光萼女娄菜种子成熟时期对萌发特性的影响
2014-04-08魏玲玲何青山杨慧玲
魏玲玲, 何青山, 杨慧玲
(河南农业大学生命科学学院,河南 郑州 450002)
在自然条件下,有些植物的种子同时(或在较窄的时间范围内)成熟,如中间锦鸡儿(Caraganaintermedia)、老芒麦(Elymussibiricus)[1,2];而另一些植物的种子成熟时间分布在较长的时间范围内,如白沙蒿(Artemisiasphaerocephala)[3].植物种子成熟的时间格局直接影响种子大小和休眠萌发特性[4],对植物生活史策略有重要影响,在种群的维持和更新过程中具有关键作用.因此,种子成熟和散布时间被认为是植物对环境的适应方式,在植物生活史对策和生态适应机制方面具有重要的研究价值[5].在不同时间成熟的种子所经历的环境条件不同,这些环境因素会对种子生物学特性产生重要影响.环境因素[6](如温度、光周期、光照强度、土壤水分和养分等)与植株内在因素(如生长不同阶段的营养物质分配、激素调节和果实位置等)相互作用,从而决定种子重量、大小、种皮厚度和种子活力.GUTTERMAN等[7]研究发现,在种子成熟过程中的光照周期长短是决定豆科植物OnonissiculaGuss.种皮透水性的关键因素:随着光周期由长到短,种皮的吸水速率由快到慢.同时,不同成熟时间的种子在母株上成熟所经历的不同环境会决定种子萌发和休眠特性的差异.这种影响已经在多个物种中得到证实,如马齿苋(Portulacaoleracea)[8],美国风铃草(CampanulaAmericana)[9]和一些灌木树种[10].此外,母株的繁殖物候也通过影响种子外在和内在(如种子内部营养含量)因素而影响萌发行为[9].种子萌发与休眠特性决定种子的萌发时间和下一代的命运[6],因此不同成熟时间引起的种子休眠与萌发特性的分化会分散植物种群内个体之间种子在短时间内大量萌发的风险,是植物适应异质性生境的一种策略.光萼女娄菜(MelandriumfirmumSieb. et Zucc. Rohrb.)又名坚硬女娄菜或粗壮女娄菜,为1年生或2年生草本,花期在05-07月份,果期在08-09月份.光萼女娄菜株高40~100 cm,花集生于茎顶及叶腋部,假轮伞状间断式总状花序,花瓣白色或粉红色;蒴果长卵形,长8~9 mm;种子呈肾形,长约1 mm,黑褐色;生于林缘草甸、山地草甸及灌丛间,在我国北部有广泛的分布[11,12].目前对光萼女娄菜的研究相对较少,主要研究集中其在药理药性方面[13]和植物分类学[14]方面,有关光萼女娄菜种子成熟的时间格局,特别是不同成熟时间的种子的萌发特性研究至今未见报道.光萼女娄菜具有长达5个月的花果期,引起植株之间甚至同一植株不同部位之间种子在成熟时间上产生很大的差异,这种差异可能对该物种适应异质性生境具有重要意义.因此,为了解不同成熟时间对光萼女娄菜种子生物学特性的影响,本研究对不同时期成熟的光萼女娄菜种子大小和吸水特性以及不同时期成熟的光萼女娄菜种子休眠和萌发特性对环境因子的响应进行了研究.
1 材料与方法
1.1种子的采集
本研究所用的光萼女娄菜种子采集于鄂尔多斯国家草地生态系统国家野外科学观测研究站(39°29′ N, 110°11′ E),海拔1 300 m.研究站位于毛乌素沙地的东南部,毛乌素沙地是中国四大沙地之一.该地区主要是固定沙丘和流动沙丘组成的沙地生态系统,年平均降水量345.8 mm,降水集中在05-09月份.
