东北天然次生林生物量对不同森林经营策略的响应
2014-03-26戎建涛何友均梁星云
戎建涛,何友均,梁星云
(1 温州科技职业学院,浙江 温州 325006; 2 中国林业科学研究院a资源信息研究所,b林业科技信息研究所,c 森林生态环境与保护研究所,北京 100091)
大气中CO2日益增加的趋势已引起世界各国的广泛关注[1],碳汇研究已成为当前的热点[2-3]。森林作为陆地上最大的生态系统,其碳储量约占陆地生态系统碳储量的57%。森林生物量和生长量是森林生态系统结构功能的基础,也决定和制约着水源涵养、土壤保持、气候改善等效益,并在维持生态平衡、改善气候等方面发挥着不可替代的作用[4]。生物量测定也是研究许多林业和生态问题的基础。
东北地区是我国木材生产的重要基地,由于历史上滥砍滥伐、经营不善等诸多因素,大面积天然林顶级群落退化为天然次生林。诸多学者在东北地区开展了生物量研究。如李文华等[5]以长白山地区天然林为研究对象,概述了主要生态系统生物量的研究方法;刘世荣等[6]对21年生兴安落叶松(LarixgmeliniRuor)群落的生物量、生物量增量、枯死量、凋落量进行了测定研究,获得群落结构初级生产力;国庆喜等[7]利用TM图像和一类清查数据构建了多元回归方程和神经网络模型,并用其估测了小兴安岭地区的森林生物量;李晓娜等[8]以东北天然次生林主要乔木树种为研究对象,模拟出地上、地下生物量方程,分析了林木生物量与个体生长的关系。然而,有关不同经营模式下天然次生林生物量的研究还鲜见报道。为此,本研究分析了粗放经营、目标树经营、无干扰3种经营模式下天然次生林的生物量差异,探讨不同经营模式对森林生态系统生物量的影响,以期为多功能森林经营提供参考依据。
1 材料与方法
1.1 研究区概述
研究区位于小兴安岭南坡的黑龙江省哈尔滨市林业局丹清河林场内(129°11′~129°25′E,46°31′~ 46°39′N)。该地区属于中温带大陆性季风气候,夏季气候湿润,降雨量集中在7和8月份,年降雨量600 mm左右,年蒸发量1 250 mm。年日照时数 2 200 h,年平均气温2 ℃,最高气温31 ℃,最低气温-35 ℃。初霜在9月末,终霜在5月中旬,无霜期120 d左右;始冻期为10月初,解冻期为5月初。地貌类型属于低山丘陵,海拔190~1 028 m,平均海拔500 m左右。土壤类型以暗棕壤为主,分布于不同的坡向和坡位,占该林场土地面积的94.9%。
1998年以来,丹清河林场开始探索目标树经营的道路。针对结构比较正常、有培养前途的林分开展了目标树经营;针对低质、退化的天然次生林,在保持原有生态系统的前提下,按照“模仿自然规律,加速发育进程”的原则,通过林分调整法、下层木补植和抚育法等人为干扰措施,将其转化成具有自育机制的、高价值的永久性多功能森林。人工作业的目的是促进高价值目标树的回归、出现和生长,酌情节制非目的树种的发育,伐除已经走向贬值阶段的立木资源,提升整体森林质量,追求均衡的生态经济价值[9-10]。
本研究选取丹清河林场的针叶林、阔叶林和针阔混交林3种森林类型,分别研究粗放经营、目标树经营和无干扰对其生物量的影响。不同经营模式的经营目标不同,因而采取的经营措施也不相同。
(1)粗放经营模式。对成过熟林以木材收获为主要目标,采用皆伐或“拔大毛”式择伐(采大留小、采好留坏),采伐强度大约40%左右;对中幼龄林进行抚育,抚育方式主要为透光抚育、生长抚育,抚育强度在30%左右;更新方式采取天然更新或者破坏了更新基础而无法更新。
