx13/2013
[1] Dea S, Bilodeau R, Sauvageau R,et al. Outbreaks in Quebec pig farms of respiratory and reproductive problems associated with encephalomyocarditis virus[J]. J Vet Diagn Invest, 1991, 3(4)∶ 275-282.
[2] Zhou Y J, Hao X F, Tian Z J,et al. Highly virulent porcine reproductive and respiratory syndrome virus emerged in China[J]. Transbound Emerg Dis, 2008, 55(3-4)∶ 152-64.
[3] Zhou L, Zhang J L, Zeng J W,et al. The 30-aminoacid deletion in the Nsp2 of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China is not related to its virulence[J]. J Virol, 2009, 83(10)∶ 5156-5167.
[4] 童光志, 周艳君, 郝晓芳, 等. 致病性猪繁殖与呼吸综合征病毒的分离鉴定及其分子流行病学分析[J]. 中国预防兽医学报, 2007, 29(5)∶ 323-327.
[5] Murtaugh M P, Elam M R, Kakach L T. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus[J]. Arch Virol, 1995,140(8)∶ 1451-1460.
[6] 郭宝清, 陈章水, 刘文兴. 从疑似PRRS流产胎儿分离PRRSV的研究[J]. 中国畜禽传染病, 1996, (2), 1-4.
[7] Zhou L, Yang H C. Porcine reproductive and respiratory syndrome in China[J]. Virus Res, 2010, 154(1-2)∶ 31-37.
[8] Zhou Y J, An T Q, Liu J X,et al. Identification of a conserved epitope cluster in N protein of porcine reproductive and respiratory syndrome virus[J]. Viral Immunology, 2006, 19(3)∶ 383-390.
[9] Garner M G, Whan I F, Gard G P,et al. The expected economic impact of selected exotic diseases on the pig industry of Australia[J]. Rev Sci Tech, 2001, 20(3)∶ 671-685.
[10] Neumann E J, Kliebenstein J B, Johnson C D,etal. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States[J]. J Am Vet Med Assoc, 2005, 227(3)∶ 385-392.
[11] Nelsen C J, Murtaugh M P, Faaberg K S. Porcine reproductive and respiratory syndrome virus comparison∶divergent evolution on two continents[J]. J Virol, 1999, 73(1)∶ 270-280.
[12] Zhou Y J, Yu H, Tian Z J,et al. Genetic diversity of the ORF5 gene of porcine reproductive and respiratory syndrome virus isolates in China from 2006 to 2008[J]. Virus Res, 2009, 144(1-2)∶ 136-144.
CHARACTERIZATION OF RECENTLY ISOLATED SHXX13/2013 STRAIN OF HGHLY PATHOGENIC PORCINE REPRODUCTIVE AND RESPIRATORY SYNDROME VIRUS
CHENG Qun, JIANG Yi-feng, YU Ling-xue, WANG Kang, YANG Shen, LI Li-wei, GAO Fei, YU Hai, TONG Wu, TONG Guang-zhi, ZHOU Yan-jun
(Shanghai Veterinary Research Institute, CAAS, Shanghai 200241, China)
In early 2013, an acute disease characterized by high fever and respiratory problems in piglets outbroke on a pig farm in Shanghai and caused many deaths. To investigate the causative agent, a total of 15 serum samples were collected and viruses were examined in RT-PCR. Eleven out of of 15 clinical samples showed strong Porcine reproductire and respiratory syndrone virus (PRRSV) signal and the same PCR fragments as compared with highly pathogenic PRRSV (HP-PRRSV). One PRRSV strain was isolated from a PCR positive sample and designated as SHxx13/2013 strain. The SHxx13/2013 strain induced cytopathic effect (CPE) on MarC-145 cells at the f rst passage, which shared similar plaque morphology and growth kinetics with HuN4 strain. The complete genome of SHxx13/2013 strain was 15 319 bp. The deletion of total 30 amino acids occurred at positions 482 and 534-562 in Nsp2 gene, which was a common feature of HP-PRRSV. The complete genome of the SHxx13/2013 strain was 99.6% identity to that of HuN4 strain, indicating that it was close to HuN4. The results concluded that HP-PRRSV was the causative agent in pig population in Shanghai.
Highly pathogenic Porcine reproductive and respiratory syndrome virus (HP-PRRSV); PCR; isolation; identif cation
S852.659.6
A
1674-6422(2014)02-0032-07
2014-01-14
国家自然科学基金项目(31100121);863计划项目(2011AA10A208-2);上海市重点基础研究项目(11JC1415200);国家生猪现代产业技术体系建设项目(NYCYTX-009);上海市科技兴农重点攻关项目(沪农科攻字(2012)第2-5号)
程群,女,硕士研究生,预防兽医学专业
童光志,E-mail:gztong@shvri.ac.cn;周艳君, E-mail:yjzhou@shvri.ac.cn