种子采集日期为2013-08-03,08-10,08-20,08-30,09-10,09-20和09-30,种子成熟的标志为果实上孔裂出现.从光萼女娄菜第1批种子成熟开始(08-03),到整株枯黄结束(09-30),共计7次.种子采集时摘取已经枯黄并且开始散布种子的果实(一般位于植株顶端),将种子从果实中轻轻倒出,确保不损坏植株其它部分.采种所用光萼女娄菜植株总数大于50株,将每株上已经成熟的种子全部收集.自第1次采种之后,对每株植株进行标记,以便下次在同株上继续采种.将同一时间采集的种子清理杂质,进行以下试验.
1.2种子大小的测定
随机选取不同时间成熟的光萼女娄菜种子各200粒,使用1/10000电子天平(Sartorius BP 221 S, Sartorius AG, Gottingen, Germany)测定重量,4个重复,测得的重量换算成种子千粒重.
1.3种子吸水特性
种子吸水特性试验在室内(气温18~25 ℃,相对湿度20 %~40 %)进行.将100粒种子置于盛有2.5 mL蒸馏水且垫有1层滤纸(Whatman No.1)的5 cm培养皿后开始计时,分别在吸水0.5,1,2,5,6,8,10,12,24,48,60 h后将种子取出,吸水纸吸干、称重,再放回培养皿中,如此重复,直至种子萌发,试验设3个重复.为了测定种子的吸水曲线,按下列公式计算种子吸水重量增加率Wr:
Wr=[(Wt-Wi)/Wi]×100
式中:Wi为种子原重量,Wt为种子吸水t时间后的种子重量.
1.4种子萌发测定
将不同日期成熟的种子分别置于直径为50 mm的培养皿中,每个处理4个重复,每重复25粒种子.培养皿垫两层滤纸(Whatman No.1),加2.5 mL蒸馏水.为防止水分蒸发,将培养皿置于自封袋中,于培养箱(江南仪器GXZ-0328)中进行萌发试验.
1.4.1 温度和光照对种子萌发的影响 试验的温度梯度为:5/15,10/20,15/25,20/30,25/35 ℃ (温度对应12 h黑暗/12 h光照).其中,5/15 ℃ 代表光萼女娄菜自然生境中4月和10月的气温,10/20 ℃代表5月和9月的气温,15/25 ℃代表6月和8月的气温,20/30 ℃代表7月中上旬的气温,25/35 ℃代表7月下旬的气温.光照条件为12 h黑暗/12 h光照和全黑暗.全黑暗处理则是将种子培养皿放于暗袋中.12 h黑暗/12 h光照条件的种子每天统计萌发率,以胚根萌出1 mm为萌发标志,将萌发的种子从培养皿移除,共统计20 d;全黑暗条件下的种子只在试验结束时统计最终萌发率.
1.4.2 划破种皮对种子萌发的影响 确保不损伤胚根的前提下,在胚根萌出部位将不同时期成熟的光萼女娄菜种子的种皮划破.在15/25 ℃、12 h黑暗/12 h光照的条件下进行20 d的萌发试验.
1.4.3 赤霉素(GA3)对种子萌发的影响 将不同时期成熟的光萼女娄菜种子置于2.5 mL不同浓度(0.1,0.01 mmol·L-1)处理的GA3溶液中连续萌发20 d,以蒸馏水为对照.采用15/25 ℃和12 h黑暗/12 h光照的条件进行萌发试验.
1.5数据处理
在统计分析之前,对数据进行平方根和反正弦转换,以保证方差齐性.采用单因素、双因素和3因素方差分析比较,各处理之间的差异(5 %水平);如果差异显著,再采用Tukey’s HSD方法进行多重比较.通过统计软件SPSS 17.0进行数据分析.
2 结果与分析
2.1种子千粒重
成熟时间对光萼女娄菜种子千粒重影响显著(F=77.199,P<0.001).种子千粒重随着成熟时间的延后而逐渐增加,最大值出现在09-20(0.192 6 g) ,为最小值(08-03)的1.29倍;而后下降(图1).9月末种子千粒重下降的可能原因是温度降低,未成熟种子提前成熟所致.