(2)目标树经营模式。①针叶林:以乡土树种为主要经营对象,主要选择具有培育价值的树种红松作为目标树;标记目标树并对其进行单株抚育管理,注重修枝操作;根据目标树体系采取强度不超过30%的抚育间伐,伐除影响目的树种生长的非目的树种和生长过密、生长不良以及感染病虫害的病腐木,每5年进行1次抚育间伐;充分利用自然能力促进更新,同时辅以人工补植的森林更新方式。②阔叶林:针对低质低效的天然次生阔叶林,调整林分结构,通过多目标经营实现正向演替;对于密度过大且质量低下的蒙古栎和糠椴进行适当采伐,合理调整林分密度,为林下更新创造条件,每5年进行1次抚育间伐,同时将剩下的优质蒙古栎作为目标树;补植水曲柳、胡桃楸等珍贵树种,形成异龄、复层、多树种林分结构。③针阔混交林:选取红松作为目标树经营,对于影响目标树经营的非目的树种进行砍除;适当伐除材质较差的臭冷杉和没有培养前途的紫椴,每5年进行1次抚育间伐;利用自然力和人工辅助更新的方式,合理清除影响林木更新的灌木和草本;补植红松、水曲柳等优质、珍贵树种,提高优质树种的比例和林木生长量。
(3)无干扰模式。以封护、自然生长为主,没有采伐和抚育,主要依靠天然更新。
不同经营模式下天然次生林的林分概况见表1。

表 1 不同经营模式下东北天然次生林的林分概况
1.2 研究方法
1.2.1 样地设置与调查 2011-08-2012-04,在黑龙江省哈尔滨市林业局丹清河林场内,选择针叶林、阔叶林和针阔混交林3种森林类型,每种森林类型选择立地条件、树种组成和林龄基本一致,经营模式分别对应粗放经营、目标树经营和无干扰的林分。在每种研究对象林分内设20 m×20 m的样地10块,总共90块样地,对样地内胸径大于1 cm的林木进行编号并展开每木调查,记录其种名、胸径、树高和冠幅。在每块样地内沿对角线设置3块2 m×2 m的小样方,调查小样方内灌木层和草本层的植物种类、个体数、高度和覆盖度等,同时收集样方内的凋落物(包括未分解和半分解凋落物)现存量。

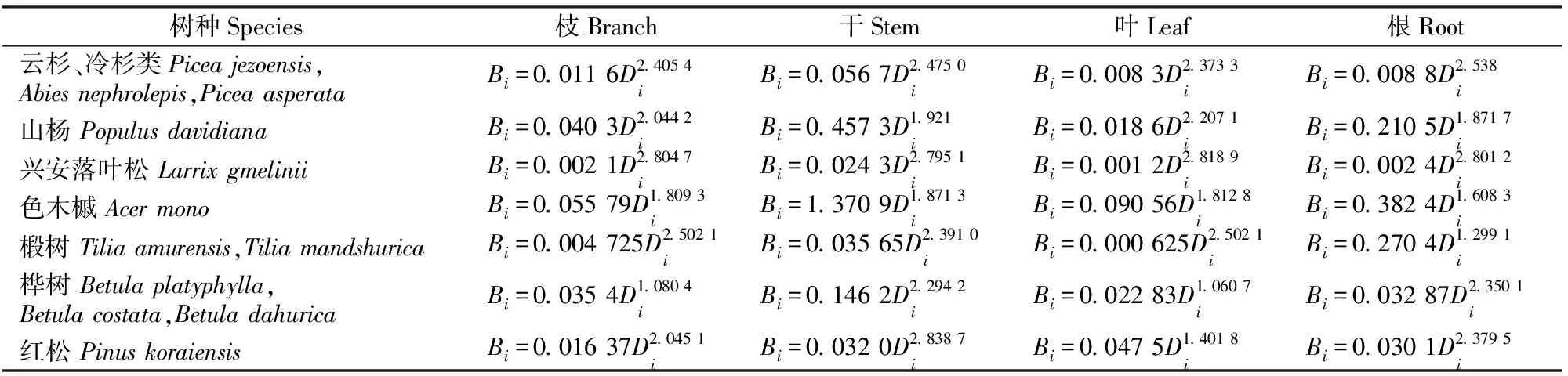
表 2 东北地区主要树种枝、干、叶、根的生物量模型
1.2.3 数据处理与统计分析 所有数据均利用SPSS 16.0软件进行统计分析,利用单因素方差分析进行差异显著性检验,对差异显著的数据进行多重比较(P<0.05,LSD,t检验)。
2 结果与分析
2.1 不同经营模式下3种森林类型乔木层生物量及其在各器官的分配
由表3可知,针叶林、阔叶林、针阔混交林在粗放经营、目标树经营、无干扰3种经营模式下林分乔木总生物量分别为79.79,174.60,117.68;96.29,159.38,106.79;105.96,163.55,141.80 t/hm2。在林分乔木总生物量组成中,干材生物量最大,占总生物量的46.52%~68.32%,比例最大的是阔叶林目标树经营模式,比例最小的是针叶林粗放经营模式。其他器官生物量所占比例依次为根(12.23%~38.07%)>枝(6.51%~17.93%)>叶(2.28%~4.45%),其中阔叶林粗放经营模式根和叶所占比例最小。乔木地上部分生物量占总生物量的比例为61.93%~87.78%,地下部分占 12.23%~34.79%。植物各器官生物量的空间配置状况在很大程度上反映了植物对光能、水分和养分的利用效率[15]。

表 3 东北地区不同经营模式下3种森林类型乔木层生物量及其分配
在针叶林、阔叶林和针阔混交林中,乔木总生物量排序均为目标树经营>无干扰>粗放经营。针叶林和针阔混交林乔木中,目标树经营、无干扰、粗放经营3种模式之间生物量差异均显著(P<0.05);阔叶林乔木中,目标树经营模式的生物量与无干扰和粗放经营模式间差异显著,而后两者之间差异不显著(P>0.05)。
2.2 不同经营模式下3种森林类型林下植被层及地表凋落物的生物量
由表4可知,在针叶林、阔叶林和针阔混交林中,凋落物生物量排序均为目标树经营大于无干扰和粗放经营;在针叶林和阔叶林中,灌木层和草本层总生物量大小表现出相同的规律,即无干扰>粗放经营>目标树经营,而针阔混交林中灌木层和草本层总生物量表现为粗放经营>无干扰>目标树经营。针叶林中,灌木层和草本层总生物量在粗放经营与目标树经营间无明显差异,但二者与无干扰差异显著(P<0.05);凋落物生物量在目标树经营与粗放经营、无干扰经营间差异显著,但后两者差异不显著(P<0.05)。阔叶林中,灌木层总生物量在目标树经营与粗放经营、无干扰间差异显著,但后两者差异不显著(P<0.05);3种经营模式间草本层总生物量差异均显著(P<0.05);凋落物生物量在粗放经营与目标树经营、无干扰间差异显著(P<0.05),但后两者之间差异不显著。针阔混交林中,灌木层总生物量在无干扰与粗放经营、目标树经营间差异均不显著,而后两者之间差异显著(P<0.05);3种经营模式林分间草本层总生物量差异均显著(P<0.05);凋落物生物量在粗放经营与目标树经营、无干扰经营间差异不显著,而后两者之间差异显著(P<0.05)。
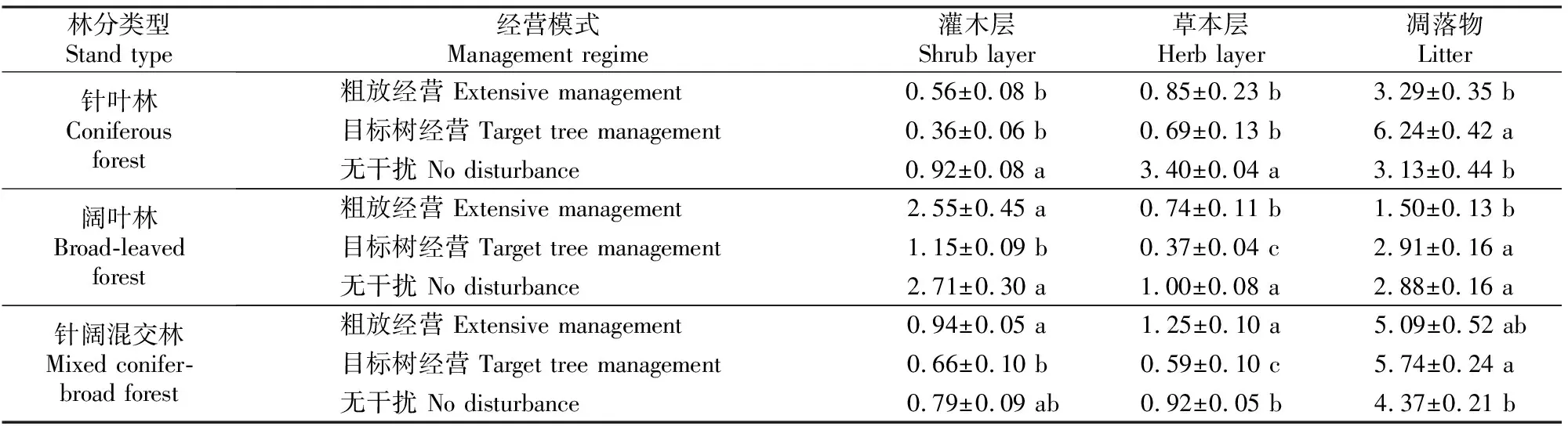
表 4 东北地区不同经营模式下3种森林类型的林下植被生物量和地表凋落物现存量
2.3 不同经营模式下3种森林类型林分总生物量及其分配格局
林分总生物量由乔木层、灌木层和草本层生物量及地表现存凋落物生物量共同构成。由表5可知,针叶林和阔叶林中,林分总生物量排序均为目标树经营>无干扰>粗放经营,针阔混交林中则为目标树经营>粗放经营>无干扰,且目标树经营与无干扰、粗放经营间差异显著(P<0.05)。

表 5 东北地区不同经营模式下3种森林类型林分总生物量及其在各林层的分配比例
目标树经营明显增加了林分总生物量,而粗放经营由于砍伐大量林木造成林分生物量降低。针叶林中,3种经营模式之间总生物量差异显著(P<0.05),乔木层、灌木层、草本层、凋落物生物量占林分总生物量的比例分别为92.55%~98.84%,0.21%~0.77%,0.48%~2.90%和2.55%~3.78%。阔叶林中,目标树经营与粗放经营、无干扰间总生物量差异显著,后两者之间差异不显著(P<0.05),乔木层、灌木层、草本层、凋落物生物量占林分总生物量的比例分别为95.78%~98.82%,0.54%~2.43%,0.23%~0.89%和1.48%~2.54%。针阔混交林中,3种经营模式之间总生物量差异显著(P<0.05),乔木层、灌木层、草本层、凋落物生物量占林分总生物量的比例分别为97.63%~98.50%,0.40%~0.73%,0.55%~0.86%和3.36%~3.91%。
3 结论与讨论