数据为平均值±标准误差,下同.不同字母表示不同成熟时间种子之间的千粒重差异显著(P<0.01).
Data are mean±SE. The same as below. Different letters indicate significant differences between different seed ripening times(P<0.01).
图1成熟时间对种子大小的影响
Fig.1Effectofdifferentripeningtimesonseedmass
2.2种子的吸水特性
光萼女娄菜种皮具有透水性,随着吸水时间的增加,种子吸水重量也随之增加(图2).09-30成熟的种子重量增加率最大(达54%),而09-20最小(22.37%).09-30和08-30在吸水后48 h开始萌发,而其他种子在吸水后60 h开始萌发(图2).不同时期成熟种子的吸水曲线一致,在0~10 h为种子吸水快速期,20~70 h为种子吸水的慢速期.
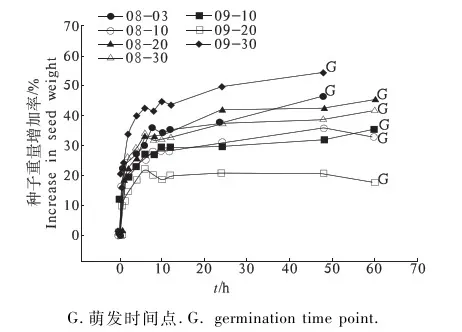
G.萌发时间点.G. germination time point.
2.3种子萌发测定
2.3.1 温度和光照对种子萌发的影响 3因素方差分析表明,光萼女娄菜种子萌发率受成熟时间(F=74.979,P<0.001)、温度(F=56.873,P<0.001)和光照(F=517.579,P<0.001)条件的显著影响,并且其交互作用也显著(P<0.001).
不同成熟时间的种子对温度和光照条件有不同的响应格局(图3,图中不同的大写字母表示相同成熟时间的种子在不同温度处理之间的萌发率差异显著;不同的小写字母表示在相同温度下,不同成熟时间的种子之间的萌发率差异显著,P<0.05).随着成熟时间的延后,种子萌发率总体下降.在光照条件下,种子萌发的最适温度范围在10~30 ℃.在10/20 ℃条件下,08-03成熟的种子萌发率达到100 %.黑暗条件则对种子萌发有强烈的抑制作用,仅早期成熟的种子(08-03和08-10)有少量的萌发.
2.3.2 种皮划破对种子萌发的影响 双因素方差分析表明,划破种皮对光萼女娄菜的种子萌发率有显著影响(F=609.245,P<0.001),划破种皮的种子萌发率显著高于未划破种皮的种子,但对于08-03成熟的种子2种处理间差异不显著.划破种皮的种子萌发率均在67 %以上,未划破种皮种子萌发率的平均值仅为30% (图4,图中不同的大写字母表示相同种皮处理下不同成熟时间的种子萌发率显著差异;不同的小写字母表示不同种皮处理下相同成熟时间的种子萌发率显著差异,P<0.01).