本研究利用东北不同树种枝、干、叶、根的生物量生长模型,计算获得针叶林、阔叶林、针阔混交林在粗放经营、目标树经营、无干扰下林分乔木的总生物量。在针叶林、阔叶林、针阔混交林中,乔木总生物量大小排序均为目标树经营>无干扰>粗放经营。研究区的天然次生林多处于中龄林,影响目标树生长的干扰树较多,经过目标树经营采伐掉影响目标树生长的干扰树,为目标树生长创造了更大的生长空间,减弱了周围树种对其光照、养分的竞争,从而促进其快速生长,而目标树是乔木层生长的主体,因此使得乔木层总生物量获得极大提高[16]。Baskent等[17]通过分析目标树经营和传统经营下山毛榉(Fagussylvatica)林的多年连续生物量数据,也得出目标树经营比传统经营可以获得更多林分生物量的结论。本研究中,乔木层各器官生物量所占的比例依次为干(46.52%~68.32%)>根(12.23%~38.07%)>枝(6.51%~17.93%)>叶(2.28%~4.45%), 这与李文华等[5]对长白山地区主要生态系统乔木层生物量在各器官的分配比例所得结论相似。
本研究发现,在针叶林、阔叶林、针阔混交林中,凋落物生物量排序为目标树经营大于其余2种经营模式。目标树经营可改善林分结构,补植的阔叶树种都会增加凋落物量,使得林分生产力增加,促进光合产物向下分配,凋落物量也随之增加,合理的林分结构同时促进了林下植被快速生长和凋落物的分解[18]。本研究中,针叶林、阔叶林中灌木层和草本层的总生物量均表现出相同的规律,即无干扰>粗放经营>目标树经营,针阔混交林表现为粗放经营>无干扰>目标树经营。这是因为目标树经营措施促进了乔木层林木的快速生长,对下层空间形成郁闭遮阴,因而减少了灌木、草本生长所需的光照等生长条件,使得灌木层、草本层生物量降低,同时目标树经营林分采取了清除林下灌草的措施,势必会造成灌草层生物量的减少。本研究也进一步证明,灌木层、草本层是林分生物量的组成与结构成分,研究其生物量对于整个森林生态系统物质和能量的固定、消耗、积累及转化有着重要意义[19]。本研究采用直接收获法获得灌木层、草本层的生物量,但是收获法不仅耗时费力、对生态系统破坏性较大,且存在很大的不确定性,因为灌木的分布通常不均匀,取样数量又很难满足统计学大样本的要求[20]。东北地区一些灌木如暴马丁香(Syringareticulata)、青楷槭(Acertegmentosum)、花楷槭(Acerukurunduense)的空间异质性较大,直接采用收获法难以对其生物量进行准确估计[14,21],下一步研究可通过建立模型的方法进行灌木、草本生物量的估计。
本研究发现,在针叶林和阔叶林中,林分总生物量排序均为目标树经营>无干扰>粗放经营,而针阔混交林为目标树经营>粗放经营>无干扰,且目标树经营与无干扰、粗放经营间差异显著。9种林分中,不同林层生物量大小排序为乔木层>凋落物>灌木层和草本层。目标树经营对于提高天然次生林林分质量具有明显效果[22-23]。采伐干扰树可为目标树及其林分整体提供适宜的生长空间,从而促进林分生物量的增加。Obergfoell[24]对比德国下萨克森州目标树经营和传统经营林分,发现目标树经营的林分具有更高的生物量。我国大部分天然次生林存在结构不完整、中幼龄比重大、森林质量低下等问题,难以充分发挥森林的各种生态效益功能。处于中高纬度的东北林区是我国主要林区之一,其林地面积约占全国林地总面积的31.4%,在我国碳汇管理和生态环境建设中起着举足轻重的作用[25]。本研究结果表明,目标树经营模式可以改善林分结构,提高林分生物量,平衡生态功能,满足人类对森林多功能的需求。
[参考文献]
[1] Houghton R A,Skole D L,Nobre C A.Annual fluxes of carbon from deforestation and regrowth in the Brazilian Amazon [J].Nature,2000,20:301-304.
[2] Dixon R K,Brown S,Houghton R A,et al.Carbon pools and flux of global forest ecosystems [J].Science,1994,263:185-190.
[3] Fallkowski P,Scholes R J,Boyle E,et al.The global carbon cycle:A test of our knowledge of earth as system [J].Science,2000,290:291-296.
[4] Houghton R A.Balancing the global carbon budget [J].Annual Review of Earth and Planetary Sciences,2007,35:313-347.
[5] 李文华,灯坤纹,李 飞.长白山主要生态系统生物量的研究 [J].森林生态系统研究,1980:43-50.
Li W H,Deng K W,Li F.Study on main ecosystem biomass in Changbai mountains [J].Forest Ecosystem Research,1980:43-50.(in Chinese)
[6] 刘世荣,柴一新,蔡体久,等.兴安落叶松人工群落生物量与净初级生产力的研究 [J].东北林业大学学报,1990,18(2):40-46.
Liu S R,Chai Y X,Cai T J,et al.Study on biomass and net primary productivity ofLarixlarchplantation community [J].Journal of Northeast Forestry University,1990,18(2):40-46.(in Chinese)
[7] 国庆喜,张 锋.基于遥感信息估测森林的生物量 [J].东北林业大学学报,2003,31(2):13-16.
Guo Q X,Zhang F.Estimation of forest biomass on remote sensing [J].Journal of Northeast Forestry University,2003,31(2):13-16. (in Chinese)
[8] 李晓娜,国庆喜,王兴昌,等.东北天然次生林下总生物量的相对生长 [J].东北林业大学学报,2010,46(8):23-32.
Li X N,Guo Q X,Wang X C,et al.Total biomass relative growth of natural secondary forest in Northeast [J].Journal of Northeast Forestry University,2010,46(8):23-32.(in Chinese)
[9] 邬可义.哈尔滨市的森林近自然和价值化经营实践 [J].世界林业研究,2010,22(6):14-18.