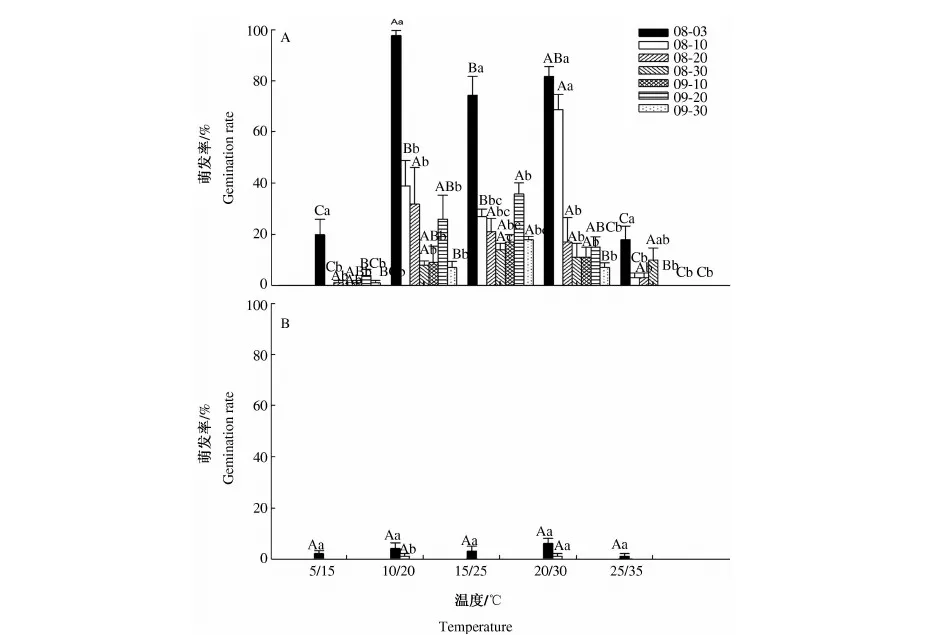
图3在光照(A)和黑暗(B)条件下,不同成熟时间的光萼女娄菜种子在不同温度下的萌发率
Fig.3Effectofripeningtime,temperatureandlightconditionsonGerminationofseedsofMelandriumfirmumthatmaturedatdifferenttimesatdifferenttemperatureregimesunderlight(A)anddark(B)conditions

图4划破种皮对不同时期成熟的光萼女娄菜种子萌发率的影响
Fig.4Effectofseedscarificationongerminationrateofseedsfromdifferentripeningtimes
2.3.3 GA3处理对种子萌发的影响 双因素方差分析表明,成熟时间(F=81.648,P<0.001)和GA3浓度(F=8.989,P<0.001)对光萼女娄菜种子萌发率有显著影响,两者交互作用影响也显著(F=2.743,P=0.001).
同一成熟时间条件下,GA3处理对08-03, 08-10, 08-20, 08-30, 09-10成熟的种子萌发率没有显著影响.然而,GA3处理显著降低了09-20和09-30成熟的种子萌发率.在相同GA3浓度处理下,08-03成熟的种子萌发率均比其他时间成熟的种子萌发率高(图5,图中不同大写字母表示相同成熟时间的种子在不同GA3浓度之间萌发率显著差异;不同小写字母表示相同GA3浓度下,不同成熟时间的种子萌发率差异,P<0.01).
3 结论与讨论
种子成熟时的环境因素(如日照长度、温度、光质和光强等)对许多植物的休眠水平有重要影响.光萼女娄菜种子的发育时间长短存在很大差异,不同时间成熟的种子所经历的环境因素各不相同,因而在种子特性上可能存在较大差异.本研究的结果证实,不同成熟时间的光萼女娄菜种子在形态、萌发与休眠特性方面存在很大差异,这种差异性可能是光萼女娄菜适应异质性生境的有效机制.

图5GA3对光萼女娄菜种子不同时期成熟萌发率的影响
Fig.5EffectofGA3treatmentongerminationofseedsfromdifferentripeningtimes
对于同一物种来说,种子大小的差异存在于不同的年际间,不同的地理种群间以及不同个体间,甚至存在于同一株植物中.光萼女娄菜先成熟的种子在初夏开花(05-06月),夏季成熟(08月),由于夏季炎热,日照长,土壤水分蒸发量大,对种子生物量的积累是不利的,所以早期成熟的种子千粒重低;后成熟的种子在夏季开花(07-08月),秋季成熟(09月),由于秋季气温适宜,昼夜温差大,而且土壤水分不易蒸发,对种子生物量的积累有利,所以后期成熟的种子千粒重增加.但是,进入深秋(09月底),光萼女娄菜整株迅速枯黄并死亡,生长代谢急剧减缓至停止,导致09-30成熟的种子发育时间过短,物质积累不充分,种子千粒重有所下降.另外,种子在植株上的位置也会对生长发育有影响[15].种子成熟的位置是从植株顶端到基部,养分和光合产物的运输方向是由主茎到末梢,种子离主茎越远,种子越小[16].光萼女娄菜先成熟的种子位于植株顶端,因此植株位置效应可能也是不同成熟时间的种子之间差异的重要原因.
光萼女娄菜种子千粒重与吸水量存在负相关,质量越小的种子,种子吸水重量增加率越大(图2).这一现象可能与以下2点原因有关:第1,小种子内所含亲水物质所占比例比大种子的可能要多;第2,在不同时间成熟的种子中,大种子的种皮发育更充分,具有更加厚重致密的种皮结构(自身不亲水),且致密的种皮不易使种子内部水分散失,所以大种子的含水量较高;而小种子种皮发育不充分,亲水的种胚在种子质量当中所占比例更大,且种子内部的水分在种子成熟后易透过种皮散失,所以大种子的含水量较低.
虽然种皮具有透水性,但是种皮依然是光萼女娄菜种子萌发的限制因素之一(图4),即种皮的机械结构依然阻碍种胚突破种皮.20 d的GA3处理并不能使种子萌发率有所提升(图5),说明种子需要通过更长更复杂的生理后熟作用(需要经历春季的低温层积作用)才能打破其生理休眠[17].
不同时间成熟的光萼女娄菜种子的萌发与休眠特性不同,这与种子发育过程中所经历的外界环境条件(如温度、降水和日照长度等)密切相关[4].首先,光萼女娄菜先成熟的种子萌发率高于晚成熟的种子(图3),这可能是因为先成熟的种子发育时外界的温度较高.这一结果与一些已有研究相类似.例如,MEYER等[18]发现,从温度低的生境中采集Artemisiatridentata种子中在15 ℃具有更深的休眠,而从温度高的生境中采集的该种子不具有休眠.其次,光萼女娄菜种子先成熟的种子和晚成熟的种子所经历的日照长度也不相同.GUTTEERMAN[19]发现,马齿苋(Portulacaoleracea)母株发育时的光周期影响种子的萌发率,种子在短日照下发育时比长日照下发育具有更高的萌发率.这与我们研究结果相反,说明日照长度可能不是解释2者种子萌发率差异的主要原因.最后,光萼女娄菜种子先成熟的种子萌发率高于晚成熟的种子,可能与2种种子发育时不同的营养物质分配有关.种子在不同成熟时间的发育过程中所处环境条件的差异会引起种子在萌发和休眠特性上产生差异.GALLOWAY[9]对美国风铃草(CampanulaAmericana)的研究也得出相似的结论:早成熟的种子比晚成熟的种子萌发率更高,因为2者的内含物和化学成分不同.光萼女娄菜种子的不同成熟时间格局和生理休眠特性决定了种子萌发对环境的温度和光照的响应格局(图3-A,图3-B).如果土壤水分充足,较早(08-03,08-10和08-20)成熟的光萼女娄菜种子可以在8月和9月份的温度条件下大量萌发.而9月之后成熟的种子,则不会在9月和10月的温度条件下大量萌发.在温带内陆气候下,植物需要经历漫长而寒冷的冬季,如果种子在成熟当年随着降雨出现而萌发,那么种子必须形成足够抵御严寒的植株体来度过冬季.较早成熟并萌发的光萼女娄菜的种子有相对较长的生长时间和较适宜的温度来进行幼苗的建成;而较晚成熟的种子如果萌发,则可能不会在秋季低温等环境条件下形成有效过冬的植株.光萼女娄菜的成熟时间和萌发对温度的响应格局,有效地降低了种子的消耗.