Wu K Y.Close to natural and valuable management of Harbin forest [J].World Forestry Research,2010,22(6):14-18.(in Chinese)
[10] 梁星云,何友均,张 谱,等.不同经营模式对丹清河林场天然次生林植物群落结构及其多样性的影响 [J].林业科学,2013,49(3):93-102.
Liang X Y,He Y J,Zhang P,et al.Effects of different forest management regimes on plant community structure and biodiversity of natural secondary forests in Danqinghe forestry farm [J].Forestry Science,2013,49(3):93-102.(in Chinese)
[11] 陈传国,郭杏芬.阔叶红松林生物量的回归方程 [J].延边林业科技,1983,2(1):2-19.
Chen C G,Guo X F.The biomass regression equation of Koream pine forest [J].Yanbian Forestry Technique,1983,2(1):2-19.(in Chinese)
[12] 丁宝永. 落叶松人工林群落生物生产力的研究 [J].植物生态学与植物学学报,1989,4(3):226-236.
Ding B Y.Biomass and productivity of Larch plantation community [J].Journal of Plant Ecology and Botany,1989,4(3):226-236.(in Chinese)
[13] 徐振邦,李 昕.长白山阔叶红松林生物生产量的研究 [J].森林生态系统研究,1985(5):33-46.
Xu Z B,Li X.Biomass of broad-leaved Korean pine forest in Changbai mountain [J].Forest Ecology System Research,1985(5):33-46.(in Chinese)
[14] 何列艳,亢新刚.长白山区林下主要灌木生物量估计与分析 [J].南京林业大学学报:自然科学版,2011,35(5):45-50.
He L Y,Kang X G.Estimation and analysis of main shrub in Changbai Mountain [J].Journal of Nanjing Forestry University:Natural Science,2011,35(5):45-50.(in Chinese)
[15] 樊后保,李燕燕,苏兵强,等.马尾松-阔叶树混交异龄林生物量与生产力分配格局 [J].生态学报,2006,26(8):2463-2473.
Fan H B,Li Y Y,Su B Q,et al.Allocation pattern of biomass and productivity in the mixed uneven-aged stands of Masson’s pine and hardwood species [J].Acta Ecologica Sinica,2006,26(8):2463-2473.(in Chinese)
[16] Larsen J B,Nielsen A B.Nature-based forest management-where are we going? Elaborating forest development types in and with practice [J].Forest Ecology and Management,2007,238:107-117.
[17] Baskent E Z,Keles S.Developing alternative forest management planning strategies incorporating timber,water and carbon values: An examination of their interactions [J].Environmental Model Assess,2008,10:323-335.
[18] Zhang D H,Ye Z F.Influence of thinning on soil fertility in artificial forests [J].Chinese Journal of Applied Ecology,2001,12:672-676.
[19] 曾慧卿,刘琪璟,冯宗炜,等.基于冠幅和植株高度的灌木生物量回归方程 [J].南京林业大学学报:自然科学版,2006,30(4):101-104.
Zeng H Q,Liu Q J,Feng Z W,et al.The shrub regression equation based on crown and height [J].Journal of Nanjing Forestry University:Natural Science,2006,30(4):101-104.(in Chinese)
[20] Alaback B P.Biomass regression equations for understory plants in coastal Alaska:Effects of species and sampling design on estimates [J].Northwest Science,1986,60(2):187-196.
[21] Patona D,Nunez J,Bao D,et al.Forage biomass of 22 shrub species from Natural Park assessed by log-log regression models [J].Journal of Arid Environments,2002,52(2):223-231.
[22] Knoke T.Analyse und optimierung der holzproduktion ineinem plenterwald,Zur forstbetriebsplanung in ungleichaltrigen waldern Foresl [M].Munchen:Forschungsber,1998:170.
[23] Sturm K.Methoden und ziele der waldbiotopkartierung [J].Naturschutz in der Forstwirtschaft Biologie und Schutz der Fledermause im Wald,1993(4):251-257.
[24] Obergfoell P.Modelle der nutzungsplanung auf der basis von wiederholungsinventuren [J].Forest Modeling,2000(6):433-441.
[25] Wang C.Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests [J].Forest Ecology and Management,2006,222:9-16.