光萼女娄菜种子萌发对光照条件敏感,在完全黑暗的条件下几乎不能萌发(图3-B),因此散落到土壤深层的种子不会大量萌发,具备形成持久的土壤种子库的潜力.另外,光萼女娄菜的果实为蒴果,呈长卵形,开口向上,果实中的种子到第2年才可以完全散布,因此,光萼女娄菜又具有气生种子库.土壤种子库和气生种子库能够将光萼女娄菜种子萌发在时间和空间上分散,有效降低了种子1次性大量萌发的风险,是种群在严酷多变的环境中有效的适应策略.
光萼女娄菜通过种子成熟的时间格局调节种子的休眠萌发特性,进而影响植株的生活史周期,为种群的维持和更新提供了多种的选择.
参考文献:
[1]陈河龙,王 赞,孙桂芝. 中间锦鸡儿结实习性[J].草业科学,2007,24(2):20-25.
[2]韩建国,毛培胜,牛忠联,等. 老芒麦种子发育过程中的生理生化变化[J].草地学报,2000,8(4):237-244.
[3]YANG X, DONG M, HUANG Z. Role of mucilage in the germination ofArtemisiasphaerocephala(Asteraceae) achenes exposed to osmotic stress and salinity[J]. Plant Physiology and Biochemistry, 2010,48(2-3):131-135.
[4]KEBLAWY E A, RAWAI A A. Effects of seed maturation time and dry storage on light and temperature requirements during germination in invasiveProsopisjuliflora[J].Flora,2006,201(2): 135-143.
[5]TEVIS L J R. Germination and growth of ephemerals induced by sprinkling a sandy desert[J].Ecology,1958,39(4):681-688.
[6]BASKIN C C, BASKIN J M. Seeds: Ecology, biogeography and evolution of dormancy and germination[M]. San Diego: Academic Press, 1998.
[7]GUTTERMAN Y, EVENARI M. The influence of day length on seed coat colour, an index of water permeability, of the desert annualOnonissiculaGuss[J]. Journal of Ecology, 1972,60(3):713-719.
[8]GUTTERMAN Y. The influence of the photoperiodic regime and red-far red light treatments ofPortulacaoleraceaL. plants on the germinability of their seeds[J]. Oecologia, 1974, 17: 27-38.
[9]GALLOWAY L F. The effect of maternal phenology on offspring characters in the herbaceous plantCampanulaAmericana[J]. Journal of Ecology. 2002,90:851-858.
[10] FENNER M. Seeds: the ecology of regeneration in plant communities [M]. 2nd ed. Wallingford UK: CABI publishing, 1992.27-59.
[11] 《内蒙古植物志》编辑委员会. 内蒙古植物志 [M]. 呼和浩特:内蒙古人民出版社,1985.
[12] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社,1996.
[13] 徐富一,池鲜英,李恩淑,等. 硬叶女娄菜止痛、消炎及对关节炎效能的研究[J].河南医学院学报,2003,18(1):31-32.
[14] 赵一之,马 平. 内蒙古女娄菜属(石竹科)二新种[J]. 植物分类学报,1989,27(3):225-227.
[15] GUTTERMAN Y. Survival adaptations and strategies of annuals occurring in the Judean and Negev Deserts of Israel[J]. Israel Journal of Plant Sciences,2002,50(3):165-175.
[16] SUSKO D J, LOVETT-DOUST L. Patterns of seed mass variation and their effects on seedling traits inAlliariapetiolata(Brassicaceae)[J].American Journal of Botany,2000,87(1):56-66.
[17] BASKIN J M, Baskin C C. A classification system for seed dormancy[J]. Seed Science Research,2004,14(1):1-16.
[18] MEYER S E, MONSEN S B, MCARTHUR E D. Germination response ofArtemisiatridentata(Asteraceae) to light and chill: patterns of between population variation[J]. Botany Gazette, 1990,151: 176-183.
[19] DORNE A J. Variation in seed germination inhibition ofChenopodiumbonus-henricusin relation to altitude of plant growth[J]. Canadian Journal of Botany,1981,59: 1893-1901.
